
Exploring Variability of Free Asparagine Content in the Grain of Bread Wheat (Triticum aestivum L.) Varieties Cultivated in Italy to Reduce Acrylamide-Forming Potential


 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
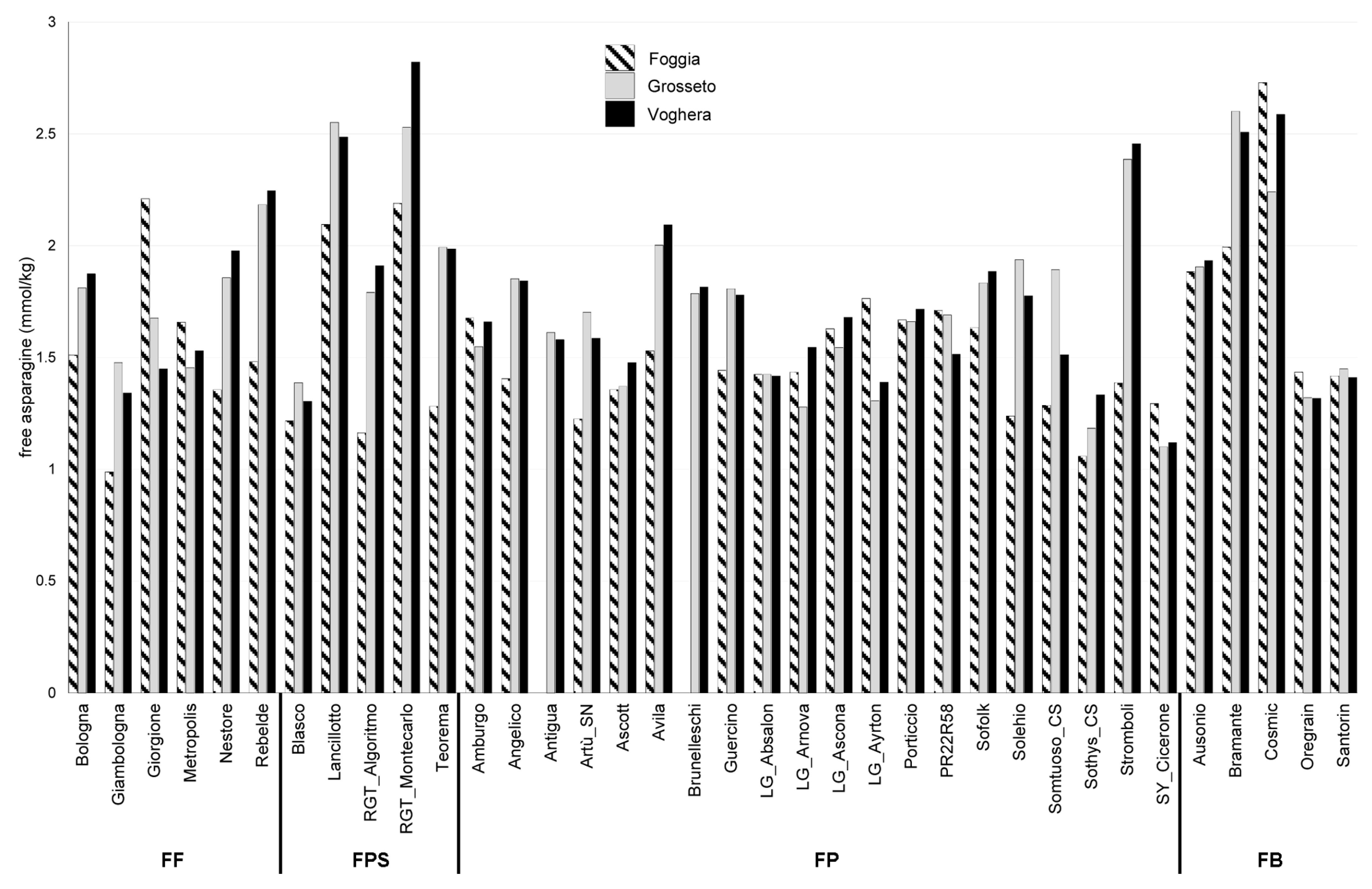
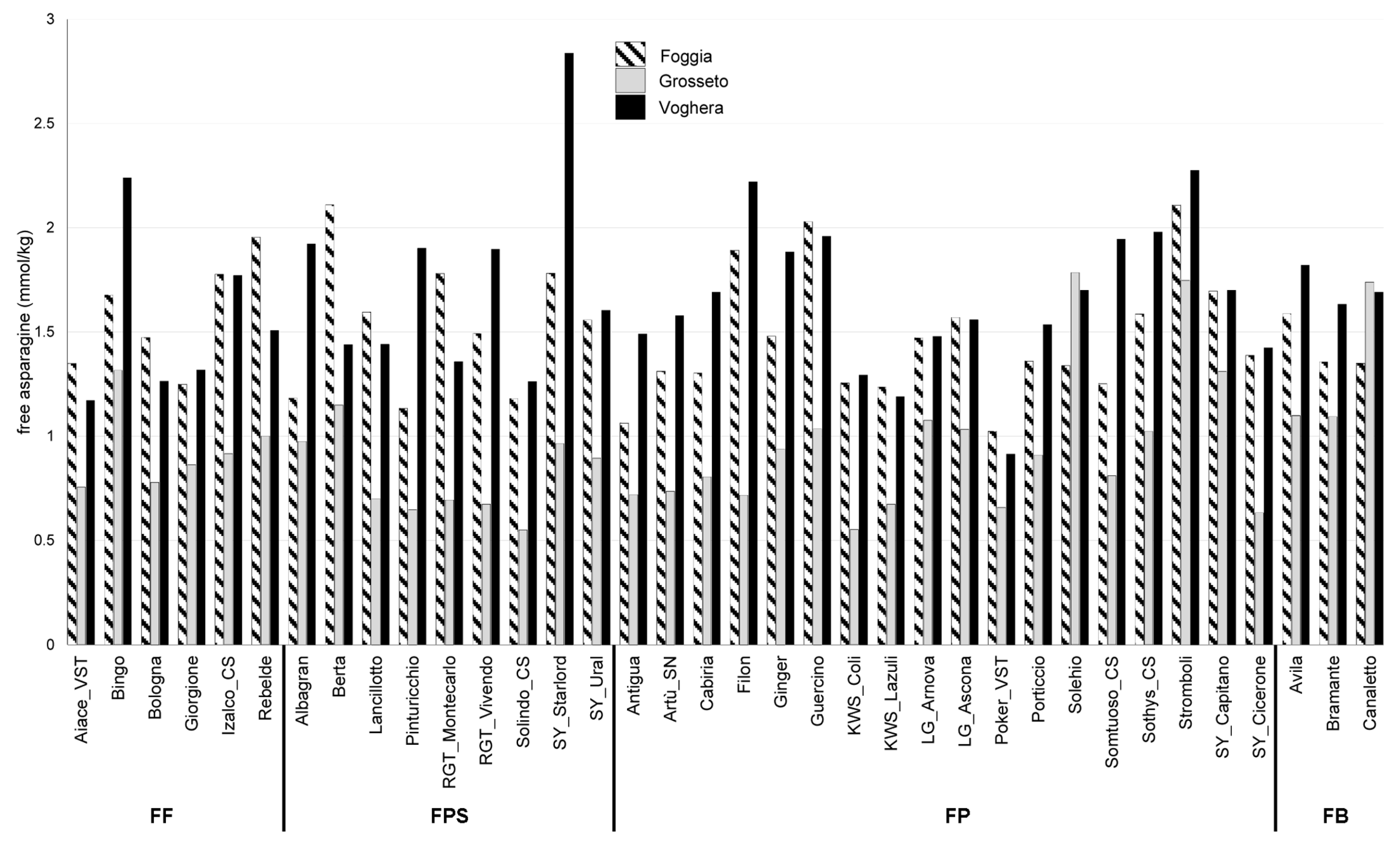
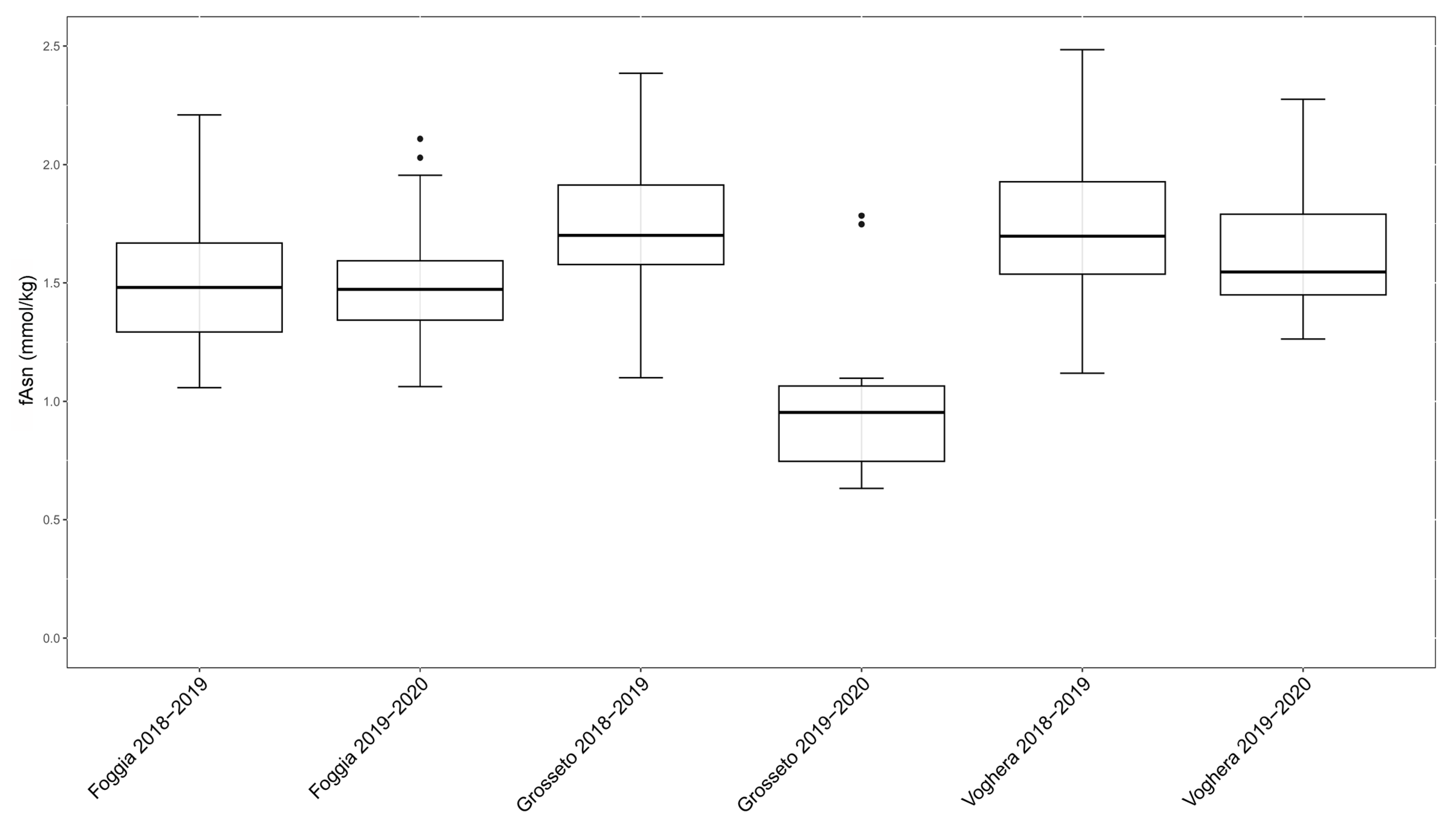
2.1. Free Asparagine Quantification in Six Field Trials Over Two Years
2.2. Evaluation of the Influence of Growing Environments on Free Asparagine Accumulation
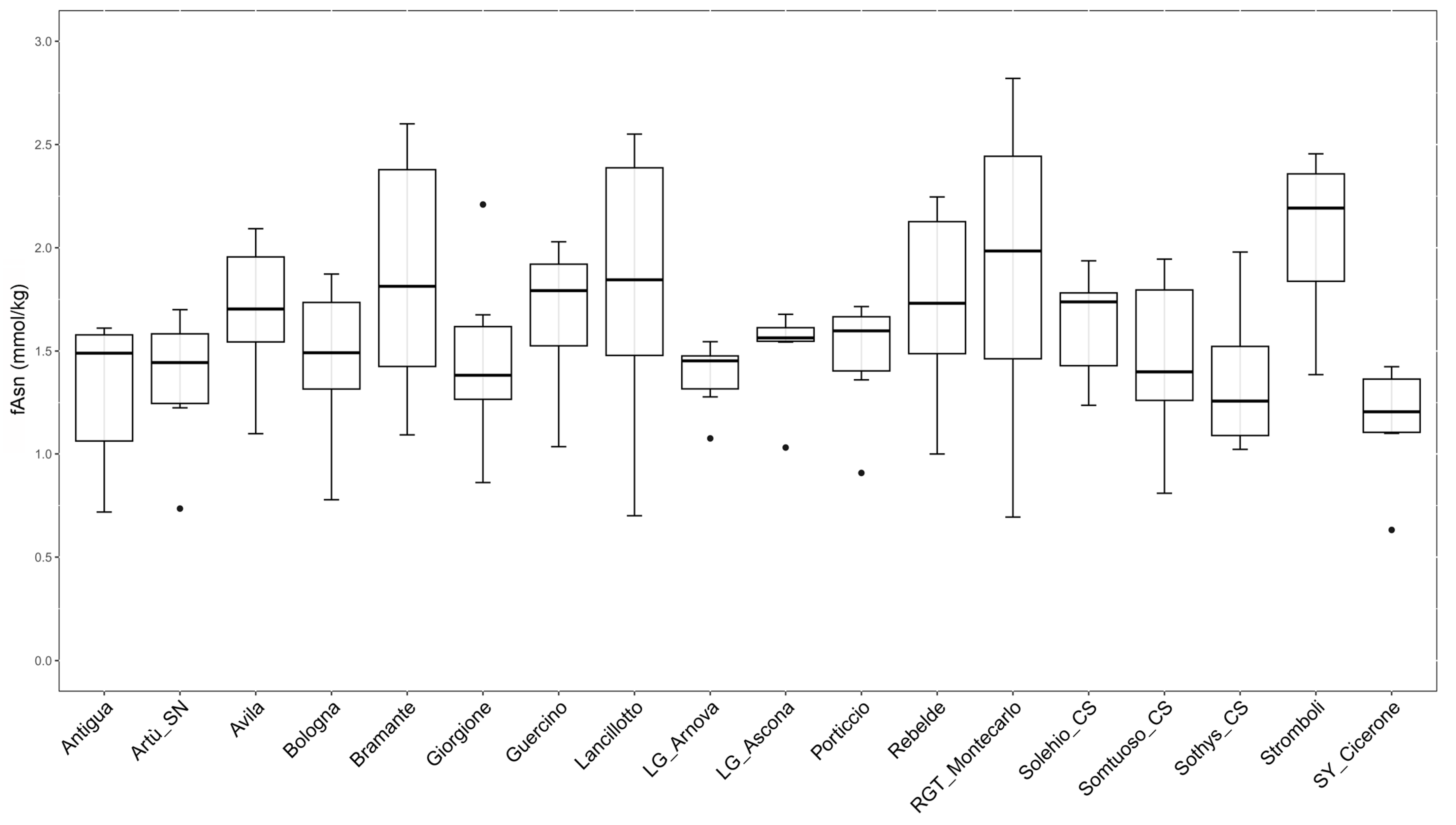
2.3. Evaluation of the Influence of Genetic Factors on Free Asparagine Accumulation
3. Materials and Methods
3.1. Plant Material and Field Trials
3.2. Flour Deproteination
3.3. Free Asparagine Quantification
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kiszonas, A.M.; Morris, C.F. Wheat breeding for quality: A historical review. Cereal Chem. 2018, 95, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Rasheed, A.; Xia, X. From markers to genome-based breeding in wheat. Theor. Appl. Genet. 2019, 132, 767–784. [Google Scholar] [CrossRef] [PubMed]
- Gaikwad, K.B.; Rani, S.; Kumar, M.; Gupta, V.; Babu, P.H.; Bainsla, N.K.; Yadav, R. Enhancing the Nutritional Quality of Major Food Crops Through Conventional and Genomics-Assisted Breeding. Front. Nutr. 2020, 7, 533453. [Google Scholar] [CrossRef] [PubMed]
- Loskutov, I.G.; Khlestkina, E.K. Wheat, barley, and oat breeding for health benefit components in grain. Plants 2021, 10, 86. [Google Scholar] [CrossRef]
- Oddy, J.; Raffan, S.; Wilkinson, M.D.; Elmore, J.S.; Halford, N.G. Understanding the Relationships between Free Asparagine in Grain and Other Traits to Breed Low-Asparagine Wheat. Plants 2022, 11, 669. [Google Scholar] [CrossRef]
- Tareke, E.; Rydberg, P.; Karlsson, P.; Eriksson, S.; Törnqvist, M. Analysis of acrylamide, a carcinogen formed in heated foodstuffs. J. Agric. Food Chem. 2002, 50, 4998–5006. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain. Scientific Opinion on acrylamide in food. EFSA J. 2015, 13, 4104. [Google Scholar] [CrossRef] [Green Version]
- International Agency for Research on Cancer. Some industrial chemicals. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; World Health Organization: Rome, Italy, 1994; Volume 60, pp. 1–568. [Google Scholar]
- Mottram, D.S.; Wedzicha, B.L.; Dodson, A.T. Food chemistry: Acrylamide is formed in the Maillard reaction. Nature 2002, 419, 448–449. [Google Scholar] [CrossRef]
- Stadler, R.H.; Blank, I.; Varga, N.; Robert, F.; Hau, J.; Guy, P.A.; Robert, M.C.; Riediker, S. Food chemistry: Acrylamide from Maillard reaction products. Nature 2002, 419, 449–450. [Google Scholar] [CrossRef]
- Granvogl, M.; Wieser, H.; Koehler, P.; Von Tucher, S.; Schieberle, P. Influence of sulfur fertilization on the amounts of free amino acids in wheat. Correlation with baking properties as well as with 3-aminopropionamide and acrylamide generation during baking. J. Agric. Food Chem. 2007, 55, 4271–4277. [Google Scholar] [CrossRef]
- Raffan, S.; Halford, N.G. Acrylamide in food: Progress in and prospects for genetic and agronomic solutions. Ann. Appl. Biol. 2019, 175, 259–281. [Google Scholar] [CrossRef] [Green Version]
- Curtis, T.Y.; Muttucumaru, N.; Shewry, P.R.; Parry, M.A.J.; Powers, S.J.; Elmore, J.S.; Mottram, D.S.; Hook, S.; Halford, N.G. Effects of genotype and environment on free amino acid levels in wheat grain: Implications for acrylamide formation during processing. J. Agric. Food Chem. 2009, 57, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Halford, N.G.; Muttucumaru, N.; Curtis, T.Y.; Parry, M.A.J. Genetic and agronomic approaches to decreasing acrylamide precursors in crop plants. Food Addit. Contam. 2007, 24, 26–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curtis, T.Y.; Powers, S.J.; Wang, R.; Halford, N.G. Effects of variety, year of cultivation and sulphur supply on the accumulation of free asparagine in the grain of commercial wheat varieties. Food Chem. 2018, 239, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Lea, P.J.; Sodek, L.; Parry, M.A.J.; Shewry, P.R.; Halford, N.G. Asparagine in plants. Ann. Appl. Biol. 2007, 150, 1–26. [Google Scholar] [CrossRef]
- Claus, A.; Schreiter, P.; Weber, A.; Graeff, S.; Herrmann, W.; Claupein, W.; Schieber, A.; Carle, R. Influence of agronomic factors and extraction rate on the acrylamide contents in yeast-leavened breads. J. Agric. Food Chem. 2006, 54, 8968–8976. [Google Scholar] [CrossRef] [PubMed]
- Corol, D.I.; Ravel, C.; Rakszegi, M.; Charmet, G.; Bedo, Z.; Beale, M.H.; Shewry, P.R.; Ward, J.L. 1H-NMR screening for the high-throughput determination of genotype and environmental effects on the content of asparagine in wheat grain. Plant Biotechnol. J. 2016, 14, 128–139. [Google Scholar] [CrossRef]
- Navrotskyi, S.; Baenziger, P.S.; Regassa, T.; Guttieri, M.J.; Rose, D.J. Variation in asparagine concentration in Nebraska wheat. Cereal Chem. 2018, 95, 264–273. [Google Scholar] [CrossRef]
- Curtis, T.Y.; Postles, J.; Halford, N.G. Reducing the potential for processing contaminant formation in cereal products. J. Cereal Sci. 2014, 59, 382–392. [Google Scholar] [CrossRef] [Green Version]
- Martinek, P.; Klem, K.; Váňová, M.; Bartáčková, V.; Večerková, L.; Bucher, P.; Hajšlová, J. Effects of nitrogen nutrition, fungicide treatment and wheat genotype on free asparagine and reducing sugars content as precursors of acrylamide formation in bread. Plant Soil Environ. 2009, 55, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Muttucumaru, N.; Halford, N.G.; Elmore, J.S.; Dodson, A.T.; Parry, M.; Shewry, P.R.; Mottram, D.S. Formation of high levels of acrylamide during the processing of flour derived from sulfate-deprived wheat. J. Agric. Food Chem. 2006, 54, 8951–8955. [Google Scholar] [CrossRef] [PubMed]
- Raffan, S.; Oddy, J.; Halford, N.G. The sulphur response in wheat grain and its implications for acrylamide formation and food safety. Int. J. Mol. Sci. 2020, 21, 3876. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T.L.; Guttieri, M.J.; Nelson, N.O.; Fritz, A.; Tilley, M. Nitrogen and sulfur effects on hard winter wheat quality and asparagine concentration. J. Cereal Sci. 2020, 93, 102969. [Google Scholar] [CrossRef]
- Curtis, T.Y.; Powers, S.J.; Halford, N.G. Effects of Fungicide Treatment on Free Amino Acid Concentration and Acrylamide-Forming Potential in Wheat. J. Agric. Food Chem. 2016, 64, 9689–9696. [Google Scholar] [CrossRef]
- Emebiri, L.C. Genetic variation and possible SNP markers for breeding wheat with low-grain asparagine, the major precursor for acrylamide formation in heat-processed products. J. Sci. Food Agric. 2014, 94, 1422–1429. [Google Scholar] [CrossRef] [PubMed]
- Rapp, M.; Schwadorf, K.; Leiser, W.L.; Würschum, T.; Longin, C.F.H. Assessing the variation and genetic architecture of asparagine content in wheat: What can plant breeding contribute to a reduction in the acrylamide precursor? Theor. Appl. Genet. 2018, 131, 2427–2437. [Google Scholar] [CrossRef] [PubMed]
- Malunga, L.N.; Ames, N.; Khorshidi, A.S.; Thandapilly, S.J.; Yan, W.; Dyck, A.; Waterer, J.; Malcolmson, L.; Cuthbert, R.; Sopiwnyk, E.; et al. Association of asparagine concentration in wheat with cultivar, location, fertilizer, and their interaction. Food Chem. 2021, 344, 128630. [Google Scholar] [CrossRef]
- Postles, J.; Powers, S.J.; Elmore, J.S.; Mottram, D.S.; Halford, N.G. Effects of variety and nutrient availability on the acrylamide-forming potential of rye grain. J. Cereal Sci. 2013, 57, 463–470. [Google Scholar] [CrossRef] [Green Version]
- Cocchi, M.; Corbellini, M.; Foca, G.; Lucisano, M.; Pagani, M.A.; Tassi, L.; Ulrici, A. Classification of bread wheat flours in different quality categories by a wavelet-based feature selection/classification algorithm on NIR spectra. Anal. Chim. Acta 2005, 544, 100–107. [Google Scholar] [CrossRef]
- Foca, G.; Ulrici, A.; Corbellini, M.; Pagani, M.A.; Lucisano, M.; Franchini, G.C.; Tassi, L. Reproducibility of the Italian ISQ method for quality classification of bread wheats: An evaluation by expert assessors. J. Sci. Food Agric. 2007, 87, 839–846. [Google Scholar] [CrossRef]
- Lecart, B.; Jacquet, N.; Anseeuw, L.; Renier, M.; Njeumen, P.; Bodson, B.; Vanderschuren, H.; Richel, A. Nonconventional enzymatic method to determine free asparagine level in whole-grain wheat. Food Chem. 2018, 251, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Žilić, S.; Dodig, D.; Basić, Z.; Vančetović, J.; Titan, P.; Đurić, N.; Tolimir, N. Free asparagine and sugars profile of cereal species: The potential of cereals for acrylamide formation in foods. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2017, 34, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Ohm, J.B.; Mergoum, M.; Simsek, S. Variation of free asparagine concentration and association withquality parameters for hard red spring wheat grown in North Dakota. Cereal Chem. 2017, 94, 712–716. [Google Scholar] [CrossRef]
- Xie, Y.; Malunga, L.N.; Ames, N.P.; Waterer, J.; Khorshidi, A.S.; Scanlon, M.G. Effects of growing environment, genotype, and commercial fertilization levels on free asparagine concentration in Western Canadian wheat. Cereal Chem. 2021, 98, 89–99. [Google Scholar] [CrossRef]
- Oddy, J.; Alarcón-Reverte, R.; Wilkinson, M.; Ravet, K.; Raffan, S.; Minter, A.; Mead, A.; Elmore, J.S.; de Almeida, I.M.; Cryer, N.C.; et al. Reduced free asparagine in wheat grain resulting from a natural deletion of TaASN-B2: Investigating and exploiting diversity in the asparagine synthetase gene family to improve wheat quality. BMC Plant Biol. 2021, 21, 302. [Google Scholar] [CrossRef]
- Žilić, S.; Aktağ, I.G.; Dodig, D.; Filipović, M.; Gökmen, V. Acrylamide formation in biscuits made of different wholegrain flours depending on their free asparagine content and baking conditions. Food Res. Int. 2020, 132, 109109. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Italian Bread Wheat Classification | Genotype Name | Field Trials |
|---|---|---|
| FF | Aiace_VST | 2019–2020 |
| Bingo_VST | 2019–2020 | |
| Bologna | both years | |
| Giambologna | 2018–2019 | |
| Giorgione | both years | |
| Izalco_CS | 2019–2020 | |
| Metropolis | 2018–2019 | |
| Nestore | 2018–2019 | |
| Rebelde | both years | |
| FPS | Albagran | 2019–2020 |
| Berta | 2019–2020 | |
| Blasco | 2018–2019 | |
| Lancillotto | both years | |
| Pinturicchio | 2019–2020 | |
| RGT_Algoritmo | 2018–2019 | |
| RGT_Montecarlo | both years | |
| RGT_Vivendo | 2019–2020 | |
| Solindo_CS | 2019–2020 | |
| SY_Starlord | 2019–2020 | |
| SY_Ural | 2019–2020 | |
| Teorema | 2018–2019 | |
| FP | Amburgo | 2018–2019 |
| Angelico | 2018–2019 | |
| Antigua | both years | |
| Artù_SN | both years | |
| Ascott | 2018–2019 | |
| Ausonio | 2018–2019 | |
| Brunelleschi | 2018–2019 | |
| Cabiria | 2019–2020 | |
| Somtuoso_CS | both years | |
| Filon | 2019–2020 | |
| Ginger | 2019–2020 | |
| Guercino | both years | |
| KWS_Coli | 2019–2020 | |
| KWS_Lazulli | 2019–2020 | |
| LG_Absalon | 2018–2019 | |
| LG_Arnova | both years | |
| LG_Ascona | both years | |
| LG_Ayrton | 2018–2019 | |
| Poker_VST | 2019–2020 | |
| Porticcio | both years | |
| PR22R58 | 2018–2019 | |
| Sofolk | 2018–2019 | |
| Solehio | both years | |
| Sothys_CS | both years | |
| Stromboli | both years | |
| SY_Capitano | 2019–2020 | |
| SY_Cicerone | both years | |
| FB | Avila | both years |
| Bramante | both years | |
| Canaletto | 2019–2020 | |
| Cosmic | 2018–2019 | |
| Oregrain | 2018–2019 | |
| Santorin | 2018–2019 |
| Year | Location | Field Coordinates | Sowing Date | Harvesting Date |
|---|---|---|---|---|
| 2018/2019 | Foggia | 41°27′45.26” N, 15°30′11.06” E | 6 December 2018 | 18 June 2019 |
| 2019/2020 | Foggia | 41°27′46.03” N, 15°30′10.02” E | 9 December 2019 | 11 June 2020 |
| 2018/2019 | Grosseto | 42°57′14.332” N, 11°5′45.956” E | 3 January 2019 | 2 July 2019 |
| 2019/2020 | Grosseto | 42°57′26.557” N, 11°5′47.68” E | 5 January 2020 | 24 June 2020 |
| 2018/2019 | Voghera | 45°1′44.83” N, 9°1′19.189” E | 25 October 2018 | 20 July 2019 |
| 2019/2020 | Voghera | 45°1′0.3” N, 8°59′44.822” E | 9 December 2019 | 12 July 2020 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tafuri, A.; Zuccaro, M.; Ravaglia, S.; Pirona, R.; Masci, S.; Sestili, F.; Lafiandra, D.; Ceriotti, A.; Baldoni, E. Exploring Variability of Free Asparagine Content in the Grain of Bread Wheat (Triticum aestivum L.) Varieties Cultivated in Italy to Reduce Acrylamide-Forming Potential. Plants 2023, 12, 1349. https://doi.org/10.3390/plants12061349
Tafuri A, Zuccaro M, Ravaglia S, Pirona R, Masci S, Sestili F, Lafiandra D, Ceriotti A, Baldoni E. Exploring Variability of Free Asparagine Content in the Grain of Bread Wheat (Triticum aestivum L.) Varieties Cultivated in Italy to Reduce Acrylamide-Forming Potential. Plants. 2023; 12(6):1349. https://doi.org/10.3390/plants12061349
Chicago/Turabian StyleTafuri, Andrea, Melania Zuccaro, Stefano Ravaglia, Raul Pirona, Stefania Masci, Francesco Sestili, Domenico Lafiandra, Aldo Ceriotti, and Elena Baldoni. 2023. "Exploring Variability of Free Asparagine Content in the Grain of Bread Wheat (Triticum aestivum L.) Varieties Cultivated in Italy to Reduce Acrylamide-Forming Potential" Plants 12, no. 6: 1349. https://doi.org/10.3390/plants12061349
APA StyleTafuri, A., Zuccaro, M., Ravaglia, S., Pirona, R., Masci, S., Sestili, F., Lafiandra, D., Ceriotti, A., & Baldoni, E. (2023). Exploring Variability of Free Asparagine Content in the Grain of Bread Wheat (Triticum aestivum L.) Varieties Cultivated in Italy to Reduce Acrylamide-Forming Potential. Plants, 12(6), 1349. https://doi.org/10.3390/plants12061349