
Insect Herbivory on Main Stem Enhances Induced Defense of Primary Tillers in Rice (Oryza sativa L.)

 ,
,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growth
2.2. LF Insects
2.3. Experimental Design
2.4. LF Bioassays
2.5. Jasmonic Acid (JA) Analyses
2.6. TrypPI Analyses
2.7. Quantitative Real-Time PCR Analysis
2.8. Enzyme Activity Analysis
2.9. Data Analysis
3. Results
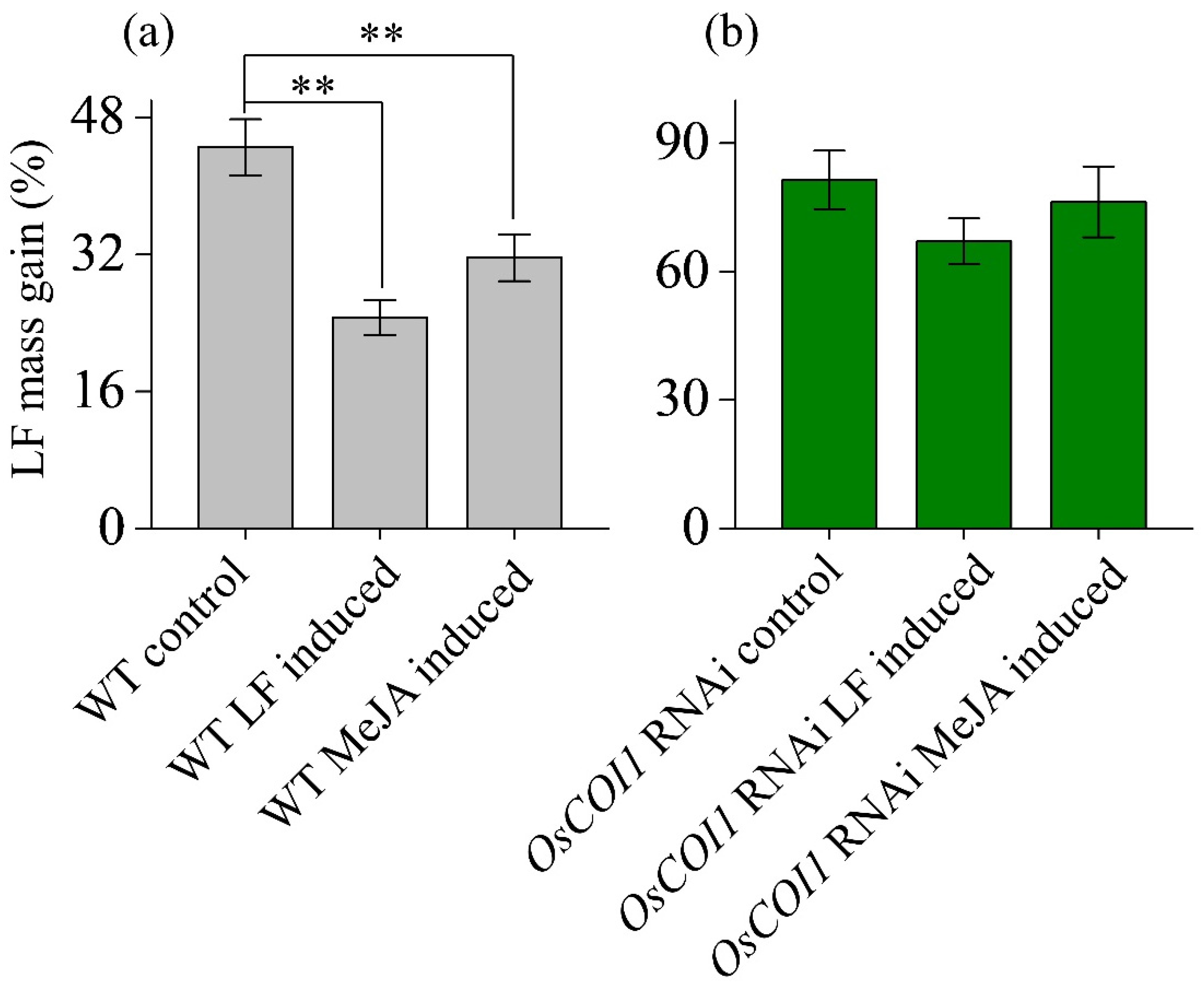
3.1. Effect on Anti-LF Resistance of Primary Tiller
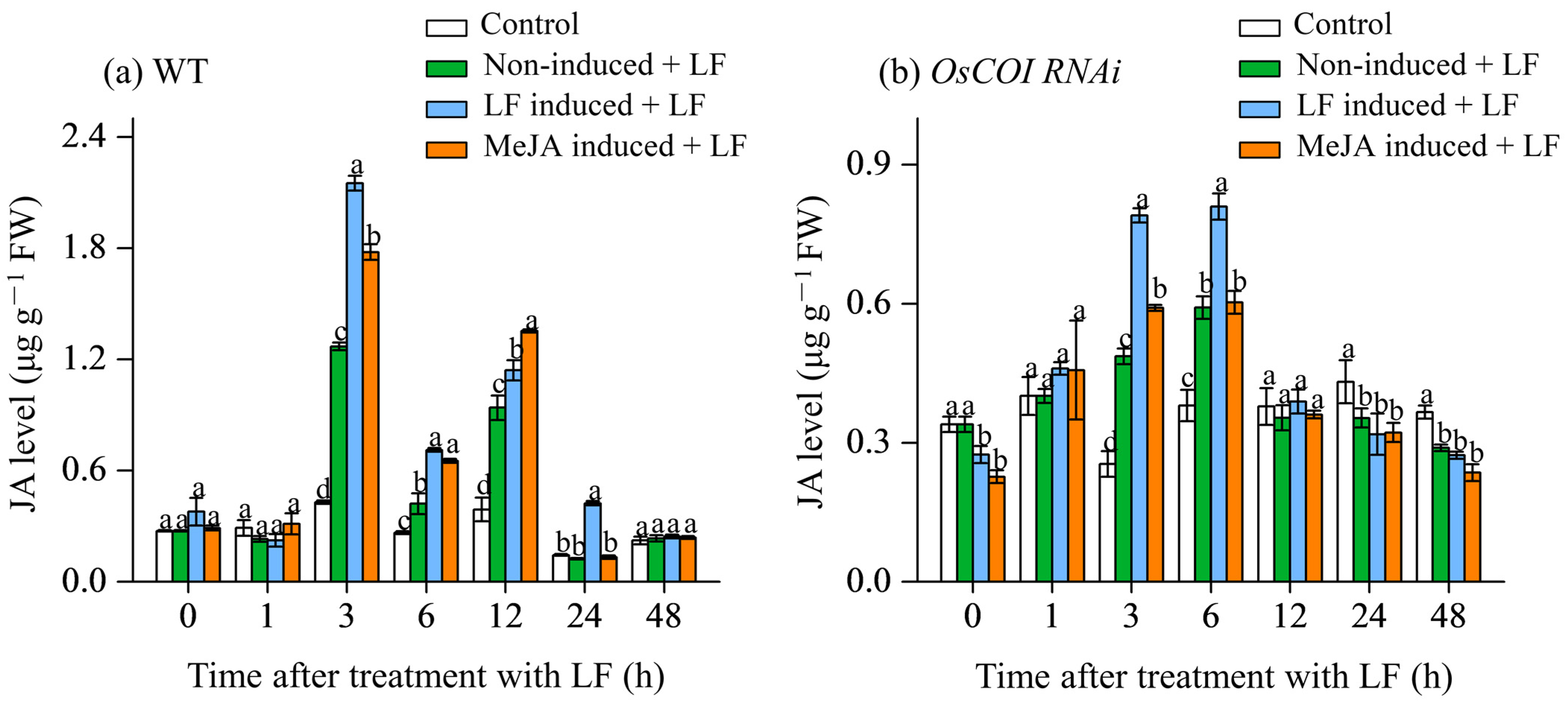
3.2. Effect on JA Levels in Primary Tiller
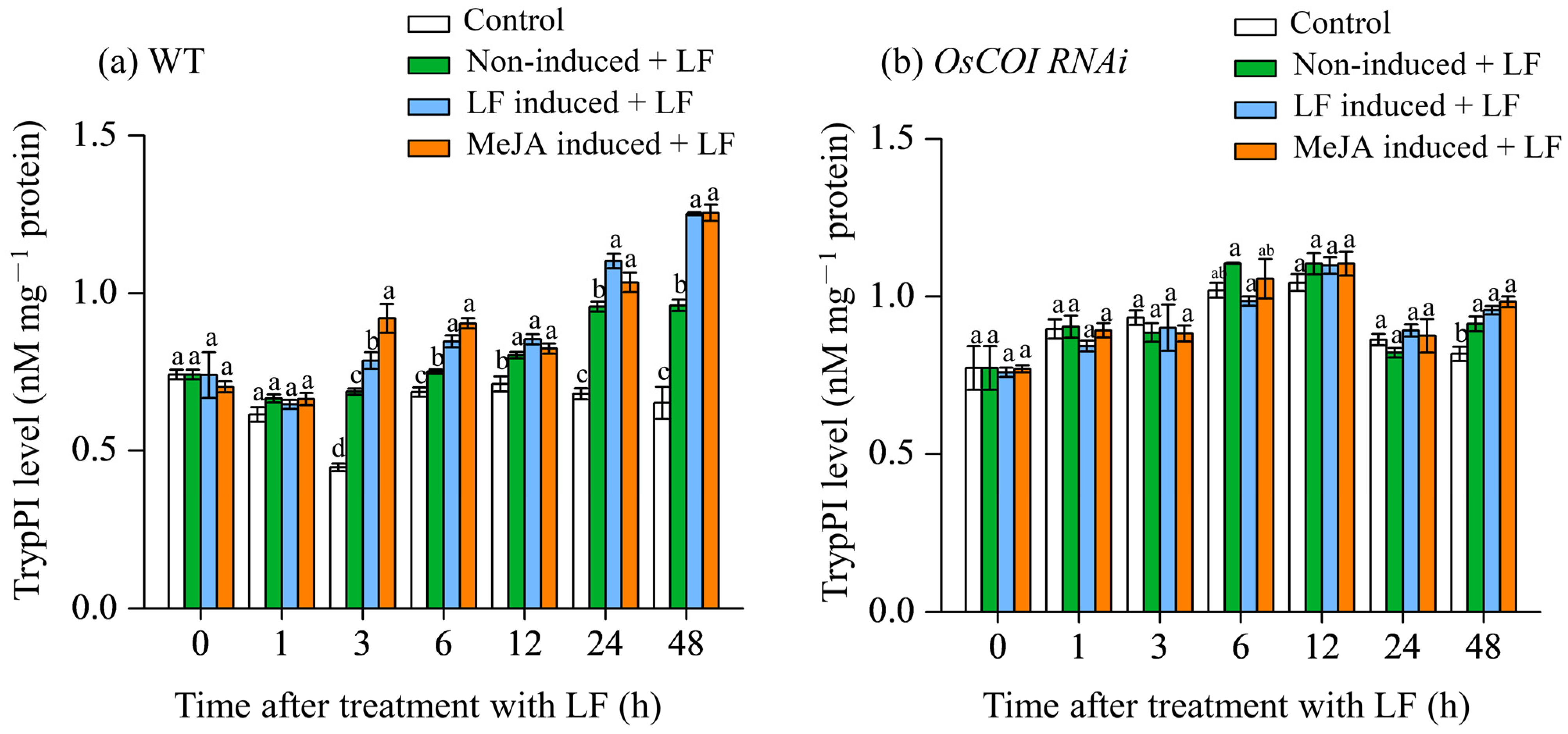
3.3. Effect on Trypsin Protease Inhibitor (TrypPI) Levels in Primary Tiller
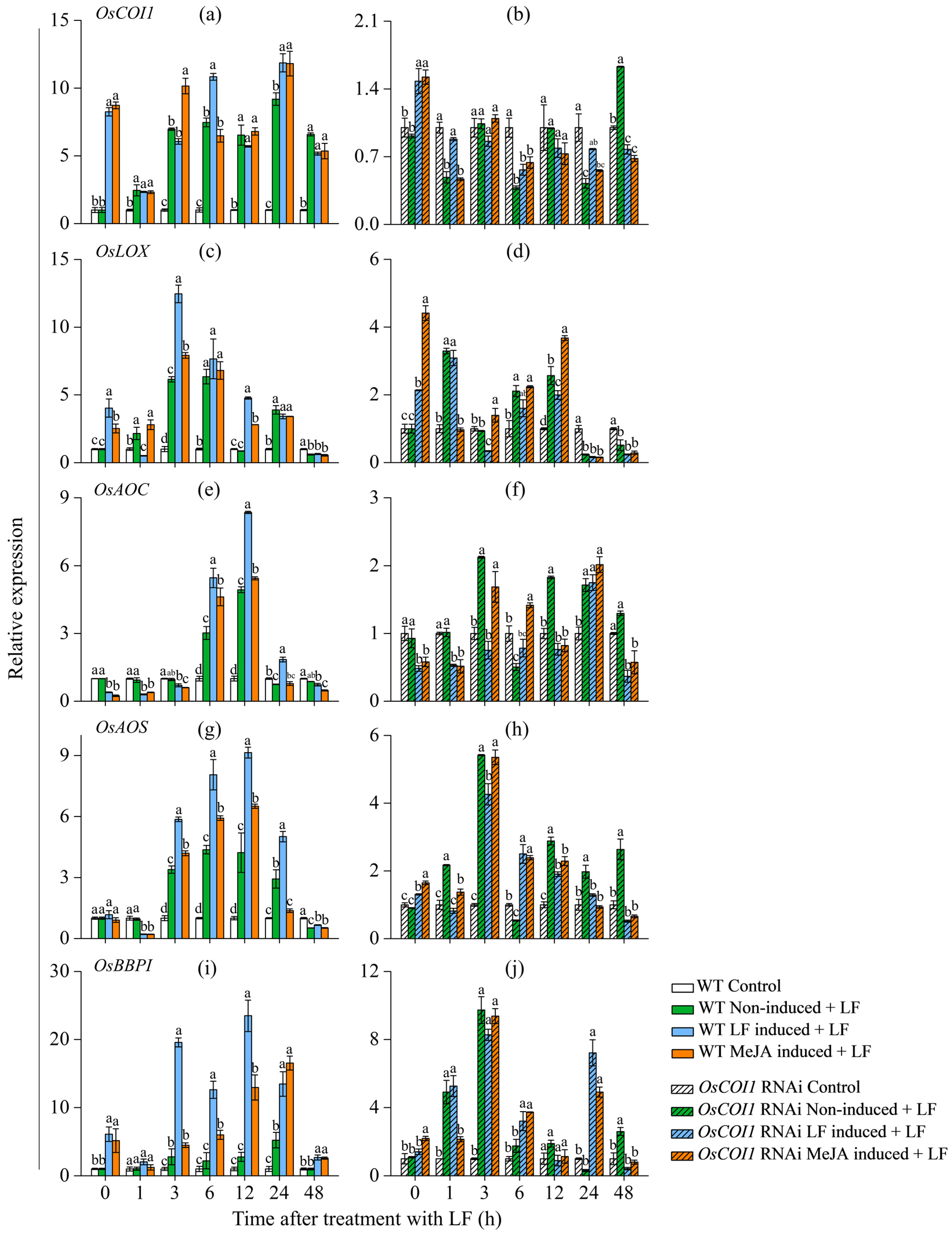
3.4. Effect on Transcript Levels of JA Signaling Genes and Bowman−Birk Protease Inhibitor Gene (OsBBPI) in Primary Tiller
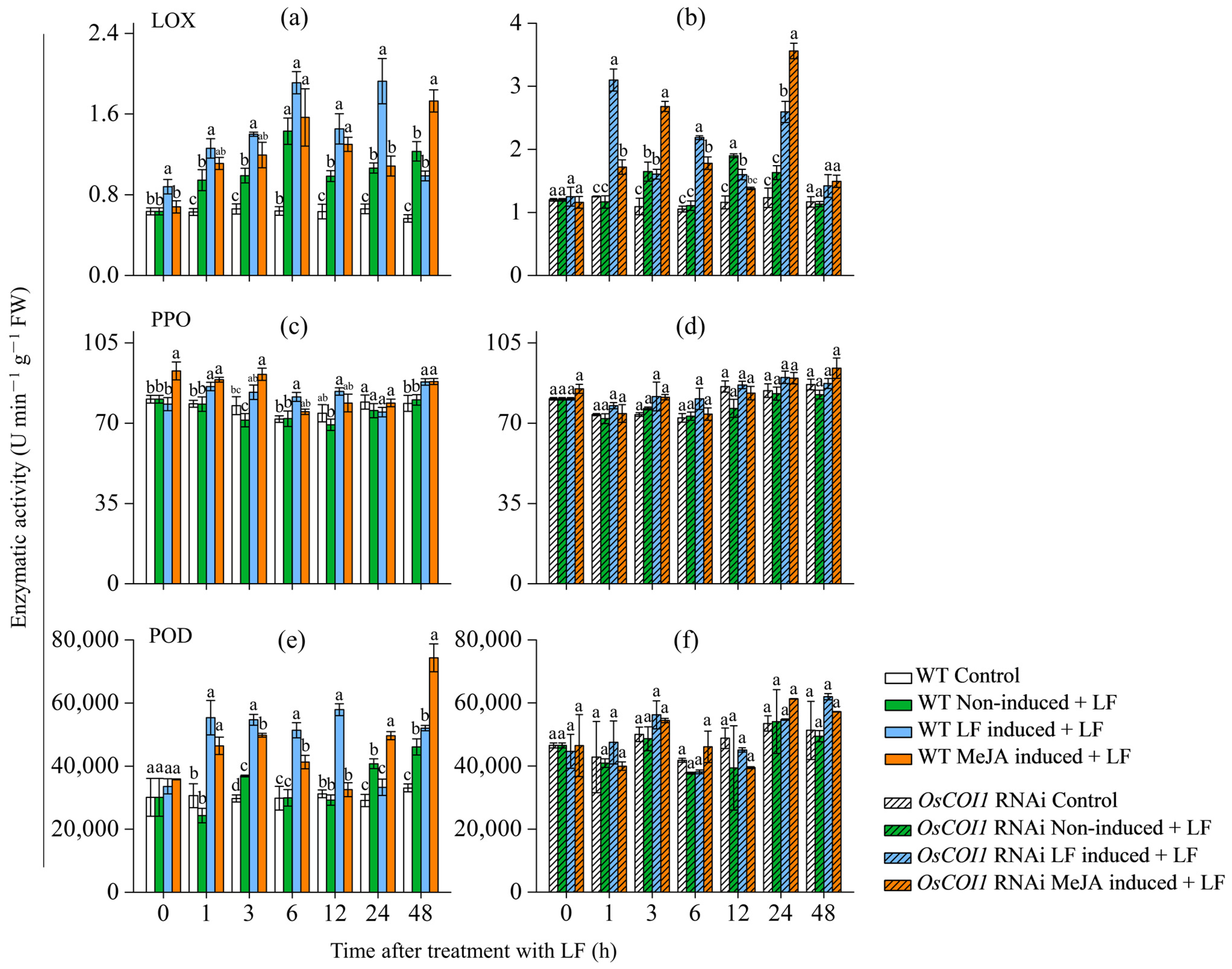
3.5. Effect on Activities of Defense-Related Enzymes in Primary Tiller
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaplan, I.; Halitschke, R.; Kessler, A.; Sardanelli, S.; Denno, R.F. Constitutive and induced defenses to herbivory in above-and belowground plant tissues. Ecology 2008, 89, 392–406. [Google Scholar] [CrossRef] [PubMed]
- Li, J.C.; Halitschke, R.; Li, D.P.; Paetz, C.; Su, H.C.; Heiling, S.; Xu, S.Q.; Baldwin, I.T. Controlled hydroxylations of diterpenoids allow for plant chemical defense without autotoxicity. Science 2021, 371, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Kessler, A.; Baldwin, I.T. Defensive function of herbivore-induced plant volatile emissions in nature. Science 2001, 291, 2141–2144. [Google Scholar] [CrossRef]
- Kant, M.R.; Jonckheere, W.; Knegt, B.; Lemos, F.; Liu, J.; Schimmel, B.C.J.; Villarroel, C.A.; Ataide, L.M.S.; Dermauw, W.; Glas, J.J. Mechanisms and ecological consequences of plant defence induction and suppression in herbivore communities. Ann. Bot. 2015, 115, 1015–1051. [Google Scholar] [CrossRef]
- Li, N.; Han, X.; Feng, D.; Yuan, D.; Huang, L.J. Signaling crosstalk between salicylic acid and ethylene/jasmonate in plant defense: Do we understand what they are whispering? Int. J. Mol. Sci. 2019, 20, 671. [Google Scholar]
- Zhang, L.; Zhang, F.; Melotto, M.; Yao, J.; He, S.Y. Jasmonate signaling and manipulation by pathogens and insects. J. Exp. Bot. 2017, 68, 1371–1385. [Google Scholar] [CrossRef] [Green Version]
- Ruan, J.J.; Zhou, Y.X.; Zhou, M.L.; Yan, J.; Khurshid, M.; Weng, W.F.; Cheng, J.P.; Zhang, K.X. Jasmonic acid signaling pathway in plants. Int. J. Mol. Sci. 2019, 20, 2479. [Google Scholar] [CrossRef] [Green Version]
- Steinbrenner, A.D.; Saldivar, E.; Hodges, N.; Palacios, N.G.; Chaparro, A.F.; Schmelz, E.A. Signatures of plant defense response specificity mediated by herbivore-associated molecular patterns in legumes. Plant J. 2022, 110, 1255–1270. [Google Scholar] [CrossRef]
- Jagodzik, P.; Tajdel-Zielinska, M.; Ciesla, A.; Marczak, M.; Ludwikow, A. Mitogen-activated protein kinase cascades in plant hormone signaling. Front. Plant Sci. 2018, 9, 1387. [Google Scholar] [CrossRef]
- Weech, M.H.; Chapleau, M.; Pan, L.; Ide, C.; Bede, J.C. Caterpillar saliva interferes with induced Arabidopsis thaliana defence responses via the systemic acquired resistance pathway. J. Exp. Bot. 2008, 59, 2437–2448. [Google Scholar] [CrossRef] [Green Version]
- Erb, M.; Reymond, P. Molecular interactions between plants and insect herbivores. Annu. Rev. Plant Biol. 2019, 70, 527–557. [Google Scholar] [CrossRef] [Green Version]
- Stratmann, J.W. Long distance run in the wound response–jasmonic acid is pulling ahead. Trends Plant Sci. 2003, 8, 247–250. [Google Scholar] [CrossRef]
- Nahar, K.; Kyndt, T.; De Vleesschauwer, D.; Höfte, M.; Gheysen, G. The jasmonate pathway is a key player in systemically induced defense against root knot nematodes in rice. Plant Physiol. 2011, 157, 305–316. [Google Scholar] [CrossRef] [Green Version]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef]
- Tian, D.; Peiffer, M.; De Moraes, C.M.; Felton, G.W. Roles of ethylene and jasmonic acid in systemic induced defense in tomato (Solanum lycopersicum) against Helicoverpa zea. Planta 2014, 239, 577–589. [Google Scholar] [CrossRef]
- Gilbert, L.; Johnson, D. Plant–plant communication through common mycorrhizal networks. Adv. Bot. Res. 2017, 82, 83–97. [Google Scholar]
- Yi, H.S.; Heil, M.; Adame-Alvarez, R.M.; Ballhorn, D.J.; Ryu, C.M. Airborne induction and priming of plant defenses against a bacterial pathogen. Plant Physiol. 2009, 151, 2152–2161. [Google Scholar] [CrossRef] [Green Version]
- Yoneya, K.; Takabayashi, J. Plant–plant communication mediated by airborne signals: Ecological and plant physiological perspectives. Plant Biotechnol. 2014, 31, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Babikova, Z.; Gilbert, L.; Bruce, T.J.; Birkett, M.; Caulfield, J.C.; Woodcock, C.; Pickett, J.A.; Johnson, D. Underground signals carried through common mycelial networks warn neighbouring plants of aphid attack. Ecol. Lett. 2013, 16, 835–843. [Google Scholar] [CrossRef]
- Song, Y.Y.; Ye, M.; Li, C.Y.; He, X.H.; Zhu-Salzman, K.Y.; Wang, R.L.; Su, Y.J.; Luo, S.M.; Zeng, R.S. Hijacking common mycorrhizal networks for herbivore-induced defence signal transfer between tomato plants. Sci. Rep. 2014, 4, 3915. [Google Scholar] [CrossRef] [Green Version]
- Hettenhausen, C.; Li, J.; Zhuang, H.; Sun, H.; Xu, Y.; Qi, J.; Zhang, J.; Lei, Y.; Qin, Y.; Sun, G.; et al. Stem parasitic plant Cuscuta australis (dodder) transfers herbivory-induced signals among plants. Proc. Natl. Acad. Sci. USA 2017, 114, E6703–E6709. [Google Scholar] [CrossRef] [Green Version]
- Bittebiere, A.K.; Benot, M.L.; Mony, C. Clonality as a key but overlooked driver of biotic interactions in plants. Perspect. Plant Ecol. Evol. Syst. 2020, 43, 125510. [Google Scholar] [CrossRef]
- Stuefer, J.F.; During, H.J.; de Kroon, H. High benefits of clonal integration in two stoloniferous species, in response to heterogeneous light environments. J. Ecol. 1994, 82, 511–518. [Google Scholar] [CrossRef]
- Roiloa, S.R.; Antelo, B.; Retuerto, R. Physiological integration modifies δ15N in the clonal plant Fragaria vesca, suggesting preferential transport of nitrogen to water-stressed offspring. Ann. Bot. 2014, 114, 399–411. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.B.; Yu, F.H.; Keser, L.H.; Dawson, W.; Fischer, M.; Dong, M.; Van Kleunen, M. United we stand, divided we fall: A meta-analysis of experiments on clonal integration and its relationship to invasiveness. Oecologia 2013, 171, 317–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keser, L.H.; Dawson, W.; Song, Y.B.; Yu, F.H.; Fischer, M.; Dong, M.; Kleunen, M. Invasive clonal plant species have a greater root-foraging plasticity than non-invasive ones. Oecologia 2014, 174, 1055–1064. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.Y.; Xu, T.T.; Wang, Y.; Li, G.Y.; Abdullah, I.; Zhong, Z.W.; Liu, J.S.; Zhu, W.Y.; Wang, L.; Wang, D.L.; et al. A meta-analysis of effects of physiological integration in clonal plants under homogeneous vs. heterogeneous environments. Funct. Ecol. 2021, 35, 578–589. [Google Scholar] [CrossRef]
- Wang, Y.J.; Liu, Y.Y.; Chen, D.; Du, D.L.; Müller-Schärer, H.; Yu, F.H. Clonal functional traits favor the invasive success of alien plants into native communities. Ecol. Appl. 2022, e2756. [Google Scholar]
- Cornelissen, J.H.C.; Song, Y.B.; Yu, F.H.; Tong, M. Plant traits and ecosystem effects of clonality: A new research agenda. Ann. Bot. 2014, 114, 369–376. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Liu, J.; Dong, M. Ecological consequences of clonal integration in plants. Front. Plant Sci. 2016, 7, 770. [Google Scholar] [CrossRef] [Green Version]
- Stuefer, J.F.; Gómez, S.; Mölken, T. Clonal integration beyond resource sharing: Implications for defence signalling and disease transmission in clonal plant networks. Evol. Ecol. 2004, 18, 647–667. [Google Scholar] [CrossRef]
- Gómez, S.; Stuefer, J.F. Members only: Induced systemic resistance to herbivory in a clonal plant network. Oecologia 2006, 147, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Gómez, S.; Onoda, Y.; Ossipov, V.; Stuefer, J.F. Systemic induced resistance: A risk-spreading strategy in clonal plant networks? New Phytol. 2008, 179, 1142–1153. [Google Scholar] [CrossRef]
- Dong, B.C.; Zhang, L.M.; Li, K.Y.; Hu, X.T.; Wang, P.; Wang, Y.J.; Luo, F.L.; Li, H.L.; Yu, F.H. Effects of clonal integration and nitrogen supply on responses of a clonal plant to short-term herbivory. J. Plant Ecol. 2019, 12, 624–635. [Google Scholar] [CrossRef]
- Wu, G.; Wilson, L.T.; McClung, A.M. Contribution of rice tillers to dry matter accumulation and yield. Agron. J. 1998, 90, 317–323. [Google Scholar] [CrossRef]
- Ye, M.; Luo, S.M.; Xie, J.F.; Li, Y.F.; Xu, T.; Liu, Y.; Song, Y.Y.; Zhu-Salzman, K.Y.; Zeng, R.S. Silencing COI1 in rice increases susceptibility to chewing insects and impairs inducible defense. PLoS ONE 2012, 7, e36214. [Google Scholar] [CrossRef]
- Ward, K.A.; Tung, P.; Lamb, N.; Abrams, S.R.; Reid, D.M.; Moloney, M.M.; Holbrook, L.A. Structural requirements for biologically active jasmonates: Induction of protease inhibitors and cotyledon senescence. Plant Growth Regul. 1999, 27, 49–56. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Kiefer, E.; Heller, W.; Ernst, D. A simple and efficient protocol for isolation of functional RNA from plant tissues rich in secondary metabolites. Plant Mol. Biol. Report. 2000, 18, 33–39. [Google Scholar] [CrossRef]
- Ye, M.; Song, Y.; Long, J.; Wang, R.L.; Baerson, S.R.; Pan, Z.Q.; Zhu-Salzman, K.Y.; Xie, J.F.; Cai, K.Z.; Luo, S.M.; et al. Priming of jasmonate-mediated antiherbivore defense responses in rice by silicon. Proc. Natl. Acad. Sci. USA 2013, 110, E3631–E3639. [Google Scholar] [CrossRef] [Green Version]
- Zauberman, G.; Ronen, R.; Akerman, M.; Weksler, A.; Rot, I.; Fuchs, Y. Post-harvest retention of the red colour of litchi fruit pericarp. Sci. Hortic. 1991, 47, 89–97. [Google Scholar] [CrossRef]
- Kraus, T.E.; Fletcher, R.A. Paclobutrazol protects wheat seedlings from heat and paraquat injury. Is detoxification of active oxygen involved? Plant Cell Physiol. 1994, 35, 45–52. [Google Scholar]
- Macri, F.; Braidot, E.; Petrussa, E.; Vianello, A. Lipoxygenase activity associated to isolated soybean plasma membranes. Biochim. Et Biophys. Acta (BBA)-Lipids Lipid Metab. 1994, 1215, 109–114. [Google Scholar] [CrossRef]
- Heinisch, O.; Kowalski, E.; Tauscher, B.; Ludwig, H. Staining for soybean lipoxygenase activity in electrophoretic gels. Eur. J. Lipid Sci. Technol. 1996, 98, 183–184. [Google Scholar] [CrossRef]
- Gómez, S.; Van Dijk, W.; Stuefer, J.F. Timing of induced resistance in a clonal plant network. Plant Biol. 2010, 12, 512–517. [Google Scholar] [CrossRef]
- Karban, R. The ecology and evolution of induced responses to herbivory and how plants perceive risk. Ecol. Entomol. 2020, 45, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Habib, H.; Fazili, K.M. Plant protease inhibitors: A defense strategy in plants. Biotechnol. Mol. Biol. 2007, 2, 68–85. [Google Scholar]
- Liu, Q.S.; Hu, X.Y.; Su, S.L.; Ning, Y.S.; Peng, Y.F.; Ye, G.Y.; Lou, Y.G.; Turlings, T.C.J.; Li, Y.H. Cooperative herbivory between two important pests of rice. Nat. Commun. 2021, 12, 6772. [Google Scholar] [CrossRef]
- Zhuang, Y.Q.; Wang, X.J.; Llorca, L.C.; Lu, J.; Lou, Y.G.; Li, R. Role of jasmonate signaling in rice resistance to the leaf folder Cnaphalocrocis medinalis. Plant Mol. Biol. 2022, 109, 627–637. [Google Scholar] [CrossRef]
- Woldemariam, M.G.; Ahern, K.; Jander, G.; Tzin, V. A role for 9-lipoxygenases in maize defense against insect herbivory. Plant Signal. Behav. 2018, 13, 4709–4723. [Google Scholar] [CrossRef] [Green Version]
- Liao, Z.H.; Wang, L.; Li, C.Z.; Cao, M.J.; Wang, J.N.; Yao, Z.L.; Zhou, S.Y.; Zhou, G.X.; Zhang, D.Y.; Lou, Y.G. The lipoxygenase gene OsRCI-1 is involved in the biosynthesis of herbivore-induced JAs and regulates plant defense and growth in rice. Plant Cell Environ. 2022, 45, 2827–2840. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sun, X. Recent advances in polyphenol oxidase-mediated plant stress responses. Phytochemistry 2021, 181, 112588. [Google Scholar] [CrossRef] [PubMed]
- Mahanil, S.; Attajarusit, J.; Stout, M.J.; Thipyapong, P. Overexpression of tomato polyphenol oxidase increases resistance to common cutworm. Plant Sci. 2008, 174, 456–466. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tong, L.; Wu, W.; Lin, Y.; Chen, D.; Zeng, R.; Lu, L.; Song, Y. Insect Herbivory on Main Stem Enhances Induced Defense of Primary Tillers in Rice (Oryza sativa L.). Plants 2023, 12, 1199. https://doi.org/10.3390/plants12051199
Tong L, Wu W, Lin Y, Chen D, Zeng R, Lu L, Song Y. Insect Herbivory on Main Stem Enhances Induced Defense of Primary Tillers in Rice (Oryza sativa L.). Plants. 2023; 12(5):1199. https://doi.org/10.3390/plants12051199
Chicago/Turabian StyleTong, Lu, Wanghui Wu, Yibin Lin, Daoqian Chen, Rensen Zeng, Long Lu, and Yuanyuan Song. 2023. "Insect Herbivory on Main Stem Enhances Induced Defense of Primary Tillers in Rice (Oryza sativa L.)" Plants 12, no. 5: 1199. https://doi.org/10.3390/plants12051199
APA StyleTong, L., Wu, W., Lin, Y., Chen, D., Zeng, R., Lu, L., & Song, Y. (2023). Insect Herbivory on Main Stem Enhances Induced Defense of Primary Tillers in Rice (Oryza sativa L.). Plants, 12(5), 1199. https://doi.org/10.3390/plants12051199