

Interspecific Drought Cuing in Plants

Abstract
:1. Introduction
2. Results
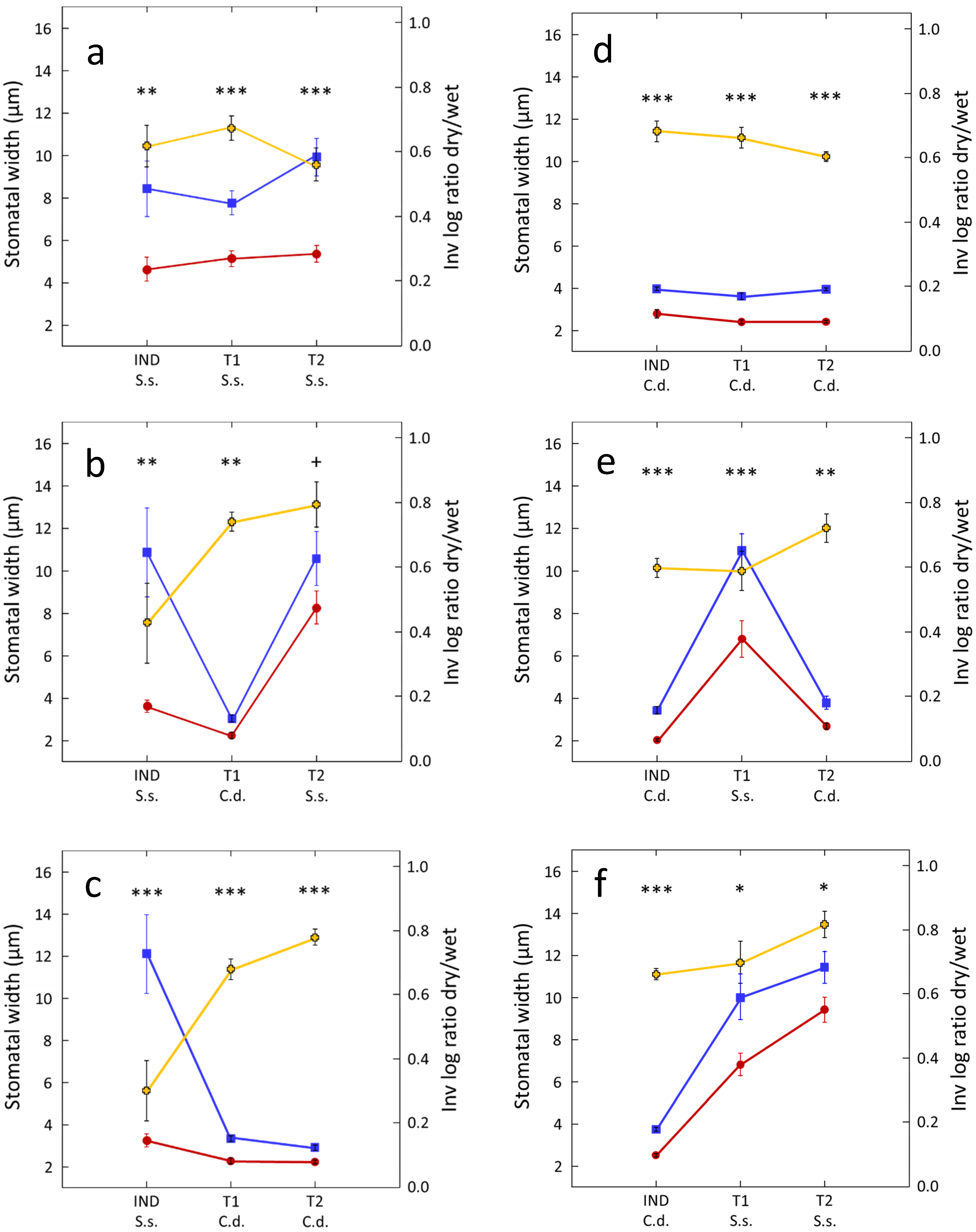
2.1. Intraspecific Drought Cuing
2.2. Interspecific Drought Cuing
3. Discussion
4. Materials and Methods
4.1. Plant Material
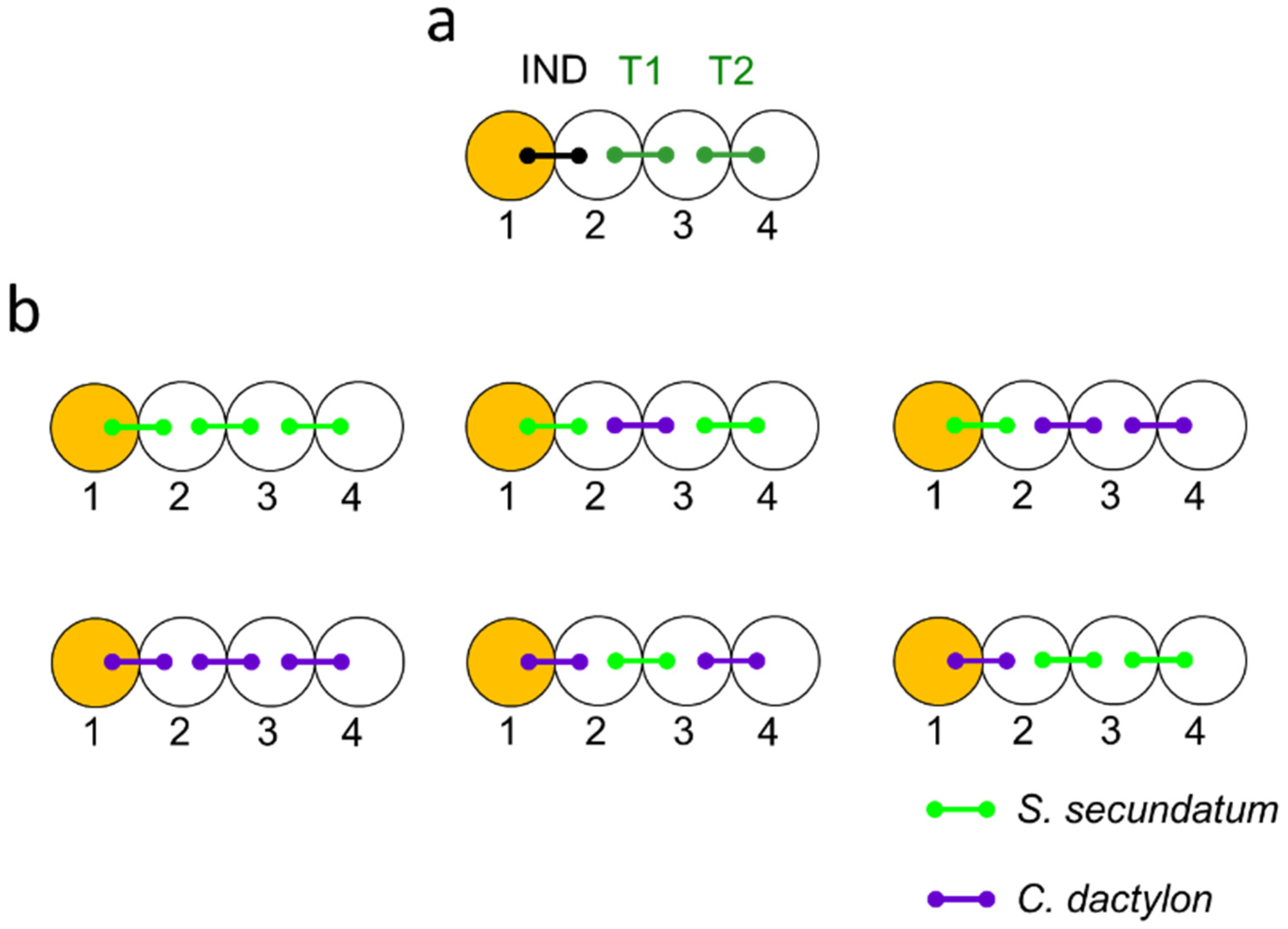
4.2. Experiment Design
- Intraspecific cuing: S.s.–S.s.–S.s., C.d.–C.d.–C.d.;
- Interspecific cuing (S. secundatum stressed-induced): S.s.–C.s.–S.s., S.s–C.d.–C.d.;
- Interspecific cuing (C. dactylon stressed-induced): C.d–S.s.–C.d., C.d.–S.s.–S.s.
4.3. Growth Conditions and Experimental Setup
4.4. Stomata Measurements
4.5. Data Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Messer, P.W.; Ellner, S.P.; Hairston, N.G. Can Population Genetics Adapt to Rapid Evolution. Trends Genet. 2016, 32, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, A.D. Evolutionary Significance of Phenotypic Plasticity in Plants. Adv. Gen. 1965, 13, 115–155. [Google Scholar] [CrossRef]
- Levins, R. Evolution in Changing Environments; Princeton University Press: Princeton, NJ, USA, 1968. [Google Scholar]
- Schlichting, C.D. The Evolution of Phenotypic Plasticity in Plants. Annu. Rev. Ecol. Syst. 1986, 17, 667–693. [Google Scholar] [CrossRef]
- Gavrilets, S.; Scheiner, S.M. The Genetics of Phenotypic Plasticity. VI. Theoretical Predictions for Directional Selection. J. Evol. Biol. 1993, 6, 49–68. [Google Scholar] [CrossRef]
- Alpert, P.; Simms, E. The Relative Advantages of Plasticity and Fixity in Different Environments: When Is It Good for a Plant to Adjust. Evol. Ecol. 2002, 16, 285–297. [Google Scholar] [CrossRef]
- Lande, R. Adaptation to an Extraordinary Environment by Evolution of Phenotypic Plasticity and Genetic Assimilation. J. Evol. Biol. 2009, 22, 1435–1446. [Google Scholar] [CrossRef]
- Novoplansky, A. Picking Battles Wisely: Plant Behaviour under Competition. Plant. Cell Environ. 2009, 32, 726–741. [Google Scholar] [CrossRef]
- Botero, C.A.; Weissing, F.J.; Wright, J.; Rubenstein, D.R. Evolutionary Tipping Points in the Capacity to Adapt to Environmental Change. Proc. Natl. Acad. Sci. USA 2015, 112, 184–189. [Google Scholar] [CrossRef] [Green Version]
- Tufto, J. Genetic Evolution, Plasticity, and Bet-Hedging as Adaptive Responses to Temporally Autocorrelated Fluctuating Selection: A Quantitative Genetic Model. Evolution 2015, 69, 2034–2049. [Google Scholar] [CrossRef] [Green Version]
- Pigliucci, M. Phenotypic Plasticity: Beyond Nature and Nurture; Johns Hopkins University Press: Baltimore, MD, USA, 2001. [Google Scholar]
- Sultan, S.E. Plasticity as an Intrinsic Property of Organisms. In Phenotypic Plasticity and Evolution: Causes, Consequences, Controversies; Pfennig, D., Ed.; CRC Press: Boca Raton, FL, USA, 2021; pp. 3–24. [Google Scholar] [CrossRef]
- Aphalo, P.J.; Ballare, C.L. On the Importance of Information-Acquiring Systems in Plant-Plant Interactions. Funct. Ecol. 1995, 9, 5–14. [Google Scholar] [CrossRef]
- Shemesh, H.; Rosen, R.; Eshel, G.; Novoplansky, A.; Ovadia, O. The Effect of Steepness of Temporal Resource Gradients on Spatial Root Allocation. Plant Signal. Behav. 2011, 6, 1356–1360. [Google Scholar] [CrossRef] [Green Version]
- Novoplansky, A. Future Perception in Plants. In Anticipation Across Disciplines; Nadin, M., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; pp. 57–70. [Google Scholar] [CrossRef]
- Falik, O.; Mauda, S.; Novoplansky, A. The Ecological Implications of Interplant Drought Cuing. J. Ecol. 2022, 111, 23–32. [Google Scholar] [CrossRef]
- Miller, M.B.; Bassler, B.L. Quorum Sensing in Bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [Green Version]
- Bruin, J.; Dicke, M.; Sabelis, M. Plants Are Better Protected against Spider-Mites after Exposure to Volatiles from Infested Conspecifics. Experientia 1992, 48, 525–529. [Google Scholar] [CrossRef]
- Arimura, G.; Matsui, K.; Takabayashi, J. Chemical and Molecular Ecology of Herbivore-Induced Plant Volatiles: Proximate Factors and Their Ultimate Functions. Plant Cell Physiol. 2009, 50, 911–923. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.Y.; Zeng, R.S.; Xu, J.F.; Li, J.; Shen, X.; Yihdego, W.G. Interplant Communication of Tomato Plants through Underground Common Mycorrhizal Networks. PLoS ONE 2010, 5, e13324. [Google Scholar] [CrossRef] [Green Version]
- Caparrotta, S.; Boni, S.; Taiti, C.; Palm, E.; Mancuso, S.; Pandolfi, C. Induction of Priming by Salt Stress in Neighboring Plants. Environ. Exp. Bot. 2017, 147, 261–270. [Google Scholar] [CrossRef]
- Shulaev, V.; Silverman, P.; Raskin, I. Airborne Signalling by Methyl Salicylate in Plant Pathogen Resistance. Nature 1997, 385, 718–721. [Google Scholar] [CrossRef]
- Yi, H.S.; Heil, M.; Adame-Álvarez, R.M.; Ballhorn, D.J.; Ryu, C.M. Airborne Induction and Priming of Plant Defenses against a Bacterial Pathogen. Plant Physiol. 2009, 151, 2152–2161. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, L.; Johnson, D. Plant–Plant Communication Through Common Mycorrhizal Networks. Adv. Bot. Res. 2017, 82, 83–97. [Google Scholar] [CrossRef]
- Oda, R.; Masataka, N. Interspecific Responses of Ringtailed Lemurs to Playback of Antipredator Alarm Calls Given by Verreaux’s Sifakas. Ethology 1996, 102, 441–453. [Google Scholar] [CrossRef]
- Zuberbühler, K. Interspecies Semantic Communication in Two Forest Primates. Proc. Biol. Sci. 2000, 267, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Dawson Pell, F.S.E.; Potvin, D.A.; Ratnayake, C.P.; Fernández-Juricic, E.; Magrath, R.D.; Radford, A.N. Birds Orient Their Heads Appropriately in Response to Functionally Referential Alarm Calls of Heterospecifics. Anim. Behav. 2018, 140, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Carlson, N.; Greene, E.; Templeton, C. Nuthatches Vary Their Alarm Calls Based upon the Source of the Eavesdropped Signals. Nat. Commun. 2020, 11, 526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ton, J.; D’Alessandro, M.; Jourdie, V.; Jakab, G.; Karlen, D.; Held, M.; Mauch-Mani, B.; Turlings, T.C.J. Priming by Airborne Signals Boosts Direct and Indirect Resistance in Maize. Plant J. 2007, 49, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Karban, R. Plant Communication. Annu. Rev. Ecol. Evol. Syst. 2021, 52, 1–24. [Google Scholar] [CrossRef]
- Karban, R.; Baldwin, I.; Baxter, K.J.; Laue, G.; Felton, G. Communication between Plants: Induced Resistance in Wild Tobacco Plants Following Clipping of Neighboring Sagebrush. Oecologia 2000, 125, 66–71. [Google Scholar] [CrossRef]
- Yao, Y.; Danna, C.H.; Zemp, F.J.; Titov, V.; Ciftci, O.N.; Przybylski, R.; Ausubel, F.M.; Kovalchuk, I. UV-C–Irradiated Arabidopsis and Tobacco Emit Volatiles That Trigger Genomic Instability in Neighboring Plants. Plant Cell 2011, 23, 3842–3852. [Google Scholar] [CrossRef] [Green Version]
- Pearse, I.S.; Porensky, L.M.; Yang, L.H.; Stanton, M.L.; Karban, R.; Bhattacharyya, L.; Cox, R.; Dove, K.; Higgins, A.; Kamoroff, C.; et al. Complex Consequences of Herbivory and Interplant Cues in Three Annual Plants. PLoS ONE 2012, 7, e38105. [Google Scholar] [CrossRef]
- Gorzelak, M.A.; Asay, A.K.; Pickles, B.J.; Simard, S.W. Inter-Plant Communication through Mycorrhizal Networks Mediates Complex Adaptive Behaviour in Plant Communities. AoB Plants 2015, 7, plv050. [Google Scholar] [CrossRef] [Green Version]
- Falik, O.; Mordoch, Y.; Quansah, L.; Fait, A.; Novoplansky, A. Rumor Has It: Relay Communication of Stress Cues in Plants. PLoS ONE 2011, 6, e23625. [Google Scholar] [CrossRef] [Green Version]
- Falik, O.; Novoplansky, A. Is ABA the exogenous vector of interplant drought cuing. Plant Signal. Behav. 2022, 17, 2129295. [Google Scholar] [CrossRef]
- Falik, O.; Mordoch, Y.; Ben-Natan, D.; Vanunu, M.; Goldstein, O.; Novoplansky, A. Plant Responsiveness to Root-Root Communication of Stress Cues. Ann. Bot. 2012, 110, 271–280. [Google Scholar] [CrossRef]
- Balacey, S.L. Investigating the Role of Volatile Signalling In Plant Responses To Drought. Ph.D Thesis, University of Adelaide, Adelaide, Australia, 2021. [Google Scholar]
- Jiang, Y.; Ye, J.; Niinemets, Ü. Dose-dependent methyl jasmonate effects on photosynthetic traits and volatile emissions: Biphasic kinetics and stomatal regulation. Plant Signal. Behav. 2021, 16, 1917169. [Google Scholar] [CrossRef]
- Wenig, M.; Ghirardo, A.; Sales, J.H.; Pabst, E.S.; Breitenbach, H.H.; Antritter, F.; Weber, B.; Lange, B.; Lenk, M.; Cameron, R.K.; et al. Systemic Acquired Resistance Networks Amplify Airborne Defense Cues. Nat. Commun. 2019, 10, 3813. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Baldwin, I.T. New Insights into Plant Responses to the Attack from Insect Herbivores. Annu. Rev. Genet. 2010, 44, 1–24. [Google Scholar] [CrossRef]
- Johnson, R.; Puthur, J.T. Seed Priming as a Cost Effective Technique for Developing Plants with Cross Tolerance to Salinity Stress. Plant Physiol. Biochem. 2021, 162, 247–257. [Google Scholar] [CrossRef]
- Bernsdorff, F.; Döring, A.C.; Gruner, K.; Schuck, S.; Bräutigam, A.; Zeier, J. Pipecolic Acid Orchestrates Plant Systemic Acquired Resistance and Defense Priming via Salicylic Acid-Dependent and -Independent Pathways. Plant Cell 2016, 28, 102–129. [Google Scholar] [CrossRef] [Green Version]
- Farmer, E.E. Jasmonate Perception Machines. Nature 2007, 448, 659–660. [Google Scholar] [CrossRef]
- Dicke, M.; Bruin, J. Chemical Information Transfer between Plants: Back to the Future. Biochem. Syst. Ecol. 2001, 29, 981–994. [Google Scholar] [CrossRef]
- Cheplick, G.P. Sibling Competition Is a Consequence of Restricted Dispersal in an Annual Cleistogamous Grass. Ecology 1993, 74, 2161–2164. [Google Scholar] [CrossRef]
- Herben, T.; Novoplansky, A. Implications of Self/Non-Self Discrimination for Spatial Patterning of Clonal Plants. Evol. Ecol. 2008, 22, 337–350. [Google Scholar] [CrossRef]
- Whigham, D.; Chapa, A. Timing and Intensity of Herbivory: Its Influence on the Performance of Clonal Woodland Herbs. Plant Species Biol. 1999, 14, 29–37. [Google Scholar] [CrossRef]
- Karban, R.; Shiojiri, K.; Huntzinger, M.; McCall, A.C. Damage-induced Resistance in Sagebrush: Volatiles are key to understand Intra- and Interplant Communication. Ecology 2006, 87, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Saona, C.R.; Rodriguez-Saona, L.E.; Frost, C.J. Herbivore-Induced Volatiles in the Perennial Shrub, Vaccinium Corymbosum, and Their Role in Inter-Branch Signaling. J. Chem. Ecol. 2009, 35, 163–175. [Google Scholar] [CrossRef]
- Bertness, M.D.; Callaway, R. Positive Interactions in Communities. Trends Ecol. Evol. 1994, 9, 191–193. [Google Scholar] [CrossRef]
- Butterfield, B.J. Effects of Facilitation on Community Stability and Dynamics: Synthesis and Future Directions. J. Ecol. 2009, 97, 1192–1201. [Google Scholar] [CrossRef]
- Ploughe, L.W.; Jacobs, E.M.; Frank, G.S.; Greenler, S.M.; Smith, M.D.; Dukes, J.S. Community Response to Extreme Drought (CRED): A Framework for Drought-Induced Shifts in Plant–Plant Interactions. New Phytol. 2019, 222, 52–69. [Google Scholar] [CrossRef] [Green Version]
- Simard, S.W.; Beiler, K.J.; Bingham, M.A.; Deslippe, J.R.; Philip, L.J.; Teste, F.P. Mycorrhizal Networks: Mechanisms, Ecology and Modelling. Fungal Biol. Rev. 2012, 26, 39–60. [Google Scholar] [CrossRef]
- Remke, M.J.; Johnson, N.C.; Wright, J.; Williamson, M.; Bowker, M.A. Sympatric Pairings of Dryland Grass Populations, Mycorrhizal Fungi and Associated Soil Biota Enhance Mutualism and Ameliorate Drought Stress. J. Ecol. 2021, 109, 1210–1223. [Google Scholar] [CrossRef]
- Kotlarz, J.; Nasiłowska, S.; Rotchimmel, K.; Kubiak, K.; Kacprzak, M. Species Diversity of Oak Stands and Its Significance for Drought Resistance. Forests 2018, 9, 126. [Google Scholar] [CrossRef] [Green Version]
- Aguirre, B.A.; Hsieh, B.; Watson, S.J.; Wright, A.J. The Experimental Manipulation of Atmospheric Drought: Teasing out the Role of Microclimate in Biodiversity Experiments. J. Ecol. 2021, 109, 1986–1999. [Google Scholar] [CrossRef]
- Liu, D.; Wang, T.; Peñuelas, J.; Piao, S. Drought Resistance Enhanced by Tree Species Diversity in Global Forests. Nat. Geosci. 2022, 15, 800–804. [Google Scholar] [CrossRef]
- Mahall, B.E.; Callaway, R.M. Root Communication among Desert Shrubs. Proc. Natl. Acad. Sci. USA 1991, 88, 874–876. [Google Scholar] [CrossRef] [Green Version]
- Callaway, R. The Detection of Neighbors by Plants. Trends Ecol. Evol. 2002, 17, 104–105. [Google Scholar] [CrossRef]
- Gruntman, M.; Novoplansky, A. Physiologically Mediated Self/Non-Self Discrimination in Roots. Proc. Natl. Acad. Sci. USA 2004, 101, 3863–3867. [Google Scholar] [CrossRef] [Green Version]
- Mescher, M.; Runyon, J.; De Moraes, C. Plant Host Finding by Parasitic Plants. Plant Signal. Behav. 2006, 1, 284–286. [Google Scholar] [CrossRef] [Green Version]
- Semchenko, M.; Saar, S.; Lepik, A. Plant Root Exudates Mediate Neighbour Recognition and Trigger Complex Behavioural Changes. New Phytol. 2014, 204, 631–637. [Google Scholar] [CrossRef]
- Ninkovic, V.; Markovic, D.; Dahlin, I. Decoding Neighbour Volatiles in Preparation for Future Competition and Implications for Tritrophic Interactions. Perspect. Plant Ecol. Evol. Syst. 2016, 23, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Falik, O.; Reides, P.; Gersani, M.; Novoplansky, A. Self/Non-Self Discrimination in Roots. J. Ecol. 2003, 91, 525–531. [Google Scholar] [CrossRef] [Green Version]
- Kigathi, R.N.; Weisser, W.W.; Reichelt, M.; Gershenzon, J.; Unsicker, S.B. Plant Volatile Emission Depends on the Species Composition of the Neighboring Plant Community. BMC Plant Biol. 2019, 19, 58. [Google Scholar] [CrossRef] [PubMed]
- Mahall, B.E.; Callaway, R.M. Effects of Regional Origin and Genotype on Intraspecific Root Communication in the Desert Shrub Ambrosia Dumosa (Asteraceae). Am. J. Bot. 1996, 83, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, O.N. Establishment of Cynodon Dactylon from Stolon and Rhizome Fragments. Weed Res. 2003, 43, 130–138. [Google Scholar] [CrossRef]
- Holm, L.G.; Plunknett, D.L.; Pancho, J.V.; Herberger, J.P. The World’s Worst Weeds: Distribution and Biology; Krieger Publishing Company: Malabar, FL, USA, 1991. [Google Scholar]
- Gould, F.W. Grasses of the Southwestern United States; The University of Arizona Press: Tucson, AZ, USA, 1951. [Google Scholar]
- Parker, K.F. An Illustrated Guide to Arizona Weeds; The University of Arizona Press: Tucson, AZ, USA, 1972. [Google Scholar]
- Horowitz, M. Bermudagrass (Cynodon Dactylon): A History of the Weed and Its Control in Israel. Phytoparasitica 1996, 24, 305–320. [Google Scholar] [CrossRef]
- CABI. Stenotaphrum Secundatum. Invasive Species Compendium. Available online: www.cabi.org/isc (accessed on 1 October 2022).
- Busey, P. St. Augustine Grass, Stenotaphrum Secundatum (Walt.) Kuntze. In Biology, Breeding, and Genetics of Turfgrasses; Casler, M.D., Duncan, R.R., Eds.; John and Wiley and Sons: Hoboken, NJ, USA, 2003; pp. 309–330. [Google Scholar]
- Geren, H.; Avcioglu, R.; Curaoglu, M. Performances of Some Warm-Season Turfgrasses under Mediterranean Conditions. African J. Biotechnol. 2009, 8, 4469–4474. [Google Scholar]
- Fuentealba, M.P.; Zhang, J.; Kenworthy, K.; Erickson, J.; Kruse, J.; Trenholm, L. Transpiration Responses of Warm-Season Turfgrass in Relation to Progressive Soil Drying. Sci. Hortic. 2016, 198, 249–253. [Google Scholar] [CrossRef]
- Kim, K.S.; Beard, J.B. Comparative Drought Resistances Among Eleven Warm-Season Turfgrasses and Associated Plant Parameters. Weed Turfgrass Sci. 2018, 7, 239–245. [Google Scholar] [CrossRef]
- Price, E.A.C.; Hutchings, M.J. The Causes and Developmental Effects of Integration and Independence between Different Parts of Glechoma Hederacea Clones. Oikos 1992, 63, 376–386. [Google Scholar] [CrossRef]
- Gutbrodt, B.; Mody, K.; Wittwer, R.; Dorn, S. Within-Plant Distribution of Induced Resistance in Apple Seedlings: Rapid Acropetal and Delayed Basipetal Responses. Planta 2011, 233, 1199–1207. [Google Scholar] [CrossRef]
- Miura, K.; Okamoto, H.; Okuma, E.; Shiba, H.; Kamada, H.; Hasegawa, P.M.; Murata, Y. SIZ1 Deficiency Causes Reduced Stomatal Aperture and Enhanced Drought Tolerance via Controlling Salicylic Acid-Induced Accumulation of Reactive Oxygen Species in Arabidopsis. Plant J. 2013, 73, 91–104. [Google Scholar] [CrossRef]
- Novoplansky, A. Hierarchy Establishment among Potentially Similar Buds. Plant. Cell Environ. 1996, 19, 781–786. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry, 4th ed.; Freeman: New York, NY, USA, 2012. [Google Scholar]



{kind=link}
{kind=link}
| Triplet Combination IND–T1–T2 | Difference in Relative Stomatal Width between T1 or T2 and IND | Student’s t-Test |
|---|---|---|
| T1 vs. IND | ||
| S.s.–C.d.–S.s. | +30% | p = 0.075 |
| S.s–C.d.–C.d. | +78% | p = 0.005 |
| C.d.–S.s.–C.d. | -6% | p = 0.338 |
| C.d.–S.s.–S.s. | +19% | p = 0.177 |
| T2 vs. IND | ||
| S.s.–C.d.–S.s. | +57% | p = 0.028 |
| S.s–C.d.–C.d. | +100% | p < 0.001 |
| C.d.–S.s.–C.d. | +24% | p = 0.071 |
| C.d.–S.s.–S.s. | +28% | p = 0.027 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Falik, O.; Novoplansky, A. Interspecific Drought Cuing in Plants. Plants 2023, 12, 1200. https://doi.org/10.3390/plants12051200
Falik O, Novoplansky A. Interspecific Drought Cuing in Plants. Plants. 2023; 12(5):1200. https://doi.org/10.3390/plants12051200
Chicago/Turabian StyleFalik, Omer, and Ariel Novoplansky. 2023. "Interspecific Drought Cuing in Plants" Plants 12, no. 5: 1200. https://doi.org/10.3390/plants12051200
APA StyleFalik, O., & Novoplansky, A. (2023). Interspecific Drought Cuing in Plants. Plants, 12(5), 1200. https://doi.org/10.3390/plants12051200