
Metabolomics Analysis Reveals Metabolites and Metabolic Pathways Involved in the Growth and Quality of Water Dropwort [Oenanthe javanica (Blume) DC.] under Nutrient Solution Culture


Abstract
:1. Introduction
2. Results
2.1. The Effects of Nutrient Solution Culture with Different Formulations on the Growth of Water Dropwort
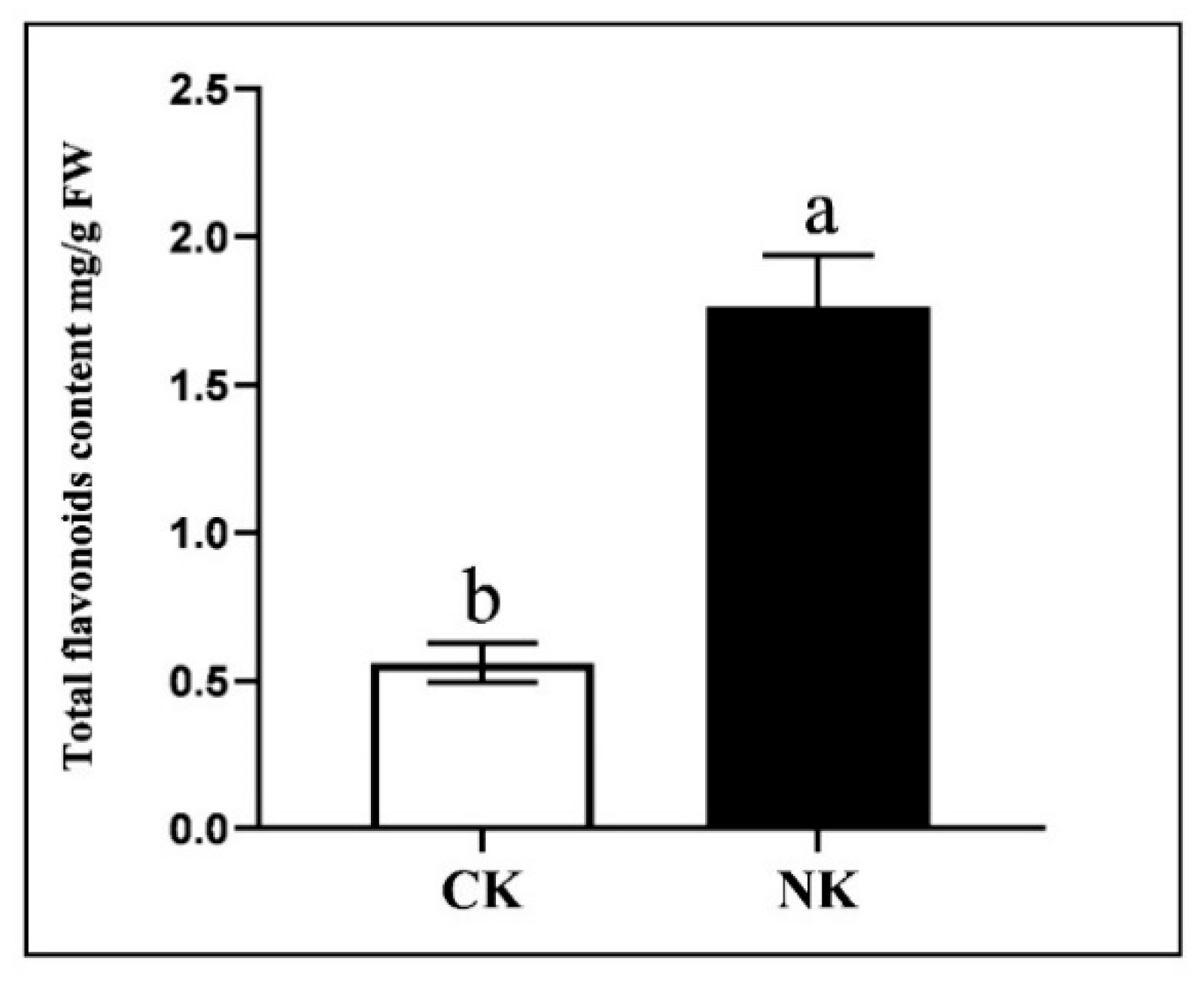
2.2. The Effects of Nutrient Solution Culture with Different Formulations on the Quality Characteristics of Water Dropwort
2.3. Comprehensive Evaluation of Growth and Quality of Spring Nutrient Solution Potted Water Dropwort Based on the Analytic Hierarchy Process (AHP)
2.3.1. Establishment of Hierarchical Structure
2.3.2. Construction and Consistency Test of Judgment Matrix
2.3.3. The Final Weight Coefficient of the Comprehensive Evaluation Index
2.3.4. Comprehensive Evaluation of Yield and Quality of Oenanthe javanica Cultivated using Different Nutrient Solution Formulas Based on Analytic Hierarchy Process
2.4. The Profiling of Metabolites
2.5. Validation of the OPLS-DA Model
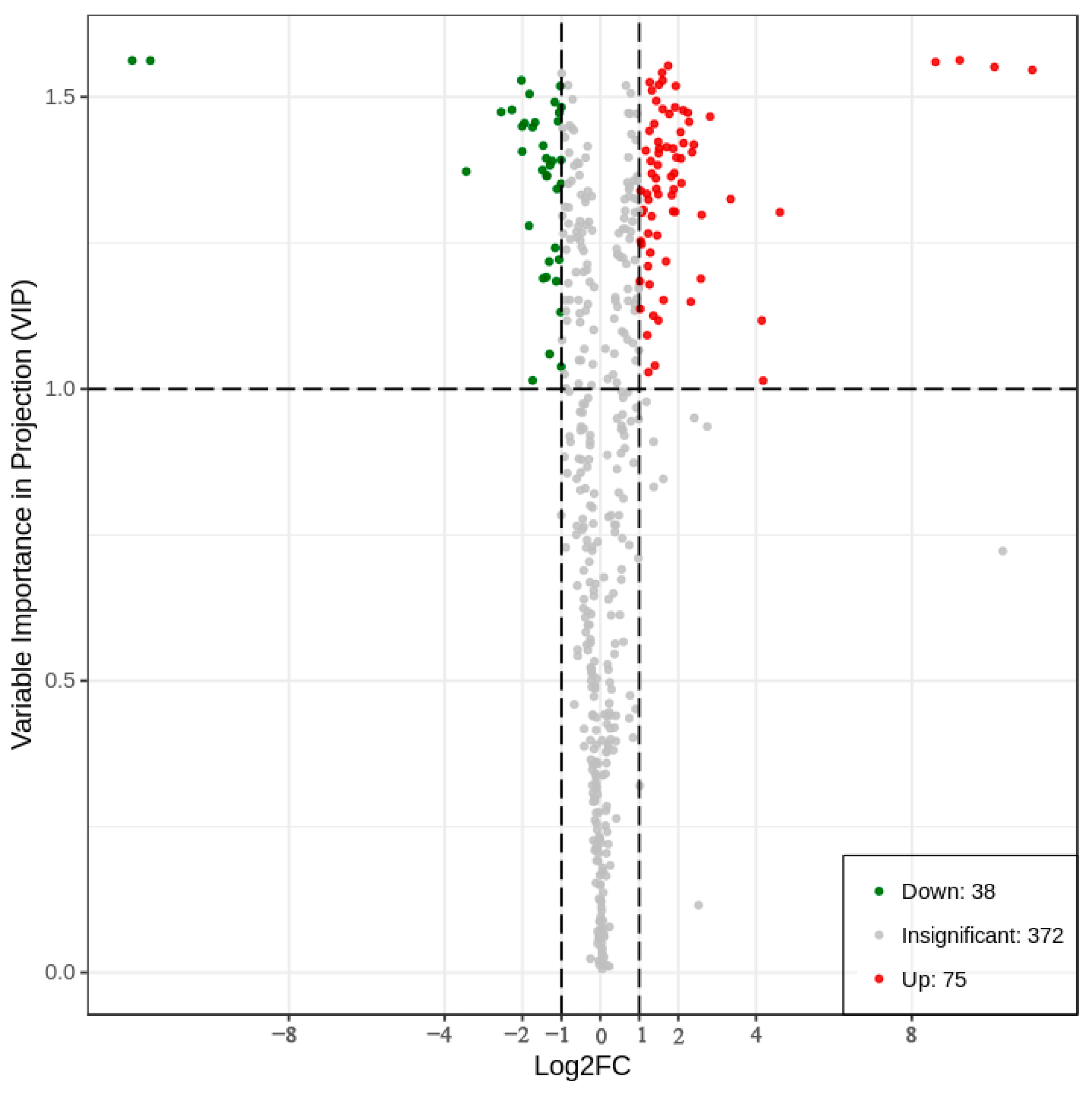
2.6. Differential Metabolite Screen
2.7. Analysis of Differential Metabolites
2.8. KEGG Annotation and Enrichment Analysis of Differential Metabolites
2.9. The Expression Levels of the Gene Involved in the Flavonoid Metabolism Pathway
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Hydroponic System
4.3. The Nutrient Solution Culture of Water Dropwort
4.4. Determination of Biomass Indexes
4.5. Determination of Physiological Indexes
4.5.1. Measurement of Chlorophyll
4.5.2. Measurement of Vitamin C (VC)
4.5.3. Measurement of Flavonoids, Total Phenolic, DPPH
4.5.4. Measurement of Dietary Fiber
4.6. Analysis Methods and Steps of the Analytic Hierarchy Process (AHP)
4.7. Metabolomics Analysis
4.7.1. Sample Extraction
4.7.2. UPLC Conditions
4.7.3. ESI−Q TRAP-MS/MS
4.7.4. Qualitative and Quantitative Metabolites
4.7.5. Sample Quality Control Analysis
4.7.6. Differential Metabolite Selected
4.7.7. KEGG Functional Annotation and Enrichment Analysis of Differential Metabolites
4.7.8. Statistical Analysis
4.8. Quantitative Real-Time PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, J.X.; Jiang, Q.; Tao, J.P.; Feng, K.; Li, T.; Duan, A.Q.; Wang, H.; Xu, Z.S.; Liu, H.; Xiong, A.S. Integrative genome, transcriptome, microRNA, and degradome analysis of water dropwort (Oenanthe javanica) in response to water stress. Hortic. Res. 2021, 8, 262. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Cho, J.H.; Kim, I.H.; Ahn, J.H.; Lee, J.C.; Chen, B.H.; Shin, B.N.; Tae, H.J.; Yoo, K.Y.; Hong, S.; et al. Oenanthe Javanica Extract Protects Against Experimentally Induced Ischemic Neuronal Damage via its Antioxidant Effects. Chin. Med. J. 2015, 128, 2932–2937. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.; Yoon, S.; Carter, O.; Bailey, G.S.; Dashwood, R.H. Antioxidant and antigenotoxic activities of Angelica keiskei, Oenanthe javanica and Brassica oleracea in the Salmonella mutagenicity assay and in HCT116 human colon cancer cells. BioFactors 2006, 26, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.A.; Jung, Y.S.; Lee, S.J.; Park, S.C.; Kim, M.J.; Lee, E.J.; Byun, H.J.; Jhee, K.H.; Lee, S.P. Hepatoprotective effects of fermented field water-dropwort (Oenanthe javanica) extract and its major constituents. Food Chem. Toxicol. 2014, 67, 154–160. [Google Scholar] [CrossRef]
- Ma, C.J.; Lee, K.Y.; Jeong, E.J.; Kim, S.H.; Park, J.; Choi, Y.H.; Kim, Y.C.; Sung, S.H. Persicarin from water dropwort (Oenanthe javanica) protects primary cultured rat cortical cells from glutamate-induced neurotoxicity. Phytother. Res. 2010, 24, 913–918. [Google Scholar] [CrossRef]
- Wang, W.N.; Yang, X.B.; Liu, H.Z.; Huang, Z.M.; Wu, G.X. Effect of Oenanthe javanica flavone on human and duck hepatitis B virus infection. Acta Pharmacol. Sin. 2005, 26, 587–592. [Google Scholar] [CrossRef]
- Feng, K.; Kan, X.Y.; Yan, Y.J.; Wang, Y.; Sun, N.; Yang, Z.Y.; Zhao, S.P.; Wu, P.; Li, L.J. Identification and characterization of terpene synthase OjTPS1 involved in β-caryophyllene biosynthesis in Oenanthe javanica (Blume) DC. Ind. Crops Prod. 2023, 192, 115998. [Google Scholar] [CrossRef]
- Kumar, S.; Huang, X.; Li, G.; Ji, Q.; Zhou, K.; Zhu, G.; Ke, W.; Hou, H.; Zhu, H.; Yang, J. Comparative Transcriptomic Analysis Provides Novel Insights into the Blanched Stem of Oenanthe javanica. Plants 2021, 10, 2484. [Google Scholar] [CrossRef]
- Feng, K.; Kan, X.Y.; Li, R.; Yan, Y.J.; Zhao, S.P.; Wu, P.; Li, L.J. Integrative Analysis of Long- and Short-Read Transcriptomes Identify the Regulation of Terpenoids Biosynthesis Under Shading Cultivation in Oenanthe javanica. Front. Genet. 2022, 13, 813216. [Google Scholar] [CrossRef]
- Song, J.W.; Shi, Y.X.; Xie, X.W.; Chai, A.L.; Li, B.J. First report of grey mould on water dropwort (Oenanthe javanica DC.) caused by Botrytis cinerea. Can. J. Plant Pathol. 2017, 39, 235–240. [Google Scholar] [CrossRef]
- Ji, Q.; Zhu, H.; Huang, X.; Zhou, K.; Liu, Z.; Sun, Y.; Wang, Z.; Ke, W. Uncovering phylogenetic relationships and genetic diversity of water dropwort using phenotypic traits and SNP markers. PLoS ONE 2021, 16, e0249825. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Wang, F.; Tan, H.W.; Li, M.Y.; Xu, Z.S.; Tan, G.F.; Xiong, A.S. De novo transcriptome assembly, gene annotation, marker development, and miRNA potential target genes validation under abiotic stresses in Oenanthe javanica. Mol. Genet. Genom. 2015, 290, 671–683. [Google Scholar] [CrossRef]
- Lu, C.L.; Li, X.F. A Review of Oenanthe javanica (Blume) DC. as Traditional Medicinal Plant and Its Therapeutic Potential. Evid. Based Complement. Altern. Med. 2019, 2019, 6495819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amalfitano, C.; Vacchio, L.; Silvano, S.; Cuciniello, A.; Caruso, G. Effects of cultural cycle and nutrient solution electrical conductivity on plant growth, yield and fruit quality of ‘Friariello’ pepper grown in hydroponics. Hortic. Sci. 2017, 44, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Rahman, A.; Azam, H.; Kim, H.S.; Kwon, M.J. Characterizing nutrient uptake kinetics for efficient crop production during Solanum lycopersicum var. cerasiforme Alef. growth in a closed indoor hydroponic system. PLoS ONE 2017, 12, e0177041. [Google Scholar] [CrossRef]
- Savvas, D.; Samantouros, K.; Paralemos, D.; Vlachakos, G.; Stamatakis, M.; Vasilatos, C. Yield and nutrient status in the root environment of tomatoes (lycopersicon esculentum) grown on chemically active and inactive inorganic substrates. Acta Hortic. 2004, 644, 377–383. [Google Scholar] [CrossRef]
- Barbosa, G.L.; Gadelha, F.D.; Kublik, N.; Proctor, A.; Reichelm, L.; Weissinger, E.; Wohlleb, G.M.; Halden, R.U. Comparison of Land, Water, and Energy Requirements of Lettuce Grown Using Hydroponic vs. Conventional Agricultural Methods. Int. J. Environ. Res. Public Health 2015, 12, 6879–6891. [Google Scholar] [CrossRef] [Green Version]
- Cifuentes-Torres, L.; Mendoza-Espinosa, L.; Reyes, J.G.; Daessle, L. Hydroponics with wastewater: A review of trends and opportunities. Water Environ. J. 2020, 35, 166–180. [Google Scholar] [CrossRef]
- Van Delden, S.H.; Nazarideljou, M.J.; Marcelis, L.F.M. Nutrient solutions for Arabidopsis thaliana: A study on nutrient solution composition in hydroponics systems. Plant Methods 2020, 16, 72. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Xiao, Z.L.; Ager, E.; Kong, L.Y.; Tan, L.B. Nutritional quality and health benefits of microgreens, a crop of modern agriculture. J. Future Foods 2021, 1, 58–66. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Pannico, A.; Graziani, G.; Kyriacou, M.; Gaspari, A.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Nutrient Supplementation Configures the Bioactive Profile and Production Characteristics of Three Brassica Microgreens Species Grown in Peat-Based Media. Agronomy 2021, 11, 346. [Google Scholar] [CrossRef]
- Kumar, S.; Li, G.; Yang, J.; Huang, X.; Ji, Q.; Liu, Z.; Ke, W.; Hou, H. Effect of Salt Stress on Growth, Physiological Parameters, and Ionic Concentration of Water Dropwort (Oenanthe javanica) Cultivars. Front. Plant Sci. 2021, 12, 660409. [Google Scholar] [CrossRef]
- Song, J.; Huang, H.; Hao, Y.; Song, S.; Zhang, Y.; Su, W.; Liu, H. Nutritional quality, mineral and antioxidant content in lettuce affected by interaction of light intensity and nutrient solution concentration. Sci. Rep. 2020, 10, 2796. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Wang, G. Nutrient concentration variations during Oenanthe javanica growth and decay in the ecological floating bed system. J. Environ. Sci. 2010, 22, 1710–1717. [Google Scholar] [CrossRef] [PubMed]
- Tacnet, J.M.; Dezert, J.; Batton-Hubert, M. AHP and Uncertainty Theories for Decision Making Using the ER-MCDA Methodology. In International Symposium on Analytic Hierarchy/Network Process; HAL Open Science: Sorrento, Italy, 2011. [Google Scholar]
- Peng, G.; Han, L.; Liu, Z.; Guo, Y.; Yan, J.; Jia, X. An Application of Fuzzy Analytic Hierarchy Process in Risk Evaluation Model. Front. Psychol. 2021, 12, 715003. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, H.K.; Juneja, S.; Muhammad, G.; Nauman, A.; Awad, N.A. Analysis of IoT-Related Ergonomics-Based Healthcare Issues Using Analytic Hierarchy Process Methodology. Sensors 2022, 22, 8232. [Google Scholar] [CrossRef] [PubMed]
- Bhushan, N.; Rai, K.; St, C. Strategic Decision Making: Applying the Analytic Hierarchy Process (Decision Engineering); Springer: London, UK, 2022. [Google Scholar]
- Ng, C.Y.; Chuah, K. Evaluation of Design Alternatives’ Environmental Performance Using AHP and ER Approaches. IEEE Syst. J. 2014, 8, 1182–1189. [Google Scholar] [CrossRef]
- Bino, R.J.; Hall, R.D.; Fiehn, O.; Kopka, J.; Saito, K.; Draper, J.; Nikolau, B.J.; Mendes, P.; Roessner-Tunali, U.; Beale, M.H.; et al. Potential of metabolomics as a functional genomics tool. Trends Plant Sci. 2004, 9, 418–425. [Google Scholar] [CrossRef]
- Fernie, A.R. Review: Metabolome characterisation in plant system analysis. Funct. Plant Biol. 2003, 30, 111–120. [Google Scholar] [CrossRef]
- Kopka, J.; Fernie, A.; Weckwerth, W.; Gibon, Y.; Stitt, M. Metabolite profiling in plant biology: Platforms and destinations. Genome Biol. 2004, 5, 109. [Google Scholar] [CrossRef] [Green Version]
- Oliver, D.J.; Nikolau, B.; Wurtele, E.S. Functional genomics: High-throughput mRNA, protein, and metabolite analyses. Metab. Eng. 2002, 4, 98–106. [Google Scholar] [CrossRef]
- Wu, P.; Liu, A.L.; Li, L.J. Metabolomics and transcriptome analysis of the biosynthesis mechanism of flavonoids in the seeds of Euryale ferox Salisb at different developmental stages. Mol. Genet. Genom. 2021, 296, 953–970. [Google Scholar] [CrossRef]
- Tiziani, R.; Miras-Moreno, B.; Malacrinò, A.; Vescio, R.; Lucini, L.; Mimmo, T.; Cesco, S.; Sorgonà, A. Drought, heat, and their combination impact the root exudation patterns and rhizosphere microbiome in maize roots. Environ. Exp. Bot. 2022, 203, 105071. [Google Scholar] [CrossRef]
- Pelletier, M.K.; Burbulis, I.E.; Winkel-Shirley, B. Disruption of specific flavonoid genes enhances the accumulation of flavonoid enzymes and end-products in Arabidopsis seedlings. Plant Mol. Biol. 1999, 40, 45–54. [Google Scholar] [CrossRef]
- Deluc, L.; Barrieu, F.; Marchive, C.; Lauvergeat, V.; Decendit, A.; Richard, T.; Carde, J.P.; Mérillon, J.M.; Hamdi, S. Characterization of a grapevine R2R3-MYB transcription factor that regulates the phenylpropanoid pathway. Plant Physiol. 2006, 140, 499–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, K.; Xu, Z.S.; Que, F.; Liu, J.X.; Wang, F.; Xiong, A.S. An R2R3-MYB transcription factor, OjMYB1, functions in anthocyanin biosynthesis in Oenanthe javanica. Planta 2018, 247, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Waraich, E.; Ahmad, R.; Ashraf, M.; Ullah, S.; Ahmad, M. Improving agricultural water use efficiency by nutrient management in crop plants. Acta Agric. Scand. Sect. B Soil Plant Sci. 2011, 61, 291–304. [Google Scholar] [CrossRef]
- Wang, N.; Fu, F.; Wang, H.; Wang, P.; He, S.; Shao, H.; Ni, Z.; Zhang, X. Effects of irrigation and nitrogen on chlorophyll content, dry matter and nitrogen accumulation in sugar beet (Beta vulgaris L.). Sci. Rep. 2021, 11, 16651. [Google Scholar] [CrossRef] [PubMed]
- Hameed, M.K.; Umar, W.; Razzaq, A.; Aziz, T.; Maqsood, M.A.; Wei, S.; Niu, Q.; Huang, D.; Chang, L. Differential Metabolic Responses of Lettuce Grown in Soil, Substrate and Hydroponic Cultivation Systems under NH(4)(+)/NO(3)(-) Application. Metabolites 2022, 12, 444. [Google Scholar] [CrossRef]
- Zhang, M.; Fang, Y.; Ji, Y.; Jiang, Z.; Wang, L. Effects of salt stress on ion content, antioxidant enzymes and protein profile in different tissues of Broussonetia papyrifera. South Afr. J. Bot. 2013, 85, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Rahneshan, Z.; Nasibi, F.; Moghadam, A.A. Effects of salinity stress on some growth, physiological, biochemical parameters and nutrients in two pistachio (Pistacia vera L.) rootstocks. J. Plant Interact. 2018, 13, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Kumar, J.; Singh, D.V.; Prasad, S. Plant Tolerance Mechanism Against Salt Stress: The Nutrient Management Approach. Biochem. Pharmacol. 2014, 3, e165. [Google Scholar] [CrossRef] [Green Version]
- Lepiniec, L.; Debeaujon, I.; Routaboul, J.M.; Baudry, A.; Pourcel, L.; Nesi, N.; Caboche, M. Genetics and biochemistry of seed flavonoids. Annu. Rev. Plant Biol. 2006, 57, 405–430. [Google Scholar] [CrossRef] [PubMed]
- Tonga, J.L.; Kamdem, M.H.K.; Pagna, J.I.M.; Fonkui, T.Y.; Tata, C.M.; Fotsing, M.C.D.; Nkengfack, E.A.; Mmutlane, E.M.; Ndinteh, D.T. Antibacterial activity of flavonoids and triterpenoids isolated from the stem bark and sap of Staudtia kamerunensis Warb. (Myristicaceae). Arab. J. Chem. 2022, 15, 104150. [Google Scholar] [CrossRef]
- Yang, L.L.; Zhou, S.W.; Hou, Y.W.; Ji, B.Y.; Pei, L.X.; Su, X.H.; Zhong, H.; Dong, C.M. Blue light induces biosynthesis of flavonoids in Epimedium sagittatum (Sieb.et Zucc.) Maxim. leaves, a study on a light-demanding medicinal shade herb. Ind. Crops Prod. 2022, 187, 115512. [Google Scholar] [CrossRef]
- Treutter, D. Significance of flavonoids in plant resistance and enhancement of their biosynthesis. Plant Biol. 2005, 7, 581–591. [Google Scholar] [CrossRef]
- Routaboul, J.M.; Dubos, C.; Beck, G.; Marquis, C.; Bidzinski, P.; Loudet, O.; Lepiniec, L. Metabolite profiling and quantitative genetics of natural variation for flavonoids in Arabidopsis. J. Exp. Bot. 2012, 63, 3749–3764. [Google Scholar] [CrossRef] [Green Version]
- Maloney, G.S.; DiNapoli, K.T.; Muday, G.K. The anthocyanin reduced tomato mutant demonstrates the role of flavonols in tomato lateral root and root hair development. Plant Physiol. 2014, 166, 614–631. [Google Scholar] [CrossRef] [Green Version]
- Ren, W.; Qiao, Z.; Wang, H.; Zhu, L.; Zhang, L. Flavonoids: Promising anticancer agents. Med. Res. Rev. 2003, 23, 519–534. [Google Scholar] [CrossRef]
- Yao, L.H.; Jiang, Y.M.; Shi, J.; Tomás-Barberán, F.A.; Datta, N.; Singanusong, R.; Chen, S.S. Flavonoids in food and their health benefits. Plant Foods Hum. Nutr. 2004, 59, 113–122. [Google Scholar] [CrossRef]
- Cushnie, T.P.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Falcone Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanner, L.A.; Li, G.; Ware, D.; Somssich, I.E.; Davis, K.R. The phenylalanine ammonia-lyase gene family in Arabidopsis thaliana. Plant Mol. Biol. 1995, 27, 327–338. [Google Scholar] [CrossRef]
- Austin, M.B.; Noel, J.P. The chalcone synthase superfamily of type III polyketide synthases. Nat. Prod. Rep. 2003, 20, 79–110. [Google Scholar] [CrossRef]
- Pang, Y.Z.; Shen, G.A.; Wu, W.S.; Liu, X.F.; Lin, J.; Tan, F.; Sun, X.F.; Tang, K.X. Characterization and expression of chalcone synthase gene from Ginkgo biloba. Plant Sci. 2005, 168, 1525–1531. [Google Scholar] [CrossRef]
- Sun, W.; Meng, X.; Liang, L.; Jiang, W.; Huang, Y.; He, J.; Hu, H.; Almqvist, J.; Gao, X.; Wang, L. Molecular and Biochemical Analysis of Chalcone Synthase from Freesia hybrid in flavonoid biosynthetic pathway. PLoS ONE 2015, 10, e0119054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koes, R.E.; Quattrocchio, F.; Mol, J.N.M. The flavonoid biosynthetic pathway in plants: Function and evolution. BioEssays 1994, 16, 123–132. [Google Scholar] [CrossRef]
- Tholl, D. Biosynthesis and biological functions of terpenoids in plants. Adv. Biochem. Eng. Biotechnol. 2015, 148, 63–106. [Google Scholar] [PubMed]
- Li, H.Z.; Cheng, Z.H. Hoagland nutrient solution promotes the growth of cucumber seedlings under light-emitting diode light. Acta Agric. Scand. Sect. B Soil Plant Sci. 2015, 65, 74–82. [Google Scholar] [CrossRef]
- Tang, Y.; Fang, Z.; Liu, M.; Zhao, D.; Tao, J. Color characteristics, pigment accumulation and biosynthetic analyses of leaf color variation in herbaceous peony (Paeonia lactiflora Pall.). 3 Biotech 2020, 10, 76. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Yu, S.; Xiong, L.; Luo, J. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: Application in the study of rice metabolomics. Mol. Plant 2013, 6, 1769–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wishart, D.S.; Jewison, T.; Guo, A.C.; Wilson, M.; Knox, C.; Liu, Y.; Djoumbou, Y.; Mandal, R.; Aziat, F.; Dong, E.; et al. HMDB 3.0—The Human Metabolome Database in 2013. Nucleic Acids Res. 2013, 41, D801–D807. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Wang, S.; Huang, Z.; Zhang, S.; Liao, Q.; Zhang, C.; Lin, T.; Qin, M.; Peng, M.; Yang, C.; et al. Rewiring of the Fruit Metabolome in Tomato Breeding. Cell 2018, 172, 249–261.e12. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.J.; Schultz, A.W.; Wang, J.; Johnson, C.H.; Yannone, S.M.; Patti, G.J.; Siuzdak, G. Liquid chromatography quadrupole time-of-flight mass spectrometry characterization of metabolites guided by the METLIN database. Nat. Protoc. 2013, 8, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Fraga, C.G.; Clowers, B.H.; Moore, R.J.; Zink, E.M. Signature-discovery approach for sample matching of a nerve-agent precursor using liquid chromatography-mass spectrometry, XCMS, and chemometrics. Anal. Chem. 2010, 82, 4165–4173. [Google Scholar] [CrossRef]
- Eriksson, L.; Johansson, E.; Kettaneh-Wold, N.; Trygg, J.; Wikstr, C.; Wold, S. Multi- and Megavariate Data Analysis. Part I Basic Principles and Applications. Second revised and enlarged edition. Ume Swed. MKS Umetrics AB 2006, 100–103. [Google Scholar]
- Zou, J.N.; Yu, H.; Yu, Q.; Jin, X.J.; Cao, L.; Wang, M.Y.; Wang, M.X.; Ren, C.Y.; Zhang, Y.X. Physiological and UPLC-MS/MS widely targeted metabolites mechanisms of alleviation of drought stress-induced soybean growth inhibition by melatonin. Ind. Crops Prod. 2021, 163, 113323. [Google Scholar] [CrossRef]
- Li, Q.; Song, J. Analysis of widely targeted metabolites of the euhalophyte Suaeda salsa under saline conditions provides new insights into salt tolerance and nutritional value in halophytic species. BMC Plant Biol. 2019, 19, 388. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Wang, S.; Tu, H.; Wan, J.; Chen, W.; Liu, X.; Luo, J.; Xu, J.; Zhang, H. Spatio-temporal distribution and natural variation of metabolites in citrus fruits. Food Chem. 2016, 199, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Plant Height (cm) | Root Length (cm) | Water Content (%) | Distribution Rate of Dry Matter (%) | Yield (g) |
|---|---|---|---|---|---|
| Hoagland (CK) | 29.03 ± 2.17 a | 17.83 ± 0.80 c | 92.29 ± 1.38 a | 78.98 ± 0.27 a | 304.57 ± 12.48 ab |
| Cooper | 30.57 ± 2.06 a | 29.48 ± 0.95 a | 92.62 ± 0.63 a | 77.89 ± 1.18 a | 299.72 ± 40.38 ab |
| Dutch greenhouse | 30.45 ± 1.01 a | 25.82 ± 2.76 ab | 92.28 ± 0.31 a | 79.17 ± 0.24 a | 329.61 ± 15.63 a |
| Garden-style | 27.63 ± 3.46 a | 14.05 ± 2.01 c | 92.10 ± 0.72 a | 75.06 ± 3.41 ab | 264.88 ± 39.11 bc |
| Yamasaki | 31.18 ± 2.18 a | 16.51 ± 1.81 c | 92.96 ± 0.55 a | 75.19 ± 1.31 ab | 300.12 ± 6.70 ab |
| SCAU | 26.83 ± 4.42 a | 23.42 ± 3.92 b | 92.35 ± 0.24 a | 72.82 ± 3.51 b | 242.30 ± 36.79 c |
| Treatments | Chlorophyll (SPAD) | VC (μg/g) | Flavonoid (mg/g FW) | Total Phenolic (mg/g FW) | DPPH (%) | IDF (%) | SDF (%) | TDF (%) |
|---|---|---|---|---|---|---|---|---|
| Hoagland | 45.00 ± 0.32 a | 43.35 ± 2.88 a | 1.52 ± 0.21 a | 8.88 ± 0.28 b | 68.18 ± 5.48 a | 55.22 ± 0.51 a | 6.10 ± 1.26 b | 61.32 ± 1.76 a |
| Cooper | 45.87 ± 1.24 a | 40.33 ± 7.60 a | 1.52 ± 0.12 a | 8.56 ± 0.77 b | 66.67 ± 6.80 a | 55.18 ± 1.08 a | 6.57 ± 1.86 ab | 61.75 ± 2.56 a |
| Dutch greenhouse | 45.19 ± 0.28 a | 42.16 ± 3.02 a | 1.61 ± 0.09 a | 9.61 ± 0.84 ab | 58.44 ± 8.22 a | 55.67 ± 1.27 a | 7.70 ± 1.08 ab | 63.37 ± 2.35 a |
| Garden-style | 44.91 ± 2.15 a | 46.94 ± 5.03 a | 1.66 ± 0.14 a | 9.44 ± 1.42 ab | 63.51 ± 4.63 a | 55.43 ± 0.96 a | 7.32 ± 1.71 ab | 62.75 ± 2.65 a |
| Yamasaki | 45.91 ± 0.76 a | 41.90 ± 3.25 a | 1.54 ± 0.2 1 a | 9.21 ± 1.10 b | 61.45 ± 1.19 a | 55.55 ± 1.65 a | 7.07 ± 0.45 ab | 62.62 ± 1.89 a |
| SCUA | 44.52 ± 2.22 a | 43.43 ± 2.46 a | 1.73 ± 0.26 a | 11.51 ± 1.87 a | 57.48 ± 9.20 a | 54.17 ± 3.71 a | 8.85 ± 1.10 a | 63.02 ± 4.10 a |
| Hierarchy | Judgment Matrix | Weight Value | Identical Test | ||||
|---|---|---|---|---|---|---|---|
| A-Ci | A | C1 | C2 | ||||
| C1 | 1 | 1/4 | 0.2 | CI = 0 | |||
| C2 | 4 | 1 | 0.8 | CR = 0 < 0.1 | |||
| C1-Pi | C1 | P1 | P2 | P3 | P4 | ||
| P1 | 1 | 1/2 | 1/3 | 1/5 | 0.0838 | CI = 0.0170 | |
| P2 | 2 | 1 | 1/2 | 1/4 | 0.1377 | CR = 0.0189 < 0.1 | |
| P3 | 3 | 2 | 1 | 1/3 | 0.2323 | ||
| P4 | 5 | 4 | 3 | 1 | 0.5462 | ||
| C2-Pi | C2 | P5 | P6 | P7 | P8 | ||
| P5 | 1 | 1 | 1/3 | 1/4 | 0.1052 | CI = 0.0278 | |
| P6 | 1 | 1 | 1/3 | 1/4 | 0.1052 | CR = 0.0309 < 0.1 | |
| P7 | 3 | 3 | 1 | 1/3 | 0.2618 | ||
| P8 | 4 | 4 | 3 | 1 | 0.5277 | ||
| Indicators | P1 | P2 | P3 | P4 | P5 | P6 | P7 | P8 |
|---|---|---|---|---|---|---|---|---|
| Final weight | 0.0168 | 0.0275 | 0.0465 | 0.1092 | 0.0842 | 0.0842 | 0.2094 | 0.4222 |
| Treatments | Hoagland | Cooper | Dutch Greenhouse | Garden-Style | Yamasaki | SCUA |
|---|---|---|---|---|---|---|
| Comprehensive evaluation value | 54.09 | 53.74 | 57.68 | 50.26 | 54.06 | 48.28 |
| Type | Number | Percentage (%) |
|---|---|---|
| All | 485 | 100.0 |
| Flavonoids | 104 | 21.4 |
| Phenolic acids | 76 | 15.7 |
| Amino acids and derivatives | 61 | 12.6 |
| Lipids | 54 | 11.1 |
| Organic acids | 40 | 8.2 |
| Nucleotides and derivatives | 40 | 8.2 |
| Lignans and Coumarins | 30 | 6.2 |
| Alkaloids | 24 | 4.9 |
| Terpenoids | 2 | 0.4 |
| Tannin | 1 | 0.2 |
| Others | 53 | 10.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, K.; Kan, X.-Y.; Liu, Q.; Yan, Y.-J.; Sun, N.; Yang, Z.-Y.; Zhao, S.-P.; Wu, P.; Li, L.-J. Metabolomics Analysis Reveals Metabolites and Metabolic Pathways Involved in the Growth and Quality of Water Dropwort [Oenanthe javanica (Blume) DC.] under Nutrient Solution Culture. Plants 2023, 12, 1459. https://doi.org/10.3390/plants12071459
Feng K, Kan X-Y, Liu Q, Yan Y-J, Sun N, Yang Z-Y, Zhao S-P, Wu P, Li L-J. Metabolomics Analysis Reveals Metabolites and Metabolic Pathways Involved in the Growth and Quality of Water Dropwort [Oenanthe javanica (Blume) DC.] under Nutrient Solution Culture. Plants. 2023; 12(7):1459. https://doi.org/10.3390/plants12071459
Chicago/Turabian StyleFeng, Kai, Xia-Yue Kan, Qing Liu, Ya-Jie Yan, Nan Sun, Zhi-Yuan Yang, Shu-Ping Zhao, Peng Wu, and Liang-Jun Li. 2023. "Metabolomics Analysis Reveals Metabolites and Metabolic Pathways Involved in the Growth and Quality of Water Dropwort [Oenanthe javanica (Blume) DC.] under Nutrient Solution Culture" Plants 12, no. 7: 1459. https://doi.org/10.3390/plants12071459
APA StyleFeng, K., Kan, X. -Y., Liu, Q., Yan, Y. -J., Sun, N., Yang, Z. -Y., Zhao, S. -P., Wu, P., & Li, L. -J. (2023). Metabolomics Analysis Reveals Metabolites and Metabolic Pathways Involved in the Growth and Quality of Water Dropwort [Oenanthe javanica (Blume) DC.] under Nutrient Solution Culture. Plants, 12(7), 1459. https://doi.org/10.3390/plants12071459