
Allelopathic Activity of Canadian Goldenrod (Solidago canadensis L.) Extracts on Seed Germination and Growth of Lettuce (Lactuca sativa L.) and Garden Pepper Cress (Lepidium sativum L.)

 ,
,  ,
,
Abstract
:1. Introduction
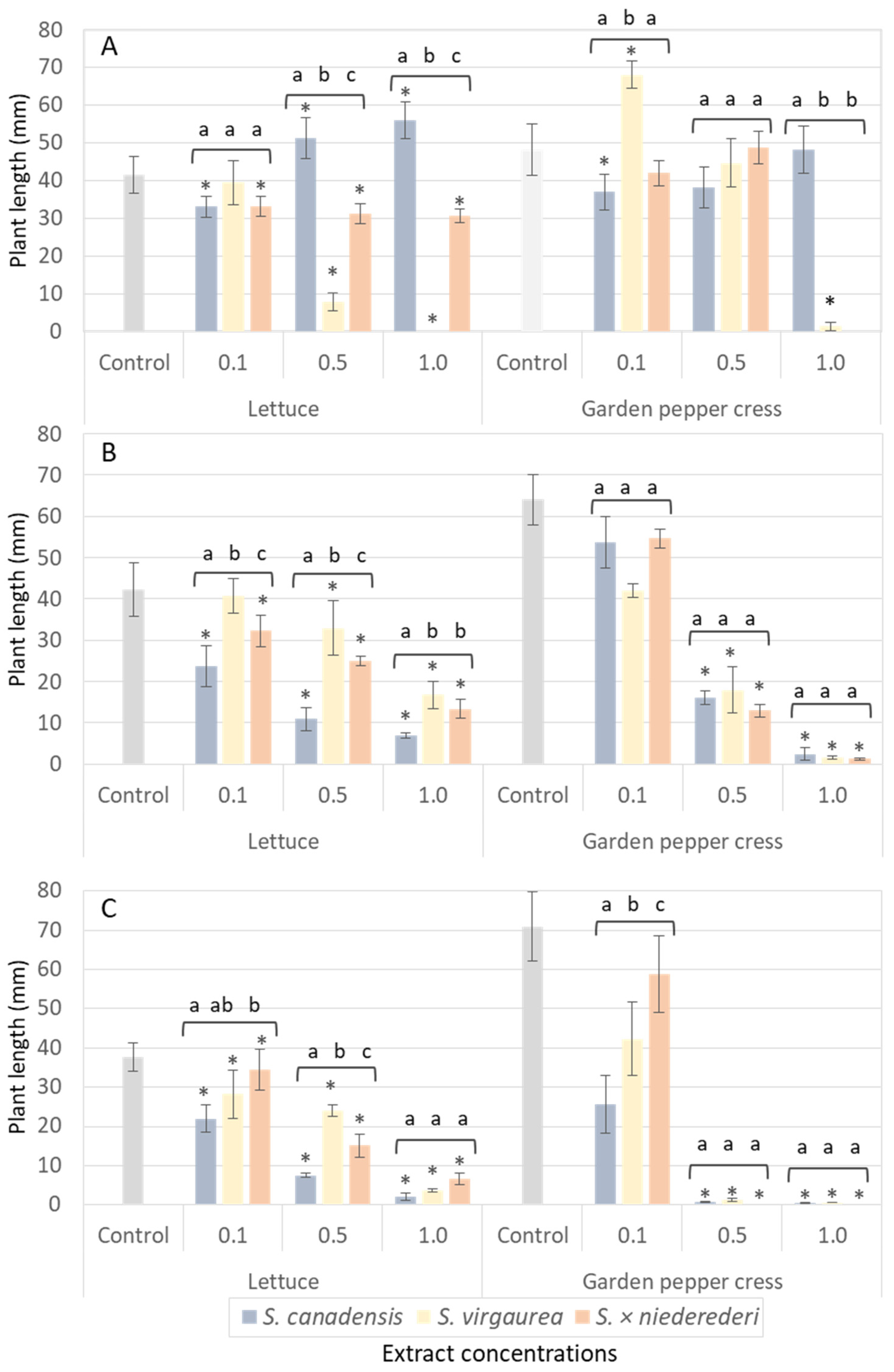
2. Results
2.1. Soil Characteristics (Conductivity, pH and Major Elements)
2.2. Chemical Composition of Solidago canadensis Methanol/Water Extracts
2.3. Chemical Composition of Volatile Organic Compounds (VOCs) in Solidago canadensis Water/Diethyl Ether Extracts
2.4. Chemical Composition of Solidago canadensis Essential Oils (EOs)
2.5. Allelopathic Effects of Solidago canadensis Water/Diethyl Ether Extracts
2.6. Relative Germination (RG) and Vigor Index (VI) of Lettuce (Lactuca sativa L.) and Garden Pepper Cress (Lepidium sativum L.) Affected by S. canadensis, S. virgaurea and S. × niederederi Extracts
2.7. Allelopathic Effects of Water Extracts of Various Solidago Species: S. canadensis, S. virgaurea and S. × niederederi
2.8. Inhibitory Effect of Solidago canadensis EOs on Seed Germination and Seedling Growth of Lettuce (Lactuca sativa) and Garden Pepper Cress (Lepidium sativum)
3. Discussion
4. Materials and Methods
4.1. Soil Analysis
4.2. Plant Material
4.3. Preparation of Various Plant Extracts
4.3.1. Essential Oil Isolation
4.3.2. Preparation of S. canadensis, S. virgaurea, S. × niederederi Extracts for Allelopathic Tests
4.3.3. Preparation of S. canadensis Extracts for HPLC-DAD-TOF Analysis
4.4. Gas Chromatographic Analysis of Solidago canadensis EOs and Extracts (Water/Diethyl Ether)
4.4.1. GC (Flame-Ionization Detector FID) Analysis
4.4.2. GC-MS Analysis
4.4.3. Identification of Individual Components
4.5. HPLC-DAD-MS (TOF) Analysis Solidago canadensis Extracts
4.6. Allelopathy of Solidago canadensis EOs
4.7. Bioassay for Seed Germination
4.8. Seedling Root Length (RL), Seedling Height (H), Germination Rate (GR), Relative Germination (RG), and Vigor Index (VI) Measurements
4.9. Statistical Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Semple, J.C.; Beck, J.B. A revised infrageneric classification of Solidago (Asteraceae: Astereae). Phytoneuron 2021, 10, 1–6. [Google Scholar]
- Walck, J.L.; Baskin, J.M.; Baskin, C.C. Relative competitive abilities and growth characteristics of a narrowly endemic and a geographically widespread Solidago species (Asteraceae). Am. J. Bot. 1999, 86, 820–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šutovská, M.; Capek, P.; Kocmálová, M.; Fraňová, S.; Pawlaczyk, L.; Gancarz, R. Characterization and biological activity of Solidago canadensis complex. Int. J. Biol. Macromol. 2013, 52, 192–197. [Google Scholar] [CrossRef]
- Grul’ová, D.; Pl’uchtová, M.; Fejér, J.; De Martino, L.; Caputo, L.; Sedlák, V.; De Feo, V. Influence of the six essential oils on invasive Solidago canadensis L. seed germination. Nat. Prod. Res. 2020, 34, 3231–3233. [Google Scholar] [CrossRef]
- Pliszko, A.; Zalewska-Gałosz, J. Molecular evidence for hybridization between invasive Solidago canadensis and native S. virgaurea. Biol. Invasions 2016, 18, 3103–3108. [Google Scholar] [CrossRef] [Green Version]
- Skokanová, K.; Mereďa, P.; Šingliarová, B.; Španiel, S. Lectotype of Solidago × niederederi (Asteraceae) selected from recently rediscovered original material. Phytotaxa 2020, 438, 62–64. [Google Scholar] [CrossRef]
- Gudžinskas, Z.; Žalneravičius, E. Solidago × snarskisii nothosp. nov. (Asteraceae) from Lithuania and its position in the infrageneric classification of the genus. Phytotaxa 2016, 253, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Karpavičienė, B.; Radušienė, J. Morphological and anatomical characterization of Solidago × niederederi and other sympatric Solidago species. Weed Sci. 2016, 64, 61–70. [Google Scholar] [CrossRef]
- Musiał, K.; Pagitz, K.; Gudžinskas, Z.; Łazarski, G.; Pliszko, A. Chromosome numbers in hybrids between invasive and native Solidago (Asteraceae) species in Europe. Phytotaxa 2020, 471, 267–275. [Google Scholar] [CrossRef]
- Anonymous. Regarding the Amendment of the Minister of the Environment of the Republic of Lithuania “Regarding the Approval of the List of Species of Invasive Organisms in Lithuania and the Recognition of Some Orders of the Minister of the Environment as Having Lost Their Validity”. Available online: https://e-seimas.lrs.lt/portal/legalAct/lt/TAD/0754e1b0b56011e6a3e9de0fc8d85cd8 (accessed on 16 August 2004).
- Królak, E. Negative and positive aspects of the presence of Canadian goldenrod in the environment. Environ. Prot. Nat. Resour. 2021, 32, 6–12. [Google Scholar] [CrossRef]
- Li, G.; Wang, J.; Zhang, J.; Li, Y.; Liu, E.; Yu, Y.; Iqbal, B.; Dai, Z.; Jia, H.; Li, J.; et al. Effects of experimental warming and Canada goldenrod invasion on the diversity and function of the soil nematode community. Sustainability 2021, 13, 13145. [Google Scholar] [CrossRef]
- Kalemba, D.; Góra, J.; Kurowska, A. Analysis of the essential oil of Solidago canadensis. Planta Med. 1990, 56, 222–223. [Google Scholar] [CrossRef] [PubMed]
- Weyerstahl, P.; Marschall, H.; Christiansen, C.; Kalemba, D.; Góra, J. Constituents of the essential oil of Solidago canadensis (“Goldenrod”) from Poland—A correction. Planta Med. 1993, 59, 281–282. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.X.; He, W.; Wen, G.Y. The constituents of essential oil from Solidago canadensis. Chin. Bull. Bot. 1999, 16, 178–180. [Google Scholar]
- Kalemba, D.; Thiem, B. Constituents of the essential oils of four micropropagated Solidago species. Flavour Fragr. J. 2004, 19, 40–43. [Google Scholar] [CrossRef]
- Kasalia, A.A.; Ekundayo, O.; Paul, C.; König, W.A. epi-Cubebanes from Solidago canadensis. Phytochemistry 2002, 59, 805–810. [Google Scholar] [CrossRef]
- Li, D.Q.; Pan, S.H.; Zhu, X.W.; Tan, L.; Cao, Y.F. Anticancer activity and chemical composition of leaf essential oil from Solidago canadensis L. in China. Adv. Mat. Res. 2012, 347, 1584–1589. [Google Scholar]
- Huang, B.K.; Lei, Y.L.; Qin, L.P.; Liu, J. Chemical composition and cytotoxic activities of the essential oil from inflorescences of Solidago canadensis L., an invasive weed in Southeastern China. J. Essent. Oil-Bear. Plants 2012, 15, 667–671. [Google Scholar] [CrossRef]
- El-Sherei, M.; Khaleel, A.; Motaal, A.A.; Abd-Elbaki, P. Effect of seasonal variation on the composition of the essential oil of Solidago canadensis cultivated in Egypt. J. Essent. Oil-Bear. Plants 2014, 17, 891–898. [Google Scholar] [CrossRef]
- Xiwu, Z.; Peng, X.; Yuefen, C.; Jicai, T.; Haimin, C.; Jishuang, C. Comparison of chemical constituents and antimicrobial activities of volatile oil from Solidago decurrens and S. canadensis. Sci. Silvae Sin. 2009, 12, 167. [Google Scholar]
- Zhang, J.; Li, B.; Chen, J.; Zhout, T. Chemical constituents and antimicrobial activity of volatile oil from Solidago canadensis L. J. Fudan Univ. Nat. Sci. 2006, 3, 412–416. [Google Scholar]
- Liu, S.; Shao, X.; Wei, Y.; Li, Y.; Xu, F.; Wang, H. Solidago canadensis L. essential oil vapor effectively inhibits Botrytis cinerea growth and preserves postharvest quality of strawberry as a food model system. Front Microbiol. 2016, 7, 1179. [Google Scholar] [CrossRef] [PubMed]
- Grul’ová, D.; Baranová, B.; Ivanova, V.; De Martino, L.; Mancini, E.; De Feo, V. Composition and bioactivity of EOs of Solidago spp. and their impact on radish and garden cress. Allelopath. J. 2016, 39, 129–142. [Google Scholar]
- Synowiec, A.; Kalemba, D.; Drozdek, E.; Bocianowski, J. Phytotoxic potential of EOs from temperate climate plants against the germination of selected weeds and crops. J. Pest Sci. 2017, 90, 407–419. [Google Scholar] [CrossRef]
- Shelepova, O.; Vinogradova, Y.; Zaitchik, B.; Ruzhitsky, A.; Grygorieva, O.; Brindza, J. Constituents of the essential oil in Solidago canadensis L. from Eurasia. Potravin. Slovak J. Food Sci. 2018, 12, 20–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elshafie, H.S.; Grul’ová, D.; Baranová, B.; Caputo, L.; De Martino, L.; Sedlák, V.; Camele, I.; De Feo, V. Antimicrobial activity and chemical composition of essential oil extracted from Solidago canadensis L. growing wild in Slovakia. Molecules 2019, 24, 1206. [Google Scholar] [CrossRef] [Green Version]
- Benelli, G.; Pavela, R.; Cianfaglione, K.; Nagy, D.U.; Canale, A.; Maggi, F. Evaluation of two invasive plant invaders in Europe (Solidago canadensis and Solidago gigantea) as possible sources of botanical insecticides. J. Pest Sci. 2019, 92, 805–821. [Google Scholar] [CrossRef]
- Wandjou, J.G.N.; Quassinti, L.; Gudžinskas, Z.; Nagy, D.U.; Cianfaglione, K.; Bramucci, M.; Maggi, F. Chemical composition and antiproliferative effect of EOs of four Solidago species (S. canadensis, S. gigantea, S. virgaurea and S. niederederi). Chem. Biodivers. 2020, 17, e2000685. [Google Scholar]
- Radušienė, J.; Karpavičienė, B.; Marksa, M.; Ivanauskas, L.; Raudonė, L. Distribution patterns of essential oil terpenes in native and invasive Solidago species and their comparative assessment. Plants 2022, 11, 1159. [Google Scholar] [CrossRef]
- Mishra, D.; Joshi, S.; Bisht, G.; Pilkhwal, S. Chemical composition and antimicrobial activity of Solidago canadensis Linn. root essential oil. J. Basic Clin. Physiol. Pharmacol. 2010, 1, 187–190. [Google Scholar]
- Apati, P.; Szentmihalyi, K.; Balazs, A.; Baumann, D.; Hamburger, M.; Kristo, T.S.; Szoke, E.; Kery, A. HPLC analysis of the flavonoids in pharmaceutical preparations from Canadian goldenrod (Solidago canadensis). Chromatographia 2002, 56, 65–68. [Google Scholar] [CrossRef]
- Reznicek, G.; Jurenitsch, J.; Kubelka, W.; Korhammer, S.; Haslinger, E.; Hiller, K. The first spectroscopically confirmed saponins from Solidago canadensis. Planta Med. 1990, 56, 554. [Google Scholar] [CrossRef]
- Reznicek, G.; Jurenitsch, J.; Plasun, M.; Korhammer, S.; Haslinger, E.; Hiller, K.; Kubelka, W. Four major saponins from Solidago canadensis. Phytochemistry 1991, 30, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Radušienė, J.; Marksa, M.; Karpavičienė, B. Assessment of Solidago × niederederi origin based on the accumulation of phenolic compounds in plant raw materials. Weed Sci. 2018, 66, 324–330. [Google Scholar] [CrossRef]
- Radušienė, J.; Marksa, M.; Ivanauskas, L.; Jakstas, V.; Karpavičienė, B. Assessment of phenolic compound accumulation in two widespread goldenrods. Ind. Crops Prod. 2015, 63, 158–166. [Google Scholar] [CrossRef]
- Lu, T.; Menelaou, M.A.; Vargas, D.; Fronczek, F.R.; Fischer, N.H. Polyacetylenes and diterpenes from Solidago canadensis. Phytochemistry 1993, 32, 1483–1488. [Google Scholar]
- Bradette-Hebert, M.-E.; Legault, J.; Lavoie, S.; Pichette, A. A new labdane diterpene from the flowers of Solidago canadensis. Chem. Pharm. Bull. 2008, 56, 82–84. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhang, X.; Lei, G.; Li, B.; Chen, J.; Zhout, T. A new phenolic glycoside from the aerial parts of Solidago canadensis. Fitoterapia 2007, 78, 69–71. [Google Scholar] [CrossRef]
- Marksa, M.; Zymone, K.; Ivanauskas, L.; Radušienė, J.; Pukalskas, A.; Raudonė, L. Antioxidant profiles of leaves and inflorescences of native, invasive and hybrid Solidago species. Ind. Crops Prod. 2020, 145, 112123. [Google Scholar] [CrossRef]
- Kołodziej, B.; Kowalski, R.; Kędzia, B. Antibacterial and antimutagenic activity of extracts above ground parts of three Solidago species: Solidago virgaurea L., Solidago canadensis L. and Solidago gigantean Ait. J. Med. Plant Res. 2011, 5, 6770–6779. [Google Scholar]
- Chaturvedula, V.S.P.; Zhou, B.N.; Gao, Z.; Thomas, S.J.; Hecht, S.M.; Kingston, D.G.I. New lupane triterpenoids from Solidago canadensis that inhibit the lease activity of DNA polymerase β. Bioorg. Med. Chem. 2004, 12, 6271–6275. [Google Scholar] [CrossRef]
- Baki, P.M.A.; El-Sherei, M.M.; Khaleel, A.E.; Motaal, A.A.A.; Abdallahc, H.M.I. Aquaretic activity of Solidago canadensis L. cultivated in Egypt and determination of the most bioactive fraction. Iran. J. Pharm. Res. 2019, 18, 922–937. [Google Scholar] [PubMed]
- Ravichandiran, V.; Deepa, N. In-vitro anticancer activity of Solidago canadensis L. Int. J. Res. Pharm. Sci. 2012, 3, 158–162. [Google Scholar]
- Deng, Y.; Zhao, Y.; Padilla-Zakourc, O.; Yanga, G. Polyphenols, antioxidant and antimicrobial activities of leaf and bark extracts of Solidago canadensis L. Ind. Crops Prod. 2015, 74, 803–809. [Google Scholar] [CrossRef]
- Apáti, P.; Kéry, Á.; Houghton, P.J.; Steventon, G.B.; Kite, G. In-vitro effect of flavonoids from Solidago canadensis extract on glutathione S-transferase. J. Pharmacol. Pharm. 2006, 58, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Kelly, A.M.; de Oliveira, T.B.; Valverde, S.S. Determination of the metabolic profile of Solidago canadensis using UFLC-PDA-ESI-TOF. Rodriguésia 2020, 71, e01062019. [Google Scholar] [CrossRef]
- Zekič, J.; Vovk, I.; Glavnik, V. Extraction and analyses of flavonoids and phenolic acids from Canadian goldenrod and Giant goldenrod. Forests 2021, 12, 40. [Google Scholar] [CrossRef]
- Likhanov, A.; Oliinyk, M.; Pashkevych, N.; Churilov, A.; Kozyr, M. The role of flavonoids in invasion strategy of Solidago canadensis L. Plants 2021, 10, 1748. [Google Scholar] [CrossRef]
- Yuan, Y.; Wang, B.; Zhang, S.; Tang, J.; Tu, C.; Hu, S.; Yong, J.W.H.; Chen, X. Enhanced allelopathy and competitive ability of invasive plant Solidago canadensis in its introduced range. J. Plant Ecol. 2013, 6, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Abhilasha, D.; Quintana, N.; Vivanco, J.; Joshi, J. Do allelopathic compounds in invasive Solidago canadensis s. l. restrain the native European flora? J. Ecol. 2008, 96, 993–1001. [Google Scholar] [CrossRef]
- Wang, C.; Xiao, H.; Zhao, L.; Liu, J.; Wang, L.; Zhang, F.; Shi, Y.; Du, D. The allelopathic effects of invasive plant Solidago canadensis on seed germination and growth of Lactuca sativa enhanced by different types of acid deposition. Ecotoxicology 2016, 25, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Zandi, P.; Barabasz-Krasny, B.; Stachurska-Swakńo, A.; Puła, J.; Możdżeń, K. Allelopathic effect of invasive Canadian goldenrod (Solidago canadensis L.) on early growth of red clover (Trifolium pratense L.). Not. Bot. Horti Agrobot. Cluj Napoca 2020, 48, 2060–2071. [Google Scholar] [CrossRef]
- Możdżeń, K.; Barabasz-Krasny, B.; Zandi, P.; Kliszcz, A.; Puła, J. Effect of aqueous extracts from Solidago canadensis L. leaves on germination and early growth stages of three cultivars of Raphanus sativus L. var. radicula Pers. Plants 2020, 9, 1549. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Tan, J.; Wan, Z.; Gu, F.; Zhu, M. Allelopathic effects of extracts from Solidago canadensis L. against seed germination and seedling growth of some plants. J. Environ. Sci. 2006, 18, 304–309. [Google Scholar]
- Li, S.L.; Li, Z.H.; Wang, Y.F.; Ruan, X.; Pan, C.D.; Wang, Q. Preliminary study for the allelopathic effect of water extracts from Solidago canadensis leaves. Adv. Mat. Res. 2013, 699, 340–349. [Google Scholar]
- Wang, K.J.; Chen, L.Z.; Yu, X.P. Preliminary study of allelopathy of Solidago canadensis L. Acta Agric. Zhejiangensis 2006, 18, 299–303. [Google Scholar]
- Baležentienė, L. Secondary metabolite accumulation and phytotoxicity of invasive species Solidago canadensis L. during the growth period. Allelopath. J. 2015, 35, 217–226. [Google Scholar]
- Gala-Czekaj, D.; Dziurka, M.; Bocianowski, J.; Synowiec, A. Autoallelopathic potential of aqueous extracts from Canadian goldenrod (Solidago canadensis L.) and giant goldenrod (S. gigantea Aiton). Acta Physiol. Plant. 2022, 44, 1. [Google Scholar] [CrossRef]
- Karpavičienė, B.; Danilovienė, J.; Vykertaitė, R. Congeneric comparison of allelopathic and autotoxic effects of four Solidago species. Bot. Serb. 2019, 43, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Adams, R.P. Essential Oil Components by Quadrupole Gas Chromatography/Mass Spectrometry, 3rd ed.; Allured Publishing Corp.: Carol Stream, IL, USA, 2001. [Google Scholar]
- Chytrý, M.; Tichý, L.; Hennekens, S.M.; Knollová, I.; Janssen, J.A.M.; Rodwell, J.S.; Peterka, T.; Marcenò, C.; Landucci, F.; Danihelka, J.; et al. ·EUNIS Habitat Classification: Expert system, characteristic species combinations and distribution maps of European habitats. Appl. Veg. Sci. 2020, 23, 648–675. [Google Scholar] [CrossRef]
- Dong, L.J.; Wei-Ming, H. The relative contributions of climate, soil, diversity and interactions to leaf trait variation and spectrum of invasive Solidago canadensis. BMC Ecol. 2019, 19, 24. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.J.; Sun, Z.K.; Gao, Y.; He, W.M. Two-year interactions between invasive Solidago canadensis and soil decrease its subsequent growth and competitive ability. J. Plant Ecol. 2015, 8, 617–622. [Google Scholar] [CrossRef] [Green Version]
- Baranová, B.; Troščáková-Kerpčárová, E.; Grul’ová, D. Survey of the Solidago canadensis L. morphological traits and essential oil production: Aboveground biomass growth and abundance of the invasive goldenrod appears to be reciprocally enhanced within the invaded stands. Plants 2022, 11, 535. [Google Scholar] [CrossRef]
- Anonymous. Regarding the Approval of the Lithuanian Hygiene Norm HN 60:2004 “Maximum Permissible Concentration of Dangerous Chemical Substances in Soil”. Available online: https://e-seimas.lrs.lt/portal/legalAct/lt/TAD/TAIS.228693/asr (accessed on 8 March 2004).
- Sako, Y.; McDonald, M.B.; Fujimura, K.; Rvans, A.F.; Bennett, M.M. A system for automated seed vigour assessment. Seed Sci. Technol. 2001, 29, 625–636. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | Conductivity, µS/cm | pH Values |
|---|---|---|
| I | 99.03 ± 0.40–103.83 ± 1.11 | 6.15 ± 0.07 |
| II | 99.07 ± 2.82–116.57 ± 10.10 | 6.48 ± 0.19 |
| III | 92.33 ± 0.55–125.63 ± 6.04 | 6.65 ± 0.27 |
| IV | 95.02 ± 0.55–115.50 ± 2.02 | 6.45 ± 0.11 |
| Sampling Sites | Ca | Mg | K | Na | Al | Mn | Cu | Cd | Cr | Ni | Pb | Zn | Fe | Mo | P |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| λ, nm | 317.93 | 285.21 | 766.49 | 589.59 | 396.15 | 257.61 | 327.39 | 228.80 | 267.72 | 231.60 | 220.35 | 213.86 | 238.20 | 202.03 | 231.67 |
| I | 1.232 | 0.391 | 0.132 | 0.009 | 2.257 | 0.577 | 0.023 | 0.003 | 0.003 | 0.001 | 0.007 | 0.033 | 3.513 | 0.002 | 0.412 |
| SD | 0.130 | 0.044 | 0.025 | 0.007 | 0.227 | 0.119 | 0.005 | 0.001 | 0.002 | 0.000 | 0.001 | 0.001 | 0.483 | 0.002 | 0.039 |
| II | 3.098 | 0.587 | 0.152 | 0.001 | 2.148 | 0.489 | 0.020 | 0.000 | 0.003 | 0.002 | 0.007 | 0.038 | 3.633 | 0.000 | 0.400 |
| SD | 0.059 | 0.046 | 0.007 | 0.001 | 0.085 | 0.072 | 0.004 | 0.000 | 0.001 | 0.001 | 0.002 | 0.013 | 0.260 | 0.000 | 0.007 |
| III | 1.409 | 0.428 | 0.152 | 0.005 | 2.038 | 0.412 | 0.021 | 0.001 | 0.005 | 0.003 | 0.006 | 0.024 | 2.750 | 0.001 | 0.300 |
| SD | 0.087 | 0.033 | 0.013 | 0.003 | 0.100 | 0.041 | 0.002 | 0.000 | 0.003 | 0.001 | 0.001 | 0.002 | 0.147 | 0.000 | 0.018 |
| IV | 1.829 | 0.421 | 0.150 | 0.008 | 2.128 | 0.442 | 0.020 | 0.002 | 0.004 | 0.001 | 0.006 | 0.034 | 3.150 | 0.001 | 0.350 |
| SD | 0.067 | 0.013 | 0.011 | 0.004 | 0.102 | 0.051 | 0.001 | 0.001 | 0.001 | 0.001 | 0.002 | 0.002 | 0.247 | 0.001 | 0.022 |
| Identity | tR, min | Molecular Formula | Molecular Mass | Observed m/z [M + H]+, Da | Observed m/z [M − H]−, Da |
|---|---|---|---|---|---|
| Neochlorogenic acid L,F | 7.7 | C16H18O9 | 354.31 | 355.12 | 352.97 |
| Chlorogenic acid L,F | 8.2 | C16H18O9 | 354.31 | 355.12 | 352.96 |
| 4-O-Caffeoylquinic acid L,F | 8.3 | C16H18O9 | 354.10 | 355.10 | 353.02 |
| Caffeoylshikimic acid R | 8.8 | C16H16O8 | 336.29 | 335.00 | |
| Cinnamic acid R | 8.9 | C9H8O2 | 148.16 | 149.02 | 147.31 |
| Feruloylquinic acid L | 9.7 | C17H20O6 | 368.30 | 366.99 | |
| Rutin L,F | 10.0 | C27H30O16 | 610.52 | 611.16 | 608.96 |
| Quercetin 4′-(E-6-O-caffeoyl)glucoside L,F | 10.6 | C30H26O15 | 626.50 | 627.16 | 625.39 |
| 3,5-Dicaffeoylquinic acid L,F | 10.7 | C25H24O12 | 516.45 | 517.13 | 514.96 |
| Narcissin L | 10.9 | C28H32O16 | 624.50 | 622.99 | |
| Quercetin-3-O-(6′′-acetyl glucoside) L,F | 11.5 | C23H22O13 | 506.4 | 507.11 | 504.94 |
| 6′′-O-Acetylglycitin? L | 11.7 | C24H24O11 | 488.40 | 488.95 | |
| Ferulic acid R | 12.5 | C10H10O4 | 194.18 | 193.97 | |
| Erythrodiol-3-acetate R | 13.0 | C32H52O3 | 484.8 | 485.05 | |
| Isoquercetin (quercetin-3-O-glucoside) L,F | 14.1 | C21H20O12 | 464.096 | 468.98 | 466.93 |
| Caffeoylshikimic acid glucoside R | 14.5 | 499.12 | 500.93 | 498.51 | |
| Hyperoside (quercetin-3-O-galactoside) L,F | 15.6 | C21H20O12 | 464.38 | 465.07 | 463.25 |
| Cyanidin 3-O-glucosyl rutinoside F,R | 22.6 | C33H41O20+ | 757.77 | 758.57 | 756.01 |
| Quercetin caffeoyl hexoside F,R | 24.8 | C35H34O19 | 758.2 | 759.05 | 757.13 |
| Saponin 1 R | 27.9 | 894.76 | |||
| Saponin 2 R | 29.9 | 1112.88 | |||
| Cinnamtannin A2 R | 35.4 | C60H50O24 | 1155.02 | 1156.91 |
| Compound (RI Exp) | Flowers | Leaves | Roots | ||||||
|---|---|---|---|---|---|---|---|---|---|
| pH = 3.0 | pH = 5.9 | pH = 11.0 | pH = 3.1 | pH = 5.4 | pH = 11.1 | pH = 3.2 | pH = 6.3 | pH = 11.2 | |
| α-Pinene * (935) | 0.8 ± 0.11 | 0.5 ± 0.11 | 1.5 ± 0.47 | 0.3 ± 0.22 | 2.5 ± 0.72 | 0.3 ± 0.21 | 9.7 ± 1.22 | 9.5 ± 2.04 | 10.2 ± 1.41 |
| β-Pinene * (978) | 0.6 ± 0.28 | 1.4 ± 0.20 | 1.5 ± 0.10 | 0.9 ± 0.61 | 1.7 ± 0.33 | 0.5 ± 0.10 | 5.5 ± 0.45 | 3.7 ± 0.22 | 2.0 ± 0.24 |
| p-Cymene (1018) | 0.6 ± 0.10 | 0.7 ± 0.33 | 0.9 ± 0.09 | 4.4 ± 0.21 | 1.6 ± 0.18 | 0.3 ± 0.01 | 2.3 ± 1.72 | 2.0 ± 0.37 | 1.2 ± 0.12 |
| Limonene (1027) | 0.5 ± 0.25 | 3.2 ± 0.44 | 0.2 ± 0.14 | 5.2 ± 1.02 | 3.5 ± 0.72 | 1.3 ± 0.33 | |||
| 1,8-Cineole * (1033) | 0.2 ± 0.01 | 0.5 ± 0.15 | 0.7 ± 0.12 | 1.1 ± 0.01 | 1.1 ± 3.23 | 0.3 ± 0.12 | 11.2 ± 1.33 | 13.4 ± 0.78 | 16.1 ± 1.71 |
| trans-Pinocarveol (1135) | 0.4 ± 0.17 | 0.3 ± 0.15 | 0.4 ± 0.12 | 5.0 ± 0.50 | 5.5 ± 0.95 | 6.0 ± 0.70 | 4.1 ± 1.60 | 4.2 ± 0.91 | 4.6 ± 1.46 |
| trans-Verbenol (1143) | 0.2 ± 0.09 | 0.9 ± 0.24 | 1.3 ± 0.54 | 9.1 ± 1.18 | 20.0 ± 1.55 | 24.8 ± 1.69 | |||
| p-Mentha-1,5-dien-8-ol (1164) | 5.9 ± 1.45 | 0.5 ± 0.15 | 0.6 ± 0.05 | ||||||
| Borneol (1165) | 6.4 ± 0.72 | 4.5 ± 1.51 | 4.5 ± 1.78 | 1.2 ± 1.02 | 2.2 ± 0.51 | 2.2 ± 1.53 | |||
| Terpinen-4-ol (1174) | 0.4 ± 0.11 | 0.3 ± 0.01 | 0.1 ± 0.01 | 1.0 ± 0.50 | 0.4 ± 0.22 | 0.6 ± 0.18 | 6.5 ± 0.75 | 7.2 ± 1.51 | 8.8 ± 1.81 |
| p-Cymen-8-ol (1183) | 0.7 ± 0.27 | 3.4 ± 0.41 | 0.5 ± 0.27 | 4.2 ± 0.95 | 4.1 ± 1.58 | 4.0 ± 0.83 | |||
| α-Terpineol 1189 | 0.8 ± 0.61 | 0.9 ± 0.33 | 1.1 ± 0.21 | 0.4 ± 0.02 | 0.5 ± 0.03 | 0.3 ± 0.01 | 17.6 ± 1.63 | 19.2 ± 2.30 | 23.3 ± 2.41 |
| Verbenone (1205) | 0.4 ± 0.09 | 0.3 ± 0.04 | 0.4 ± 0.11 | 7.6 ± 1.32 | 8.7 ± 0.67 | 9.4 ± 1.13 | |||
| trans-Carveol (1219) | 3.0 ± 1.37 | 3.4 ± 0.05 | 4.2 ± 0.73 | ||||||
| 4-vinyl-Guaiacol (1310) | 11.5 ± 0.19 | 8.9 ± 0.86 | 3.0 ± 0.95 | 0.7 ± 0.09 | 1.5 ± 0.76 | 0.7 ± 0.04 | |||
| 1,2-Limonene-diol (1321) | 2.1 ± 0.33 | 2.1 ± 0.27 | 1.9 ± 1.11 | 3.9 ± 0.67 | 4.0 ± 0.18 | 6.5 ± 1.07 | |||
| Eugenol (1359) | 16.1 ± 1.53 | 11.9 ± 1.94 | 12.2 ± 0.33 | 2.2 ± 0.27 | 1.7 ± 0.63 | 1.8 ± 0.98 | |||
| trans-Myrtanol acetate (1381) | 2.0 ± 1.04 | 4.7 ± 1.78 | 5.4 ± 1.43 | ||||||
| 7,8-Dihydro-3-oxo-α-ionol (1704) | 8.7 ± 1.64 | 7.1 ± 1.79 | 7.8 ± 1.33 |
| Compound a | b RI Lit | c RI Exp | Flowers | Leaves | Roots |
|---|---|---|---|---|---|
| α-Pinene * | 939 | 938 | 21.6 ± 3.25 | 1.2 ± 0.15 | 2.6 ± 0.75 |
| β-Pinene * | 980 | 984 | 3.2 ± 0.25 | 0.2 ± 0.13 | 26.2 ± 2.23 |
| β-Myrcene | 991 | 990 | 3.0 ± 0.55 | 0.1 ± 0.01 | 4.1 ± 0.92 |
| Limonene | 1029 | 1030 | 10.2 ± 1.55 | 0.6 ± 0.10 | 35.0 ± 2.60 |
| trans-Pinocarveol | 1139 | 1136 | 1.1 ± 0.35 | 4.4 ± 1.50 | 0.2 ± 0.11 |
| cis-Verbenol | 1140 | 1145 | 0.4 ± 0.23 | 2.7 ± 0.20 | 0.1 ± 0.01 |
| trans-Verbenol | 1144 | 1146 | 4.5 ± 1.61 | 21.3 ± 1.04 | 0.1 ± 0.04 |
| o-Mentha-1,5-dien-8-ol | 1164 | 1165 | 2.2 ± 1.04 | 3.0 ± 0.41 | 0.1 ± 0.01 |
| p-Mentha-1,5-dien-8-ol | 1166 | 1170 | 0.1 ± 0.01 | 4.3 ± 1.55 | 0.1 ± 0.01 |
| Borneol | 1165 | 1168 | 0.1 ± 0.01 | 4.4 ± 1.01 | 0.1 ± 0.02 |
| Verbenone | 1204 | 1206 | 1.7 ± 0.62 | 12.5 ± 1.53 | 0.7 ± 0.35 |
| trans-Carveol | 1217 | 1215 | 1.6 ± 0.84 | 4.5 ± 1.10 | 0.2 ± 0.15 |
| Bornyl acetate | 1285 | 1290 | 6.6 ± 0.83 | 6.0 ± 0.70 | 0.1 ± 0.01 |
| β-Elemene | 1391 | 1393 | 2.9 ± 0.44 | 0.5 ± 0.24 | 3.6 ± 1.01 |
| Germacrene D | 1480 | 1485 | 15.1 ± 5.35 | 2.0 ± 1.55 | 2.0 ± 0.25 |
| Viridiflorol | 1590 | 1594 | 0.5 ± 0.21 | 3.0 ± 0.75 | 0.1 ± 0.02 |
| Germacra-4(15),5,10(14)-trien-1-α-ol | 1686 | 1685 | 0.5 ± 0.14 | 4.3 ± 1.10 | 0.2 ± 0.15 |
| Curlone (Turmerone) | 1701? | 1698 | 3.1 ± 1.35 | ||
| Lupenyl acetate | 2145? | 9.8 ± 2.35 | |||
| Average Total | 96.2 ± 1.52 | 94.1 ± 2.44 | 89.7 ± 0.95 |
| Index | Equation |
|---|---|
| Germination rate (GR), % | GR = final number of germinated seeds after 5 days of incubation/20 × 100% |
| Relative germination (RG), % | RG = GRtr (%)/GRcn (%) × 100 GRtr—mean seed germination for each treatment GRcn—mean seed germination for control |
| Vigor index (VI) | VI = PL (mm) × GR (%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Judžentienė, A.; Būdienė, J.; Labanauskas, L.; Stancelytė, D.; Nedveckytė, I. Allelopathic Activity of Canadian Goldenrod (Solidago canadensis L.) Extracts on Seed Germination and Growth of Lettuce (Lactuca sativa L.) and Garden Pepper Cress (Lepidium sativum L.). Plants 2023, 12, 1421. https://doi.org/10.3390/plants12071421
Judžentienė A, Būdienė J, Labanauskas L, Stancelytė D, Nedveckytė I. Allelopathic Activity of Canadian Goldenrod (Solidago canadensis L.) Extracts on Seed Germination and Growth of Lettuce (Lactuca sativa L.) and Garden Pepper Cress (Lepidium sativum L.). Plants. 2023; 12(7):1421. https://doi.org/10.3390/plants12071421
Chicago/Turabian StyleJudžentienė, Asta, Jurga Būdienė, Linas Labanauskas, Donata Stancelytė, and Irena Nedveckytė. 2023. "Allelopathic Activity of Canadian Goldenrod (Solidago canadensis L.) Extracts on Seed Germination and Growth of Lettuce (Lactuca sativa L.) and Garden Pepper Cress (Lepidium sativum L.)" Plants 12, no. 7: 1421. https://doi.org/10.3390/plants12071421
APA StyleJudžentienė, A., Būdienė, J., Labanauskas, L., Stancelytė, D., & Nedveckytė, I. (2023). Allelopathic Activity of Canadian Goldenrod (Solidago canadensis L.) Extracts on Seed Germination and Growth of Lettuce (Lactuca sativa L.) and Garden Pepper Cress (Lepidium sativum L.). Plants, 12(7), 1421. https://doi.org/10.3390/plants12071421