

Strategies and Methods for Improving the Efficiency of CRISPR/Cas9 Gene Editing in Plant Molecular Breeding
 , , , ,
, , , ,
Abstract
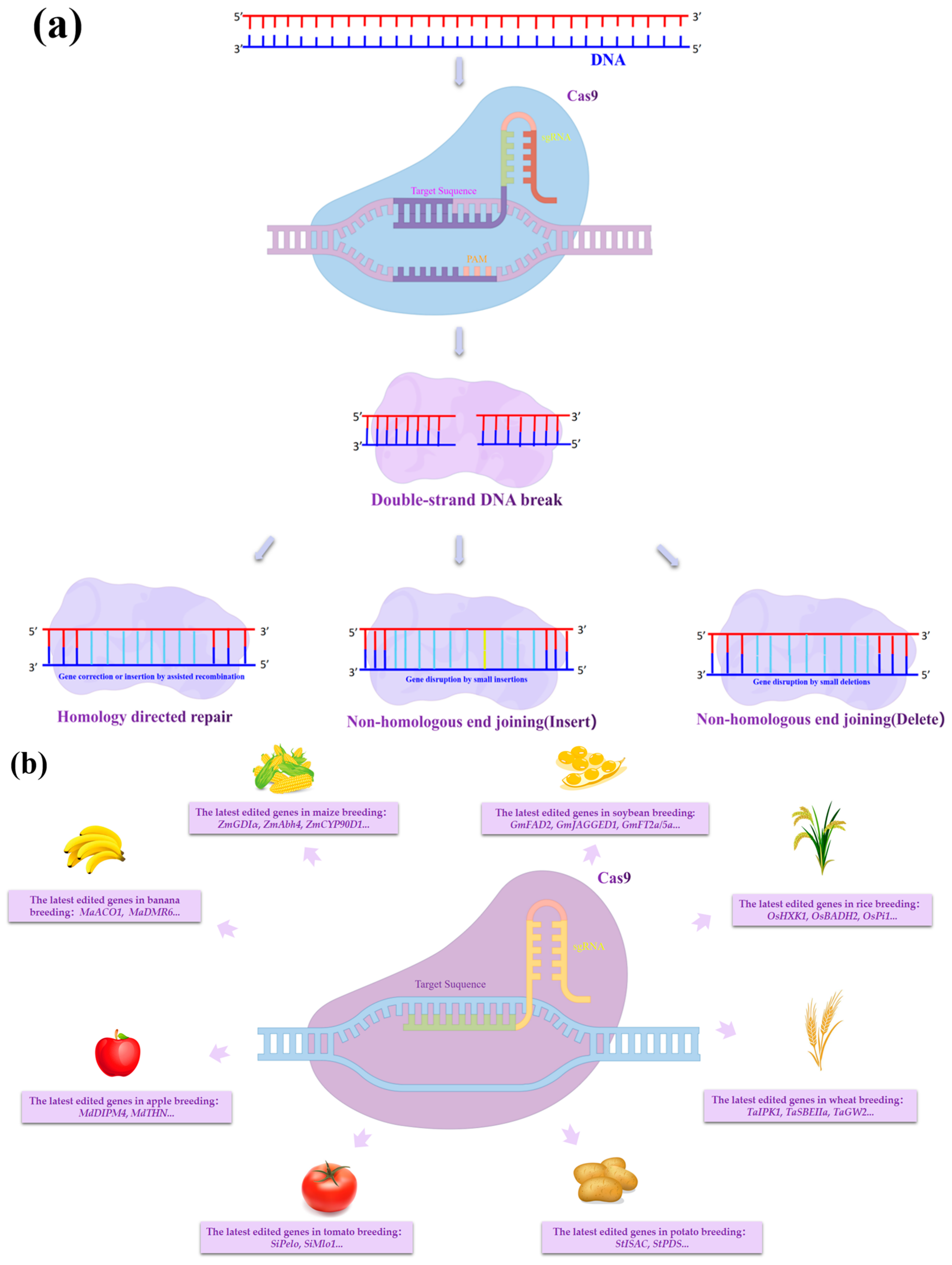
:1. Introduction
2. Advances in CRISPR/Cas9 Gene Editing Technology in the Field of Plant Molecular Breeding
3. Strategies and Methods for Optimizing CRISPR/Cas9 Gene-Editing Technology
3.1. Development of Cas9 Variant Enzymes to Expand Recognition of PAM and Improve Editing Efficiency

{kind=link}
| Cas9 Nuclease | Origin | Identifying PAM | Cutting Activation | Improving the Efficiency of Gene Editing | References |
|---|---|---|---|---|---|
| SpCas9 | S. pyogenes | NGGN | 49% | 80% | [108] |
| SaCas9 | S. aureus | NNNRRT, NNGRRT | 50% | 60.6% | [110] |
| ScCas9 | S.canis | NTG-, NGG-, NCG- | 53.6% | 57.2% | [111] |
| xCas9 | S. pyogenes | NG, GAA, GAT | 32% | 21.1% | [112] |
| Cas9-NG | S. pyogenes | NG | 30% | 56.8% | [108] |
| eSpCas9 | S. pyogenes | NGG | 40% | 80% | [113] |
| evoCas9 | S. pyogenes | NGG | 15% | - | [114] |
| SpCas9-HF2 | S. pyogenes | NGG | 34% | 65% | [115] |
| Sniper Cas9 | S. pyogenes | NGG | 46% | - | [116] |
| HypaCas9 | S. pyogenes | NGG | 30% | - | [116] |
3.2. Efficient Expression of Multiple sgRNAs Improves the Efficiency of Gene Editing
| Species | Name of Gene | Transformation Method | Gene Function | Number of sgRNA | Gene Editing Efficiency | Acquisition of New Traits | References |
|---|---|---|---|---|---|---|---|
| Soybean | GmFAD2 | Agrobacterium-mediated method | Regulation of oil synthesis | 2 | 53.3% | Creating high oleic acid soybeans | [142] |
| GmIPK1 | Agrobacterium-mediated method | Regulation of phytic acid content | 1 | 84.3% | Creating low phytic acid soybean seeds | [143] | |
| GmPDH1 | Agrobacterium-mediated method | Regulating pod breakage | 3 | 43.4% | Creating pods of unbreakable soybeans | [144] | |
| GmPDS | Agrobacterium-mediated method | Modulating the albino and dwarf phenotypes | 2 | 87.5% | Soybean for the creation of dwarf and albino phenotypes | [145] | |
| GmRS2,GmRS3 | Agrobacterium-mediated method | Regulation of oligosaccharide content in soybeans | 2 | 50.5% | Creating low oligosaccharide soybeans | [146] | |
| Rice | OsPUT | Agrobacterium-mediated method | Regulation of paraquat resistance in rice | 3 | 75% | Creating glufosinate tolerant rice | [147] |
| OsWx | Agrobacterium-mediated method | Regulation of starch content in rice | 5 | 82.5% | Creating low starch content rice | [148] | |
| OsDjA2, OsERF104 | Electroporation | Regulation of rice resistance to plague and blight | 1 | 66.65% | Creation of plague-resistant rice | [149] | |
| OsGlu | Agrobacterium-mediated method | Regulation of protein content in rice | 4 | 79.2% | Creation of high protein rice | [150] | |
| OsSAP | Agrobacterium-mediated method | Regulating drought tolerance in rice | 1 | 43.2% | Creating drought-resistant rice | [151] | |
| Maize | ZmGDIα | Agrobacterium-mediated method | Regulation of coarse and short traits in maize | 1 | 40.98% | Creating coarse dwarf disease resistant maize | [152] |
| ZmAbh4 | Agrobacterium-mediated method | Regulation of maize water use efficiency | 1 | 26.7% | Creation of high moisture utilisation maize | [153] | |
| ZmTGA9 | Agrobacterium-mediated method | Regulation of male sterility traits | 2 | 80% | Creation of male sterile maize | [68] | |
| ZmFER1 | Agrobacterium-mediated method | Regulation of resistance to Fusarium spike rot | 1 | 60% | Creation of Fusarium spike rot resistant maize | [154] | |
| ZmMYB69 | Agrobacterium-mediated method | Regulation of lignin synthesis in maize | 2 | 40% | Creation of lignin synthesis inhibiting maize | [155] |
3.3. Exploring Efficient Promoter-Activated Expression of Cas9 to Improve Editing Efficiency
3.4. Other Strategies and Methods to Improve the Efficiency of CRISPR/Cas Family Editing
4. Discussion and Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jansen, R.; Embden, J.D.A.V.; Gaastra, W.; Schouls, L.M. Identification of genes that are associated with DNA repeats in prokaryotes. Mol. Microbiol. 2002, 43, 1565–1575. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.L.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.B.; Jiang, W.Y.; Marraffini, L.A.; et al. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [Green Version]
- Esvelt, K.M.; Wang, H.H. Genome-scale engineering for systems and synthetic biology. Mol. Syst. Biol. 2013, 9, 641. [Google Scholar] [CrossRef]
- Shan, Q.W.; Wang, Y.P.; Chen, K.L.; Liang, Z.; Li, J.; Zhang, Y.; Zhang, K.; Liu, J.X.; Voytas, D.F.; Zheng, X.L.; et al. Rapid and Efficient Gene Modification in Rice and Brachypodium Using TALENs. Mol. Plant. 2013, 6, 1365–1368. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.H.; Guell, M.; Byrne, S.; Yang, J.L.; De Los Angeles, A.; Mali, P.; Aach, J.; Kim-Kiselak, C.; Briggs, A.W.; Rios, X.; et al. Optimization of scarless human stem cell genome editing. Nucleic Acids Res. 2013, 41, 9049–9061. [Google Scholar] [CrossRef]
- Tzur, Y.B.; Friedland, A.E.; Nadarajan, S.; Church, G.M.; Calarco, J.A.; Colaiacovo, M.P. Heritable Custom Genomic Modifications in Caenorhabditis elegans via a CRISPR-Cas9 System. Genetics 2013, 195, 1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.F.; Foden, J.A.; Khayter, C.; Maeder, M.L.; Reyon, D.; Joung, J.K.; Sander, J.D. High-frequency off-target mutagenesis induced by CRISPR-Cas nucleases in human cells. Nat. Biotechnol. 2013, 31, 822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, Y.C.; Yang, X.; Pan, C.Y.; Wang, C.; Wang, K.J. Advance of Clustered Regularly Interspaced Short Palindromic Repeats-Cas9 System and Its Application in Crop Improvement. Front. Plant Sci. 2022, 13, 839001. [Google Scholar] [CrossRef]
- Mao, Y.F.; Zhang, H.; Xu, N.F.; Zhang, B.T.; Gou, F.; Zhu, J.K. Application of the CRISPRCas System for Efficient Genome Engineering in Plants. Mol. Plant. 2013, 6, 2008–2011. [Google Scholar] [CrossRef] [Green Version]
- Belhaj, K.; Chaparro-Garcia, A.; Kamoun, S.; Nekrasov, V. Plant genome editing made easy: Targeted mutagenesis in model and crop plants using the CRISPR/Cas system. Plant Methods 2013, 9, 39. [Google Scholar] [CrossRef] [Green Version]
- Alamillo, J.M.; Lopez, C.M.; Martinez Rivas, F.J.; Torralbo, F.; Bulut, M.; Alseekh, S. Clustered regularly interspaced short palindromic repeats/CRISPR-associated protein and hairy roots: A perfect match for gene functional analysis and crop improvement. Curr. Opin. Biotechnol. 2023, 79, 102876. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, X.; Li, K.; Yao, Q.; Zhong, D.; Deng, Q.; Lu, Y. Large-scale genome editing in plants: Approaches, applications, and future perspectives. Curr. Opin. Biotechnol. 2023, 79, 102875. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.; Chang, F.; Sun, Z.L.; Li, N.; Meng, Q.Y. New Method of Genome Editing Derived From CRISPR/Cas9. Prog. Biochem. Biophys. 2013, 40, 691–702. [Google Scholar] [CrossRef]
- Shamshirgaran, Y.; Liu, J.; Sumer, H.; Verma, P.J.; Taheri-Ghahfarokhi, A. Tools for Efficient Genome Editing; ZFN, TALEN, and CRISPR. Methods Mol. Biol. 2022, 2495, 29–46. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Raboanatahiry, N.; Zhu, B.; Li, M.T. Progress in Genome Editing Technology and Its Application in Plants. Front. Plant Sci. 2017, 8, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saifaldeen, M.; Al-Ansari, D.E.; Ramotar, D.; Aouida, M. CRISPR FokI Dead Cas9 System: Principles and Applications in Genome Engineering. Cells 2020, 9, 2518. [Google Scholar] [CrossRef]
- Cui, Y.B.; Xu, J.M.; Cheng, M.X.; Liao, X.K.; Peng, S.L. Review of CRISPR/Cas9 sgRNA Design Tools. Interdiscip. Sci. 2018, 10, 455–465. [Google Scholar] [CrossRef]
- Nowak, C.M.; Lawson, S.; Zerez, M.; Bleris, L. Guide RNA engineering for versatile Cas9 functionality. Nucleic Acids Res. 2016, 44, 9555–9564. [Google Scholar] [CrossRef] [Green Version]
- Nashimoto, M. TRUE Gene Silencing. Int. J. Mol. Sci. 2022, 23, 5387. [Google Scholar] [CrossRef] [PubMed]
- Ran, F.A.; Hsu, P.D.; Lin, C.Y.; Gootenberg, J.S.; Konermann, S.; Trevino, A.E.; Scott, D.A.; Inoue, A.; Matoba, S.; Zhang, Y.; et al. Double Nicking by RNA-Guided CRISPR Cas9 for Enhanced Genome Editing Specificity. Cell 2013, 154, 1380–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayavaradhan, R.; Pillis, D.M.; Goodman, M.; Zhang, F.; Zhang, Y.; Andreassen, P.R.; Malik, P. CRISPR-Cas9 fusion to dominant-negative 53BP1 enhances HDR and inhibits NHEJ specifically at Cas9 target sites. Nat. Commun. 2019, 10, 2866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Movahedi, A.; Wei, H.; Zhou, X.; Fountain, J.C.; Chen, Z.; Mu, Z.; Sun, W.; Zhang, J.; Li, D.; Guo, B.; et al. Precise exogenous insertion and sequence replacements in poplar by simultaneous HDR overexpression and NHEJ suppression using CRISPR-Cas9. Hortic. Res. Engl. 2022, 9, c154. [Google Scholar] [CrossRef]
- Kato-Inui, T.; Takahashi, G.; Hsu, S.; Miyaoka, Y. Clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated protein 9 with improved proof-reading enhances homology-directed repair. Nucleic Acids Res. 2018, 46, 4677–4688. [Google Scholar] [CrossRef] [Green Version]
- Malzahn, A.; Lowder, L.; Qi, Y.P. Plant genome editing with TALEN and CRISPR. Cell Biosci. 2017, 7, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, G.Y.; Jia, M.L.; Chen, K.; Kong, X.C.; Khattak, B.; Xie, C.X.; Li, A.L.; Mao, L. CRISPR/Cas9: A powerful tool for crop genome editing. Crop J. 2016, 4, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Chaudhary, R.; Deshmukh, R.; Tiwari, S. Opportunities and challenges with CRISPR-Cas mediated homologous recombination based precise editing in plants and animals. Plant Mol. Biol. 2023, 111, 1–20. [Google Scholar] [CrossRef]
- Maruyama, T.; Dougan, S.K.; Truttmann, M.C.; Bilate, A.M.; Ingram, J.R.; Ploegh, H.L. Increasing the efficiency of precise genome editing with CRISPR-Cas9 by inhibition of nonhomologous end joining. Nat. Biotechnol. 2015, 33, 260–538. [Google Scholar] [CrossRef]
- Boti, M.A.; Athanasopoulou, K.; Adamopoulos, P.G.; Sideris, D.C.; Scorilas, A. Recent Advances in Genome-Engineering Strategies. Genes 2023, 14, 129. [Google Scholar] [CrossRef]
- Lee, C.M.; Davis, T.H.; Deshmukh, H.; Bao, G. Chromatin-Dependent Loci Accessibility Affects CRISPR-Cas9 Targeting Efficiency. Mol. Ther. 2016, 24, S54. [Google Scholar] [CrossRef]
- Mekler, V.; Kuznedelov, K.; Severinov, K. Quantification of the affinities of CRISPR/Cas9 nucleases for cognate protospacer adjacent motif (PAM) sequences. J. Biol. Chem. 2020, 295, 6509–6517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, K.T.; Floe, L.; Petersen, T.S.; Huang, J.R.; Xu, F.P.; Bolund, L.; Luo, Y.L.; Lin, L. Chromatin accessibility and guide sequence secondary structure affect CRISPR-Cas9 gene editing efficiency. FEBS Lett. 2017, 591, 1892–1901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanderheijden, N.; Delputte, P.L.; Favoreel, H.W.; Vandekerckhove, J.; Van Damme, J.; van Woensel, P.A.; Nauwynck, H.J. Involvement of sialoadhesin in entry of porcine reproductive and respiratory syndrome virus into porcine alveolar macrophages. J. Virol. 2003, 77, 8207–8215. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.A.; Erdmann, S.; Mojica, F.; Garrett, R.A. Protospacer recognition motifs: Mixed identities and functional diversity. RNA Biol. 2013, 10, 891–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.J.; Hong, S.Y.; Chen, W.J.; Zuo, E.W.; Yang, H. Advances in detecting and reducing off-target effects generated by CRISPR-mediated genome editing. J. Genet. Genom. 2019, 46, 513–521. [Google Scholar] [CrossRef]
- Myers, J.W.; Chi, J.; Gong, D.; Schaner, M.E.; Brown, P.O.; Ferrell, J.E. Minimizing off-target effects by using diced siRNAs for RNA interference. J. RNAi Gene Silenc. 2006, 2, 181–194. [Google Scholar]
- Narushima, J.; Kimata, S.; Shiwa, Y.; Gondo, T.; Akimoto, S.; Soga, K.; Yoshiba, S.; Nakamura, K.; Shibata, N.; Kondo, K. Unbiased prediction of off-target sites in genome-edited rice using SITE-Seq analysis on a web-based platform. Genes Cells 2022, 27, 706–718. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, G.L.; Yang, G.; Gu, H.F.; Huang, S.S.; Yu, W.X.; Qin, G.Z.; Liu, X.Y.; Zhou, F.L.; Huang, X.X.; et al. Increasing the targeting scope and efficiency of base editing with Proxy-BE strategy. FEBS Lett. 2020, 594, 1319–1328. [Google Scholar] [CrossRef] [PubMed]
- Tang, L. GENOME EDITING Prime editing progress. Nat. Methods 2021, 18, 592. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.L.; Qi, C.Y.; Yang, L.J.; Quan, Q.; Liu, B.L.; Zhong, Z.H.; Tang, X.; Fan, T.T.; Zhou, J.P.; Zhang, Y. The Improvement of CRISPR-Cas9 System with Ubiquitin-Associated Domain Fusion for Efficient Plant Genome Editing. Front. Plant Sci. 2020, 11, 621. [Google Scholar] [CrossRef]
- Xie, K.B.; Yang, Y.N. RNA-Guided Genome Editing in Plants Using a CRISPRCas System. Mol. Plant. 2013, 6, 1975–1983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.Z.; Zhou, H.B.; Bi, H.H.; Fromm, M.; Yang, B.; Weeks, D.P. Demonstration of CRISPR/Cas9/sgRNA-mediated targeted gene modification in Arabidopsis, tobacco, sorghum and rice. Nucleic Acids Res. 2013, 41, e188. [Google Scholar] [CrossRef] [PubMed]
- Mahfouz, M.M.; Piatek, A.; Stewart, C.N. Genome engineering via TALENs and CRISPR/Cas9 systems: Challenges and perspectives. Plant Biotechnol. J. 2014, 12, 1006–1014. [Google Scholar] [CrossRef]
- Jiang, W.Z.; Yang, B.; Weeks, D.P. Efficient CRISPR/Cas9-Mediated Gene Editing in Arabidopsis thaliana and Inheritance of Modified Genes in the T2 and T3 Generations. PLoS ONE 2014, 9, e99225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, Q.W.; Wang, Y.P.; Li, J.; Zhang, Y.; Chen, K.L.; Liang, Z.; Zhang, K.; Liu, J.X.; Xi, J.J.; Qiu, J.L.; et al. Targeted genome modification of crop plants using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 686–688. [Google Scholar] [CrossRef] [PubMed]
- Nekrasov, V.; Staskawicz, B.; Weigel, D.; Jones, J.; Kamoun, S. Targeted mutagenesis in the model plant Nicotiana benthamiana using Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31, 691–693. [Google Scholar] [CrossRef] [PubMed]
- Li, J.F.; Norville, J.E.; Aach, J.; McCormack, M.; Zhang, D.D.; Bush, J.; Church, G.M.; Sheen, J. Multiplex and homologous recombination-mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9. Nat. Biotechnol. 2013, 31, 688–691. [Google Scholar] [CrossRef]
- Ma, X.L.; Zhu, Q.L.; Chen, Y.L.; Liu, Y.G. CRISPR/Cas9 Platforms for Genome Editing in Plants: Developments and Applications. Mol. Plant. 2016, 9, 961–974. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.L.; Ma, X.L.; Xie, X.R.; Liu, Y.G. CRISPR/Cas9-Based Genome Editing in Plants. Prog. Mol. Biol. Transl. Sci. 2017, 149, 133–150. [Google Scholar] [CrossRef]
- Schaeffer, S.M.; Nakata, P.A. CRISPR/Cas9-mediated genome editing and gene replacement in plants: Transitioning from lab to field. Plant Sci. 2015, 240, 130–142. [Google Scholar] [CrossRef]
- Ding, Y.D.; Li, H.; Chen, L.L.; Xie, K.B. Recent Advances in Genome Editing Using CRISPR/Cas9. Front. Plant Sci. 2016, 7, 703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karkute, S.G.; Singh, A.K.; Gupta, O.P.; Singh, P.M.; Singh, B. CRISPR/Cas9 Mediated Genome Engineering for Improvement of Horticultural Crops. Front. Plant Sci. 2017, 8, 1635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rani, R.; Yadav, P.; Barbadikar, K.M.; Baliyan, N.; Malhotra, E.V.; Singh, B.K.; Kumar, A.; Singh, D. CRISPR/Cas9: A promising way to exploit genetic variation in plants. Biotechnol. Lett. 2016, 38, 1991–2006. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Chen, W.D.; Li, Y.S.; Sun, L.; Chai, Y.H.; Chen, H.X.; Nie, H.C.; Huang, C.L. CRISPR/Cas9 Technology and Its Utility for Crop Improvement. Int. J. Mol. Sci. 2022, 23, 10442. [Google Scholar] [CrossRef] [PubMed]
- Haq, S.; Zheng, D.F.; Feng, N.J.; Jiang, X.Y.; Qiao, F.; He, J.S.; Qiu, Q.S. Progresses of CRISPR/Cas9 genome editing in forage crops. J. Plant Physiol. 2022, 279, 153860. [Google Scholar] [CrossRef] [PubMed]
- Impens, L.; Jacobs, T.B.; Nelissen, H.; Inze, D.; Pauwels, L. Mini-Review: Transgenerational CRISPR/Cas9 Gene Editing in Plants. Front. Genome Ed. 2022, 4, 825042. [Google Scholar] [CrossRef]
- Hinge, V.R.; Chavhan, R.L.; Kale, S.P.; Suprasanna, P.; Kadam, U.S. Engineering Resistance Against Viruses in Field Crops Using CRISPR-Cas9. Curr. Genom. 2021, 22, 214–231. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, T.; Noman, M.; Shahid, M.; Muhammad, S.; Ul Qamar, M.T.; Ali, M.A.; Maqsood, A.; Hafeez, R.; Ogunyemi, S.O.; Li, B. Potential Application of CRISPR/Cas9 System to Engineer Abiotic Stress Tolerance in Plants. Protein Pept. Lett. 2021, 28, 861–877. [Google Scholar] [CrossRef]
- Shan, S.C.; Soltis, P.S.; Soltis, D.E.; Yang, B. Considerations in adapting CRISPR/Cas9 in nongenetic model plant systems. Appl. Plant Sci. 2020, 8, e11314. [Google Scholar] [CrossRef]
- Manghwar, H.; Lindsey, K.; Zhang, X.L.; Jin, S.X. CRISPR/Cas System: Recent Advances and Future Prospects for Genome Editing. Trends Plant Sci. 2019, 24, 1102–1125. [Google Scholar] [CrossRef] [Green Version]
- Liang, Z.; Wu, Y.Q.; Ma, L.L.; Guo, Y.J.; Ran, Y.D. Efficient Genome Editing in Setaria italica Using CRISPR/Cas9 and Base Editors. Front. Plant Sci. 2022, 12, 3349. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Z.; Zhang, S.W.; Jiang, Y.L.; Yan, T.W.; Fang, C.W.; Hou, Q.C.; Wu, S.W.; Xie, K.; An, X.L.; Wan, X.Y. Use of CRISPR/Cas9-Based Gene Editing to Simultaneously Mutate Multiple Homologous Genes Required for Pollen Development and Male Fertility in Maize. Cells 2022, 11, 439. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.H.; Wang, W.P.; Ali, S.; Luo, X.; Xie, L.A. CRISPR/Cas9-Mediated Multiple Knockouts in Abscisic Acid Receptor Genes Reduced the Sensitivity to ABA during Soybean Seed Germination. Int. J. Mol. Sci. 2022, 23, 16173. [Google Scholar] [CrossRef]
- Jiang, Y.L.; An, X.L.; Li, Z.W.; Yan, T.W.; Zhu, T.T.; Xie, K.; Liu, S.S.; Hou, Q.C.; Zhao, L.N.; Wu, S.W.; et al. CRISPR/Cas9-based discovery of maize transcription factors regulating male sterility and their functional conservation in plants. Plant Biotechnol. J. 2021, 19, 1769–1784. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.H.; Luo, T.T.; Hua, Y.H.; Yan, X.H.; Liu, X.; Liu, Y.; Liu, Y.P.; Zhang, B.L.; Liu, R.; Zhu, Z.Z.; et al. Assessment of the Characteristics of Waxy Rice Mutants Generated by CRISPR/Cas9. Front. Plant Sci. 2022, 13, 1829. [Google Scholar] [CrossRef]
- Tianye, Z.; Haichao, H.; Ziqiong, W.; Tianyou, F.; Lu, Y.; Zhang, J.; Wenqing, G.; Yilin, Z.; Sun, M.; Liu, P.; et al. Wheat yellow mosaic virus NIb targets TaVTC2 to elicit broad-spectrum pathogen resistance in wheat. Plant Biotechnol. J. 2023, 2023, 1–16. [Google Scholar] [CrossRef]
- Yang, T.; Ali, M.; Lin, L.; Li, P.; He, H.; Zhu, Q.; Sun, C.; Wu, N.; Zhang, X.; Huang, T.; et al. Recoloring tomato fruit by CRISPR/Cas9-mediated multiplex gene editing. Hortic. Res. Engl. 2023, 10, c214. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, J.H.; Jang, Y.H.; Yu, J.; Bae, S.; Kim, M.S.; Cho, Y.G.; Jung, Y.J.; Kang, K.K. Transcriptome and Metabolite Profiling of Tomato SGR-Knockout Null Lines Using the CRISPR/Cas9 System. Int. J. Mol. Sci. 2023, 24, 109. [Google Scholar] [CrossRef]
- Osakabe, Y.; Liang, Z.C.; Ren, C.; Nishitani, C.; Osakabe, K.; Wada, M.; Komori, S.; Malnoy, M.; Velasco, R.; Poli, M.; et al. CRISPR-Cas9-mediated genome editing in apple and grapevine. Nat. Protoc. 2018, 13, 2844–2863. [Google Scholar] [CrossRef]
- Pompili, V.; Dalla Costa, L.; Piazza, S.; Pindo, M.; Malnoy, M. Reduced fire blight susceptibility in apple cultivars using a high-efficiency CRISPR/Cas9-FLP/FRT-based gene editing system. Plant Biotechnol. J. 2020, 18, 845–858. [Google Scholar] [CrossRef]
- Ma, J.; Zheng, A.H.; Zhou, P.; Yuan, Q.; Wu, R.; Chen, C.Y.; Wu, X.Z.; Zhang, F.; Sun, B. Targeted Editing of the StPDS Gene using the CRISPR/Cas9 system in Tetraploid Potato. Emir. J. Food Agric. 2019, 31, 482–490. [Google Scholar] [CrossRef]
- Noureen, A.; Khan, M.Z.; Amin, I.; Zainab, T.; Mansoor, S. CRISPR/Cas9-Mediated Targeting of Susceptibility Factor eIF4E-Enhanced Resistance Against Potato Virus Y. Front. Genet. 2022, 13, 922019. [Google Scholar] [CrossRef]
- Gao, X.H.; Chen, J.L.; Dai, X.H.; Zhang, D.; Zhao, Y.D. An Effective Strategy for Reliably Isolating Heritable and Cas9-Free Arabidopsis Mutants Generated by CRISPR/Cas9-Mediated Genome Editing. Plant Physiol. 2016, 171, 1794–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowder, L.G.; Zhang, D.W.; Baltes, N.J.; Paul, J.W.; Tang, X.; Zheng, X.L.; Voytas, D.F.; Hsieh, T.F.; Zhang, Y.; Qi, Y.P. A CRISPR/Cas9 Toolbox for Multiplexed Plant Genome Editing and Transcriptional Regulation. Plant Physiol. 2015, 169, 971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, M.; Haider, M.S.; Zhai, Y.; Khan, M.; Pappu, H.R. Towards developing resistance to chickpea chlorotic dwarf virus through CRISPR/Cas9-mediated gene editing using multiplexed gRNAs. J. Plant Dis. Prot. 2022, 130, 23–33. [Google Scholar] [CrossRef]
- Ma, X.; Liu, Y. CRISPR/Cas9-Based Multiplex Genome Editing in Monocot and Dicot Plants. Curr. Protoc. Mol. Biol. 2016, 115, 31–36. [Google Scholar] [CrossRef]
- Zhang, F.; Huang, Z.W. Mechanistic insights into the versatile class II CRISPR toolbox. Trends Biochem.Sci. 2022, 47, 433–450. [Google Scholar] [CrossRef]
- Westra, E.R.; Nilges, B.; van Erp, P.; van der Oost, J.; Dame, R.T.; Brouns, S. Cascade-mediated binding and bending of negatively supercoiled DNA. RNA Biol. 2012, 9, 1134–1138. [Google Scholar] [CrossRef] [Green Version]
- Collias, D.; Beisel, C.L. CRISPR technologies and the search for the PAM-free nuclease. Nat. Commun. 2021, 12, 555. [Google Scholar] [CrossRef]
- Murovec, J.; Pirc, Z.; Yang, B. New variants of CRISPR RNA-guided genome editing enzymes. Plant Biotechnol. J. 2017, 15, 917–926. [Google Scholar] [CrossRef] [Green Version]
- Uranga, M.; Daros, J.A. Tools and targets: The dual role of plant viruses in CRISPR-Cas genome editing. Plant Genome 2022, e20220. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Beisel, C.L. The tracrRNA in CRISPR Biology and Technologies. Annu. Rev. Genet. 2021, 55, 161–181. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.S.; Gong, S.; Yu, H.H.; Taylor, D.W.; Johnson, K.A. Kinetic characterization of Cas9 enzymes. Methods Enzymol. 2019, 616, 289–311. [Google Scholar] [CrossRef] [PubMed]
- Mikami, M.; Toki, S.; Endo, M. In Planta Processing of the SpCas9-gRNA Complex. Plant Cell Physiol. 2017, 58, 1857–1867. [Google Scholar] [CrossRef]
- Kang, M.J.; Zuo, Z.C.; Yin, Z.X.; Gu, J.R. Molecular Mechanism of D1135E-Induced Discriminated CRISPR-Cas9 PAM Recognition. J. Chem. Inf. Model. 2022, 62, 3057–3066. [Google Scholar] [CrossRef]
- Hirano, S.; Nishimasu, H.; Ishitani, R.; Nureki, O. Structural Basis for the Altered PAM Specificities of Engineered CRISPR-Cas9. Mol. Cell 2016, 61, 886–894. [Google Scholar] [CrossRef] [Green Version]
- Gurel, F.; Zhang, Y.; Sretenovic, S.; Qi, Y. CRISPR-Cas nucleases and base editors for plant genome editing. aBIOTECH 2020, 1, 74–87. [Google Scholar] [CrossRef] [Green Version]
- Kim, N.; Kim, H.K.; Lee, S.; Seo, J.H.; Choi, J.W.; Park, J.; Min, S.; Yoon, S.; Cho, S.R.; Kim, H.H. Prediction of the sequence-specific cleavage activity of Cas9 variants. Nat. Biotechnol. 2020, 38, 1328. [Google Scholar] [CrossRef]
- Zhang, D.B.; Zhang, H.W.; Li, T.D.; Chen, K.L.; Qiu, J.L.; Gao, C.X. Perfectly matched 20-nucleotide guide RNA sequences enable robust genome editing using high-fidelity SpCas9 nucleases. Genome Biol. 2017, 18, 191. [Google Scholar] [CrossRef] [Green Version]
- Ge, Z.X.; Zheng, L.Q.; Zhao, Y.L.; Jiang, J.H.; Zhang, E.J.; Liu, T.X.; Gu, H.Y.; Qu, L.J. Engineered xCas9 and SpCas9-NG variants broaden PAM recognition sites to generate mutations in Arabidopsis plants. Plant Biotechnol. J. 2019, 17, 1865–1867. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.D.; Scott, D.A.; Weinstein, J.A.; Ran, F.A.; Konermann, S.; Agarwala, V.; Li, Y.Q.; Fine, E.J.; Wu, X.B.; Shalem, O.; et al. DNA targeting specificity of RNA-guided Cas9 nucleases. Nat. Biotechnol. 2013, 31, 827. [Google Scholar] [CrossRef] [PubMed]
- Kleinstiver, B.P.; Pattanayak, V.; Prew, M.S.; Tsai, S.Q.; Nguyen, N.T.; Zheng, Z.L.; Joung, J.K. High-fidelity CRISPR-Cas9 nucleases with no detectable genome-wide off-target effects. Nature 2016, 529, 490. [Google Scholar] [CrossRef] [Green Version]
- Idoko-Akoh, A.; Taylor, L.; Sang, H.M.; McGrew, M.J. High fidelity CRISPR/Cas9 increases precise monoallelic and biallelic editing events in primordial germ cells. Sci. Rep. 2018, 8, 15126. [Google Scholar] [CrossRef] [Green Version]
- Negishi, K.; Kaya, H.; Abe, K.; Hara, N.; Saika, H.; Toki, S. An adenine base editor with expanded targeting scope using SpCas9-NGv1 in rice. Plant Biotechnol. J. 2019, 17, 1476–1478. [Google Scholar] [CrossRef]
- Zhang, Q.; Xing, H.L.; Wang, Z.P.; Zhang, H.Y.; Yang, F.; Wang, X.C.; Chen, Q.J. Potential high-frequency off-target mutagenesis induced by CRISPR/Cas9 in Arabidopsis and its prevention. Plant Mol. Biol. 2018, 96, 445–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Z.H.; Sretenovic, S.; Ren, Q.R.; Yang, L.J.; Bao, Y.; Qi, C.Y.; Yuan, M.Z.; He, Y.; Liu, S.S.; Liu, X.P.; et al. Improving Plant Genome Editing with High-Fidelity xCas9 and Non-canonical PAM-Targeting Cas9-NG. Mol. Plant 2019, 12, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, S.; Rahman, H.; Yamanaka, N.; Ishizaki, C.; Islam, S.; Aiso, T.; Hirata, S.; Yamamoto, M.; Kobayashi, K.; Kaya, H. A Simple Heat Treatment Increases SpCas9-Mediated Mutation Efficiency in Arabidopsis. Plant Cell Physiol. 2021, 62, 1676–1686. [Google Scholar] [CrossRef] [PubMed]
- Carrijo, J.; Illa-Berenguer, E.; LaFayette, P.; Torres, N.; Aragao, F.; Parrott, W.; Vianna, G.R. Two efficient CRISPR/Cas9 systems for gene editing in soybean. Transgenic Res. 2021, 30, 239–249. [Google Scholar] [CrossRef]
- Liu, T.L.; Zeng, D.C.; Zheng, Z.Y.; Lin, Z.S.; Xue, Y.; Li, T.; Xie, X.R.; Ma, G.L.; Liu, Y.G.; Zhu, Q.L. The ScCas9(++) variant expands the CRISPR toolbox for genome editing in plants. J. Integr. Plant Biol. 2021, 63, 1611–1619. [Google Scholar] [CrossRef]
- Li, X.R.; Zuo, X.; Li, M.M.; Yang, X.; Zhi, J.Y.; Sun, H.Z.; Xie, C.X.; Zhang, Z.Y.; Wang, F.Q. Efficient CRISPR/Cas9-mediated genome editing in Rehmannia glutinosa. Plant Cell Rep. 2021, 40, 1695–1707. [Google Scholar] [CrossRef]
- Jedlickova, V.; Macova, K.; Stefkova, M.; Butula, J.; Stavenikova, J.; Sedlacek, M.; Robert, H.S. Hairy root transformation system as a tool for CRISPR/Cas9-directed genome editing in oilseed rape (Brassica napus). Front. Plant Sci. 2022, 13, 2514. [Google Scholar] [CrossRef] [PubMed]
- Qin, R.Y.; Li, J.; Liu, X.S.; Xu, R.F.; Yang, J.B.; Wei, P.C. SpCas9-NG self-targets the sgRNA sequence in plant genome editing. Nat. Plants 2020, 6, 197. [Google Scholar] [CrossRef]
- Hua, K.; Tao, X.P.; Han, P.J.; Wang, R.; Zhu, J.K. Genome Engineering in Rice Using Cas9 Variants that Recognize NG PAM Sequences. Mol. Plant. 2019, 12, 1003–1014. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, M. Recent advances and applications of base editing systems. Sheng Wu Gong Cheng Xue Bao Chin. J. Biotechnol. 2021, 37, 2307–2321. [Google Scholar] [CrossRef]
- Li, Y.; Li, W.J.; Li, J. The CRISPR/Cas9 revolution continues: From base editing to prime editing in plant science. J. Genet. Genomics 2021, 48, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Sichani, A.S.; Ranjbar, M.; Baneshi, M.; Zadeh, F.T.; Fallahi, J. A Review on Advanced CRISPR-Based Genome-Editing Tools: Base Editing and Prime Editing. Mol. Biotechnol. 2022. [Google Scholar] [CrossRef]
- Wu, Y.; Xu, W.; Wang, F.P.; Zhao, S.; Feng, F.; Song, J.L.; Zhang, C.W.; Yang, J.X. Increasing Cytosine Base Editing Scope and Efficiency with Engineered Cas9-PmCDA1 Fusions and the Modified sgRNA in Rice. Front. Genet. 2019, 10, 379. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.T.; Zeng, D.C.; Zhao, Y.C.; Wang, Y.X.; Liu, T.L.; Li, S.C.; Xue, Y.; Luo, Y.Y.; Xie, X.R.; Chen, L.T.; et al. PhieABEs: A PAM-less/free high-efficiency adenine base editor toolbox with wide target scope in plants. Plant Biotechnol. J. 2022, 20, 934–943. [Google Scholar] [CrossRef]
- Qiao, D.X.; Wang, J.Y.; Lu, M.H.; Xin, C.P.; Chai, Y.P.; Jiang, Y.Y.; Sun, W.; Cao, Z.H.; Guo, S.Y.; Wang, X.C.; et al. Optimized prime editing efficiently generates heritable mutations in maize. J. Integr. Plant Biol. 2023. [Google Scholar] [CrossRef]
- Kaya, H.; Ishibashi, K.; Toki, S. A Split Staphylococcus aureus Cas9 as a Compact Genome-Editing Tool in Plants. Plant Cell Physiol. 2017, 58, 643–649. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.G.; Kuang, Y.J.; Lu, Z.W.; Li, X.Q.; Xu, Z.Y.; Ren, B.; Zhou, X.P.; Zhou, H.B. CRISPR/Sc++-mediated genome editing in rice. J. Integr. Plant Biol. 2021, 63, 1606–1610. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Meng, X.B.; Hu, X.X.; Sun, T.T.; Li, J.Y.; Wang, K.J.; Yu, H. xCas9 expands the scope of genome editing with reduced efficiency in rice. Plant Biotechnol. J. 2019, 17, 709–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Song, W.; Yang, Y.X.; Wu, Y.; Lv, X.X.; Yuan, S.; Liu, Y.; Yang, J.X. Multiplex nucleotide editing by high-fidelity Cas9 variants with improved efficiency in rice. BMC Plant Biol. 2019, 19, 511. [Google Scholar] [CrossRef] [PubMed]
- Casini, A.; Olivieri, M.; Petris, G.; Montagna, C.; Reginato, G.; Maule, G.; Lorenzin, F.; Prandi, D.; Romanel, A.; Demichelis, F.; et al. A highly specific SpCas9 variant is identified by in vivo screening in yeast. Nat. Biotechnol. 2018, 36, 265. [Google Scholar] [CrossRef] [PubMed]
- Grzechnik, P.; Szczepaniak, S.A.; Dhir, S.; Pastucha, A.; Parslow, H.; Matuszek, Z.; Mischo, H.E.; Kufel, J.; Proudfoot, N.J. Nuclear fate of yeast snoRNA is determined by co-transcriptional Rnt1 cleavage. Nat. Commun. 2018, 9, 1783. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Wang, S.; Yu, L.; Zhang, X.; Chen, W. Advances of Cas9/sgRNA delivery system for gene editing. Sheng Wu Gong Cheng Xue Bao Chin. J. Biotechnol. 2021, 37, 3880–3889. [Google Scholar] [CrossRef]
- Hu, W.X.; Rong, Y.; Guo, Y.; Jiang, F.; Tian, W.; Chen, H.; Dong, S.S.; Yang, T.L. ExsgRNA: Reduce off-target efficiency by on-target mismatched sgRNA. Brief. Bioinform. 2022, 23, bbac183. [Google Scholar] [CrossRef]
- Wu, J.; Yin, H. Engineering guide RNA to reduce the off-target effects of CRISPR. J. Genet. Genom. 2019, 46, 523–529. [Google Scholar] [CrossRef]
- Zhou, Y.; Fu, Q.; Shi, H.; Zhou, G. CRISPR Guide RNA Library Screens in Human Induced Pluripotent Stem Cells. Methods Mol. Biol. 2022, 2549, 233–257. [Google Scholar] [CrossRef]
- Moreb, E.A.; Lynch, M.D. Genome dependent Cas9/gRNA search time underlies sequence dependent gRNA activity. Nat. Commun. 2021, 12, 5034. [Google Scholar] [CrossRef]
- Li, Z.B.; Cheng, Q.; Gan, Z.R.; Hou, Z.H.; Zhang, Y.H.; Li, Y.L.; Li, H.Y.; Nan, H.Y.; Yang, C.; Chen, L.N.; et al. Multiplex CRISPR/Cas9-mediated knockout of soybean LNK2 advances flowering time. Crop J. 2021, 9, 767–776. [Google Scholar] [CrossRef]
- Wang, T.Y.; Xun, H.W.; Wang, W.; Ding, X.Y.; Tian, H.A.; Hussain, S.; Dong, Q.L.; Li, Y.Y.; Cheng, Y.X.; Wang, C.; et al. Mutation of GmAITR Genes by CRISPR/Cas9 Genome Editing Results in Enhanced Salinity Stress Tolerance in Soybean. Front. Plant Sci. 2021, 12, 2752. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Tian, L.N. An efficient and specific CRISPR-Cas9 genome editing system targeting soybean phytoene desaturase genes. BMC Biotechnol. 2022, 22, 7. [Google Scholar] [CrossRef]
- Thomson, M. Genome editing applications in plants: High-throughput CRISPR/Cas editing for crop improvement. J. Anim. Sci. 2019, 97, 56. [Google Scholar] [CrossRef]
- Ursache, R.; Fujita, S.; Tendon, V.D.; Geldner, N. Combined fluorescent seed selection and multiplex CRISPR/Cas9 assembly for fast generation of multiple Arabidopsis mutants. Plant Methods 2021, 17, 111. [Google Scholar] [CrossRef]
- Zheng, X.L.; Zhang, S.T.; Liang, Y.L.; Zhang, R.; Liu, L.; Qin, P.C.; Zhang, Z.; Wang, Y.; Zhou, J.P.; Tang, X.; et al. Loss-function mutants of OsCKX gene family based on CRISPR-Cas systems revealed their diversified roles in rice. Plant Genome 2023, e20283. [Google Scholar] [CrossRef]
- Yu, Z.M.; Chen, Q.Y.; Chen, W.W.; Zhang, X.; Mei, F.L.; Zhang, P.C.; Zhao, M.; Wang, X.H.; Shi, N.N.; Jackson, S.; et al. Multigene editing via CRISPR/Cas9 guided by a single-sgRNA seed in Arabidopsis. J. Integr. Plant Biol. 2018, 60, 376–381. [Google Scholar] [CrossRef] [Green Version]
- Bao, A.L.; Chen, H.F.; Chen, L.M.; Chen, S.L.; Hao, Q.N.; Guo, W.; Qiu, D.Z.; Shan, Z.H.; Yang, Z.L.; Yuan, S.L.; et al. CRISPR/Cas9-mediated targeted mutagenesis of GmSPL9 genes alters plant architecture in soybean. BMC Plant Biol. 2019, 19, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.X.; Liu, X.Q.; Zheng, X.X.; Wang, W.X.; Yin, X.Q.; Liu, H.F.; Ma, C.L.; Niu, X.M.; Zhu, J.K.; Wang, F. Creation of aromatic maize by CRISPR/Cas. J. Integr. Plant Biol. 2021, 63, 1664–1670. [Google Scholar] [CrossRef]
- Ren, C.; Liu, Y.F.; Guo, Y.C.; Duan, W.; Fan, P.G.; Li, S.H.; Liang, Z.C. Optimizing the CRISPR/Cas9 system for genome editing in grape by using grape promoters. Hortic. Res. Engl. 2021, 8, 52. [Google Scholar] [CrossRef]
- Cao, L.; Wang, Z.R.; Ma, H.Y.; Liu, T.F.; Ji, J.; Duan, K.X. Multiplex CRISPR/Cas9-mediated raffinose synthase gene editing reduces raffinose family oligosaccharides in soybean. Front. Plant Sci. 2022, 13, 1048967. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Wahl, N.J.; Thomson, M.J.; Cason, J.M.; McCutchen, B.F.; Septiningsih, E.M. Optimization of Protoplast Isolation and Transformation for a Pilot Study of Genome Editing in Peanut by Targeting the Allergen Gene Ara h 2. Int. J. Mol. Sci. 2022, 23, 837. [Google Scholar] [CrossRef]
- Zhang, S.; Wu, S.P.; Hu, C.H.; Yang, Q.S.; Dong, T.; Sheng, O.; Deng, G.M.; He, W.D.; Dou, T.X.; Li, C.Y.; et al. Increased mutation efficiency of CRISPR/Cas9 genome editing in banana by optimized construct. PeerJ 2022, 10, e12664. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.N.; Kim, H.J.; Chung, Y.S.; Kim, H.U. Construction of Multiple Guide RNAs in CRISPR/Cas9 Vector Using Stepwise or Simultaneous Golden Gate Cloning: Case Study for Targeting the FAD2 and FATB Multigene in Soybean. Plants 2021, 10, 2542. [Google Scholar] [CrossRef] [PubMed]
- Bai, M.Y.; Yuan, J.H.; Kuang, H.Q.; Gong, P.P.; Li, S.N.; Zhang, Z.H.; Liu, B.; Sun, J.F.; Yang, M.X.; Yang, L.; et al. Generation of a multiplex mutagenesis population via pooled CRISPR-Cas9 in soya bean. Plant Biotechnol. J. 2020, 18, 721–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattiello, L.; Rutgers, M.; Sua-Rojas, M.F.; Tavares, R.; Soares, J.S.; Begcy, K.; Menossi, M. Molecular and Computational Strategies to Increase the Efficiency of CRISPR-Based Techniques. Front. Plant Sci. 2022, 13, 2625. [Google Scholar] [CrossRef]
- Reuven, N.; Shaul, Y. Selecting for CRISPR-Edited Knock-In Cells. Int. J. Mol. Sci. 2022, 23, 11919. [Google Scholar] [CrossRef]
- Carlsen, F.M.; Johansen, I.E.; Yang, Z.; Liu, Y.; Westberg, I.N.; Kieu, N.P.; Jorgensen, B.; Lenman, M.; Andreasson, E.; Nielsen, K.L.; et al. Corrigendum: Strategies for Efficient Gene Editing in Protoplasts of Solanum tuberosum Theme: Determining gRNA Efficiency Design by Utilizing Protoplast (Research). Front. Genome Ed. 2022, 4, 914100. [Google Scholar] [CrossRef]
- Pan, W.B.; Liu, X.; Li, D.Y.; Zhang, H.W. Establishment of an Efficient Genome Editing System in Lettuce Without Sacrificing Specificity. Front. Plant Sci. 2022, 13, 1985. [Google Scholar] [CrossRef]
- Kumar, R.; Kamuda, T.; Budhathoki, R.; Tang, D.; Yer, H.; Zhao, Y.; Li, Y. Agrobacterium- and a single Cas9-sgRNA transcript system-mediated high efficiency gene editing in perennial ryegrass. Front. Genome Ed. 2022, 4, 960414. [Google Scholar] [CrossRef]
- Mu, R.L.; Lyu, X.; Ji, R.H.; Liu, J.; Zhao, T.; Li, H.Y.; Liu, B. GmBICs Modulate Low Blue Light-Induced Stem Elongation in Soybean. Front. Plant Sci. 2022, 13, 803122. [Google Scholar] [CrossRef]
- Do, P.T.; Nguyen, C.X.; Bui, H.T.; Tran, L.; Stacey, G.; Gillman, J.D.; Zhang, Z.; Stacey, M.G. Demonstration of highly efficient dual gRNA CRISPR/Cas9 editing of the homeologous GmFAD2-1A and GmFAD2-1B genes to yield a high oleic, low linoleic and alpha-linolenic acid phenotype in soybean. BMC Plant Biol. 2019, 19, 311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.H.; Shin, G.; Kim, H.J.; Lee, S.B.; Moon, J.Y.; Jeong, J.C.; Choi, H.K.; Kim, I.A.; Song, H.J.; Kim, C.Y.; et al. Mutation of GmIPK1 Gene Using CRISPR/Cas9 Reduced Phytic Acid Content in Soybean Seeds. Int. J. Mol. Sci. 2022, 23, 10583. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.F.; Xu, W.Y.; Liu, T.; Chen, R.Y.; Zhu, H.; Zhang, H.Y.; Cai, C.M.; Li, S. Functional characterization of soybean (Glycine max) DIRIGENT genes reveals an important role of GmDIR27 in the regulation of pod dehiscence. Genomics 2021, 113, 979–990. [Google Scholar] [CrossRef]
- Kim, K.H.; Lim, S.; Kang, Y.J.; Yoon, M.Y.; Nam, M.; Jun, T.H.; Seo, M.J.; Baek, S.B.; Lee, J.H.; Moon, J.K.; et al. Optimization of a Virus-Induced Gene Silencing System with Soybean yellow common mosaic virus for Gene Function Studies in Soybeans. Plant Pathol. J. 2016, 32, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.; Mota, L.M.; Fonseca, L.; Bueno, R.D.; Piovesan, N.D.; Fontes, E.; Dal-Bianco, M. Effect of a mutation in Raffinose Synthase 2 (GmRS2) on soybean quality traits. Crop Breed. Appl. Biotechnol. 2019, 19, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Lyu, Y.; Cao, L.; Huang, W.; Liu, J.; Lu, H. Disruption of three polyamine uptake transporter genes in rice by CRISPR/Cas9 gene editing confers tolerance to herbicide paraquat. aBIOTECH 2022, 3, 140–145. [Google Scholar] [CrossRef]
- Alam, M.S.; Yang, Z.K.; Li, C.; Yan, Y.; Liu, Z.; Nazir, M.M.; Xu, J.H. Loss-of-function mutations of OsbHLH044 transcription factor lead to salinity sensitivity and a greater chalkiness in rice (Oryza sativa L.). Plant Physiol. Biochem. 2022, 193, 110–123. [Google Scholar] [CrossRef]
- Avora, F.; Eunier, A.; Ernet, A.V.; Ortefaix, M.P.; Milazzo, J.; Dreit, H.A.; Harreau, D.; Franco, O.L.; Mehta, A. CRISPR/Cas9-Targeted Knockout of Rice Susceptibility Genes OsDjA2 and OsERF104 Reveals Alternative Sources of Resistance to Pyricularia oryzae. Rice Sci. 2022, 29, 535–544. [Google Scholar] [CrossRef]
- Vescovi, V.; Kopp, W.; Guisan, J.M.; Giordano, R.; Mendes, A.A.; Tardioli, P.W. Improved catalytic properties of Candida antarctica lipase B multi-attached on tailor-made hydrophobic silica containing octyl and multifunctional amino- glutaraldehyde spacer arms. Process Biochem. 2016, 51, 2055–2066. [Google Scholar] [CrossRef]
- Park, J.R.; Kim, E.G.; Jang, Y.H.; Jan, R.; Farooq, M.; Ubaidillah, M.; Kim, K.M. Applications of CRISPR/Cas9 as New Strategies for Short Breeding to Drought Gene in Rice. Front. Plant Sci. 2022, 13, 850441. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Kong, M.; Yang, F.; Zhu, J.; Qi, X.; Weng, J.; Di, D.; Xie, C. Targeted generation of Null Mutants in ZmGDIα confers resistance against maize rough dwarf disease without agronomic penalty. Plant Biotechnol. J. 2022, 20, 803–805. [Google Scholar] [CrossRef] [PubMed]
- Blankenagel, S.; Eggels, S.; Frey, M.; Grill, E.; Bauer, E.; Dawid, C.; Fernie, A.R.; Haberer, G.; Hammerl, R.; Medeiros, D.B.; et al. Natural alleles of the abscisic acid catabolism gene ZmAbh4 modulate water use efficiency and carbon isotope discrimination in maize. Plant Cell 2022, 34, 3860–3872. [Google Scholar] [CrossRef]
- Liu, C.L.; Kong, M.; Zhu, J.J.; Qi, X.T.; Duan, C.X.; Xie, C.X. Engineering null mutants in ZmFER1 confers resistance to ear rot caused by Fusarium verticillioides in maize. Plant Biotechnol. J. 2022, 20, 2045–2047. [Google Scholar] [CrossRef]
- Qiang, Z.Q.; Sun, H.H.; Ge, F.H.; Li, W.; Li, C.J.; Wang, S.W.; Zhang, B.C.; Zhu, L.; Zhang, S.S.; Wang, X.Q.; et al. The transcription factor ZmMYB69 represses lignin biosynthesis by activating ZmMYB31/42 expression in maize. Plant Physiol. 2022, 189, 1916–1919. [Google Scholar] [CrossRef]
- Wada, N.; Ueta, R.; Osakabe, Y.; Osakabe, K. Precision genome editing in plants: State-of-the-art in CRISPR/Cas9-based genome engineering. BMC Plant Biol. 2020, 20, 234. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, R.; Ueta, R.; Abe, C.; Osakabe, Y.; Osakabe, K. Efficient Multiplex Genome Editing Induces Precise, and Self-Ligated Type Mutations in Tomato Plants. Front. Plant Sci. 2018, 9, 916. [Google Scholar] [CrossRef]
- Kishi-Kaboshi, M.; Aida, R.; Sasaki, K. Parsley ubiquitin promoter displays higher activity than the CaMV 35S promoter and the chrysanthemum actin 2 promoter for productive, constitutive, and durable expression of a transgene in Chrysanthemum morifolium. Breed. Sci. 2019, 69, 536–544. [Google Scholar] [CrossRef] [Green Version]
- Wolabu, T.W.; Park, J.J.; Chen, M.; Cong, L.L.; Ge, Y.X.; Jiang, Q.Z.; Debnath, S.; Li, G.M.; Wen, J.Q.; Wang, Z.Y. Improving the genome editing efficiency of CRISPR/Cas9 in Arabidopsis and Medicago truncatula. Planta 2020, 252, 15. [Google Scholar] [CrossRef]
- Li, J.; Wang, Z.; He, G.M.; Ma, L.G.; Deng, X.W. CRISPR/Cas9-mediated disruption of TaNP1 genes results in complete male sterility in bread wheat. J. Genet. Genom. 2020, 47, 263–272. [Google Scholar] [CrossRef]
- An, Y.; Geng, Y.; Yao, J.G.; Wang, C.; Du, J. An Improved CRISPR/Cas9 System for Genome Editing in Populus by Using Mannopine Synthase (MAS) Promoter. Front. Plant Sci. 2021, 12, 703546. [Google Scholar] [CrossRef] [PubMed]
- Massel, K.; Lam, Y.; Hintzsche, J.; Lester, N.; Botella, J.R.; Godwin, I.D. Endogenous U6 promoters improve CRISPR/Cas9 editing efficiencies in Sorghum bicolor and show potential for applications in other cereals. Plant Cell Rep. 2022, 41, 489–492. [Google Scholar] [CrossRef]
- Liu, S.; Wang, X.Y.; Li, Q.Q.; Peng, W.T.; Zhang, Z.M.; Chu, P.F.; Guo, S.J.; Fan, Y.L.; Lyu, S. AtGCS promoter-driven clustered regularly interspaced short palindromic repeats/Cas9 highly efficiently generates homozygous/biallelic mutations in the transformed roots by Agrobacterium rhizogenes-mediated transformation. Front. Plant Sci. 2022, 13, 952428. [Google Scholar] [CrossRef]
- Li, Q.; Sapkota, M.; van der Knaap, E. Perspectives of CRISPR/Cas-mediated cis-engineering in horticulture: Unlocking the neglected potential for crop improvement. Hortic. Res. Engl. 2020, 7, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, L.H.; Wei, S.W.; Wu, Y.R.; Hu, R.L.; Li, H.J.; Yang, W.C.; Xie, Q. High-Efficiency Genome Editing in Arabidopsis Using YAO Promoter-Driven CRISPR/Cas9 System. Mol. Plant. 2015, 8, 1820–1823. [Google Scholar] [CrossRef] [Green Version]
- Ordon, J.; Bressan, M.; Kretschmer, C.; Dall’Osto, L.; Marillonnet, S.; Bassi, R.; Stuttmann, J. Optimized Cas9 expression systems for highly efficient Arabidopsis genome editing facilitate isolation of complex alleles in a single generation. Funct. Integr. Genom. 2020, 20, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.; Li, T.; Dittman, J.D.; Su, J.B.; Li, R.Q.; Gassmann, W.; Peng, D.L.; Whitham, S.A.; Liu, S.M.; Yang, B. CRISPR/Cas9-Based Gene Editing Using Egg Cell-Specific Promoters in Arabidopsis and Soybean. Front. Plant Sci. 2020, 11, 800. [Google Scholar] [CrossRef]
- Kong, X.J.; Pan, W.B.; Sun, N.X.; Zhang, T.Y.; Liu, L.J.; Zhang, H.W. GLABRA2-based selection efficiently enriches Cas9-generated nonchimeric mutants in the T1 generation. Plant Physiol. 2021, 187, 758–768. [Google Scholar] [CrossRef]
- Wang, Y.J.; Lu, X.H.; Zhen, X.H.; Yang, H.; Che, Y.; Hou, J.Y.; Geng, M.T.; Liu, J.; Hu, X.W.; Li, R.M.; et al. A Transformation and Genome Editing System for Cassava Cultivar SC8. Genes 2022, 13, 1650. [Google Scholar] [CrossRef]
- Rahman, F.; Mishra, A.; Gupta, A.; Sharma, R. Spatiotemporal Regulation of CRISPR/Cas9 Enables Efficient, Precise, and Heritable Edits in Plant Genomes. Front. Genome Ed. 2022, 4, 870108. [Google Scholar] [CrossRef]
- Najera, V.A.; Twyman, R.M.; Christou, P.; Zhu, C.F. Applications of multiplex genome editing in higher plants. Curr. Opin. Biotechnol. 2019, 59, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Kor, S.D.; Chowdhury, N.; Keot, A.K.; Yogendra, K.; Chikkaputtaiah, C.; Sudhakar Reddy, P. RNA Pol III promoters-key players in precisely targeted plant genome editing. Front. Genet. 2022, 13, 989199. [Google Scholar] [CrossRef]
- Mao, Y.F.; Yang, X.X.; Zhou, Y.T.; Zhang, Z.J.; Botella, J.R.; Zhu, J.K. Manipulating plant RNA-silencing pathways to improve the gene editing efficiency of CRISPR/Cas9 systems. Genome Biol. 2018, 19, 149. [Google Scholar] [CrossRef]
- Wang, X.Y.; Lu, J.Y.; Lao, K.W.; Wang, S.K.; Mo, X.W.; Xu, X.T.; Chen, X.M.; Mo, B.X. Increasing the efficiency of CRISPR/Cas9-based gene editing by suppressing RNAi in plants. Sci. China Life Sci. 2019, 62, 982–984. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.C.; Li, S.Y.; Li, Z.L.; Li, H.Y.; Song, W.B.; Zhao, H.M.; Lai, J.S.; Xia, L.Q.; Li, D.W.; Zhang, Y.L. A barley stripe mosaic virus-based guide RNA delivery system for targeted mutagenesis in wheat and maize. Mol. Plant Pathol. 2019, 20, 1463–1474. [Google Scholar] [CrossRef]
- Tang, T.; Yu, X.W.; Yang, H.; Gao, Q.; Ji, H.T.; Wang, Y.X.; Yan, G.B.; Peng, Y.; Luo, H.F.; Liu, K.D.; et al. Development and Validation of an Effective CRISPR/Cas9 Vector for Efficiently Isolating Positive Transformants and Transgene-Free Mutants in a Wide Range of Plant Species. Front. Plant Sci. 2018, 9, 1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, B.L.; Moller, S.R.; Mravec, J.; Jorgensen, B.; Christensen, M.; Liu, Y.; Wandall, H.H.; Bennett, E.P.; Yang, Z. Improved CRISPR/Cas9 gene editing by fluorescence activated cell sorting of green fluorescence protein tagged protoplasts. BMC Biotechnol. 2019, 19, 36. [Google Scholar] [CrossRef]
- Trinh, D.D.; Le, N.T.; Bui, T.P.; Le, T.; Nguyen, C.X.; Chu, H.H.; Do, P.T. A sequential transformation method for validating soybean genome editing by CRISPR/Cas9 system. Saudi J. Biol. Sci. 2022, 29, 103420. [Google Scholar] [CrossRef]
- Milner, M.J.; Craze, M.; Hope, M.S.; Wallington, E.J. Turning Up the Temperature on CRISPR: Increased Temperature Can Improve the Editing Efficiency of Wheat Using CRISPR/Cas9. Front. Plant Sci. 2020, 11, 1780. [Google Scholar] [CrossRef]
- Liu, X.J.; Yang, J.T.; Song, Y.Y.; Zhang, X.C.; Wang, X.J.; Wang, Z.X. Effects of sgRNA length and number on gene editing efficiency and predicted mutations generated in rice. Crop J. 2022, 10, 577–581. [Google Scholar] [CrossRef]
- Khan, S.; Sallard, E. Current and Prospective Applications of CRISPR-Cas12a in Pluricellular Organisms. Mol. Biotechnol. 2022, 2, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Paul, B.; Montoya, G. CRISPR-Cas12a: Functional overview and applications. Biomed. J. 2020, 43, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, A.; Kancharla, N.; Javalkote, V.S.; Dasgupta, S.; Brutnell, T.P. CRISPR-Cas12a (Cpf1): A Versatile Tool in the Plant Genome Editing Tool Box for Agricultural Advancement. Front. Plant Sci. 2020, 11, 584151. [Google Scholar] [CrossRef]
- Nascimento, F.D.S.; Rocha, A.D.J.; Soares, J.M.D.S.; Mascarenhas, M.S.; Ferreira, M.D.S.; Morais Lino, L.S.; Ramos, A.P.D.S.; Diniz, L.E.C.; Mendes, T.A.D.O.; Ferreira, C.F.; et al. Gene Editing for Plant Resistance to Abiotic Factors: A Systematic Review. Plants 2023, 12, 305. [Google Scholar] [CrossRef]
- Hu, X.X.; Meng, X.B.; Li, J.Y.; Wang, K.J.; Yu, H. Improving the efficiency of the CRISPR-Cas12a system with tRNA-crRNA arrays. Crop J. 2020, 8, 403–407. [Google Scholar] [CrossRef]
- Jordan, W.T.; Currie, S.; Schmitz, R.J. Multiplex genome editing in Arabidopsis thaliana using Mb3Cas12a. Plant Direct 2021, 5, e344. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Geng, Y.; Yao, J.G.; Fu, C.X.; Lu, M.Z.; Wang, C.; Du, J. Efficient Genome Editing in Populus Using CRISPR/Cas12a. Front. Plant Sci. 2020, 11, 593938. [Google Scholar] [CrossRef]
- Wang, W.; Tian, B.; Pan, Q.L.; Chen, Y.Y.; He, F.; Bai, G.H.; Akhunova, A.; Trick, H.N.; Akhunov, E. Expanding the range of editable targets in the wheat genome using the variants of the Cas12a and Cas9 nucleases. Plant Biotechnol. J. 2021, 19, 2428–2441. [Google Scholar] [CrossRef]
- Nagy, E.D.; Kuehn, R.; Wang, D.F.; Shrawat, A.; Duda, D.M.; Groat, J.R.; Yang, P.Z.; Beach, S.; Zhang, Y.J.; Rymarquis, L.; et al. Site-directed integration of exogenous DNA into the soybean genome by LbCas12a fused to a plant viral HUH endonuclease. Plant J. 2022, 111, 905–916. [Google Scholar] [CrossRef]
- Pu, X.J.; Liu, L.N.; Li, P.; Huo, H.Q.; Dong, X.M.; Xie, K.B.; Yang, H.; Liu, L. A CRISPR/LbCas12a-based method for highly efficient multiplex gene editing in Physcomitrella patens. Plant J. 2019, 100, 863–872. [Google Scholar] [CrossRef]
- Yang, Z.; Xu, P. Implementing CRISPR-Cas12a for Efficient Genome Editing in Yarrowia lipolytica. Methods Mol. Biol. 2021, 2307, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Schroepfer, S.; Flachowsky, H. Tracing CRISPR/Cas12a Mediated Genome Editing Events in Apple Using High-Throughput Genotyping by PCR Capillary Gel Electrophoresis. Int. J. Mol. Sci. 2021, 22, 12611. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Liu, G.Q.; Zhou, J.P.; Ren, Q.R.; You, Q.; Tian, L.; Xin, X.H.; Zhong, Z.H.; Liu, B.L.; Zheng, X.L.; et al. A large-scale whole-genome sequencing analysis reveals highly specific genome editing by both Cas9 and Cpf1 (Cas12a) nucleases in rice. Genome Biol. 2018, 19, 84. [Google Scholar] [CrossRef] [Green Version]
- Bayat, H.; Modarressi, M.H.; Rahimpour, A. The Conspicuity of CRISPR-Cpf1 System as a Significant Breakthrough in Genome Editing. Curr. Microbiol. 2018, 75, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, J.S. Bypassing GMO regulations with CRISPR gene editing. Nat. Biotechnol. 2016, 34, 1014–1015. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Mo, J.; Zhao, X.; Li, G.; Zhang, X. CRISPR/Cas-mediated DNA base editing technology and its application in biomedicine and agriculture. Sheng Wu Gong Cheng Xue Bao Chin. J. Biotechnol. 2021, 37, 3071–3087. [Google Scholar] [CrossRef]
- Molla, K.A.; Sretenovic, S.; Bansal, K.C.; Qi, Y.P. Precise plant genome editing using base editors and prime editors. Nat. Plants 2021, 7, 1166–1187. [Google Scholar] [CrossRef]
- Hua, K.; Han, P.J.; Zhu, J.K. Improvement of base editors and prime editors advances precision genome engineering in plants. Plant Physiol. 2022, 188, 1795–1810. [Google Scholar] [CrossRef]
- Monsur, M.B.; Shao, G.N.; Lv, Y.S.; Ahmad, S.; Wei, X.J.; Hu, P.S.; Tang, S.Q. Base Editing: The Ever Expanding Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) Tool Kit for Precise Genome Editing in Plants. Genes 2020, 11, 466. [Google Scholar] [CrossRef]
- Li, J.; Zhang, C.; He, Y.; Li, S.; Yan, L.; Li, Y.; Zhu, Z.; Xia, L. Plant base editing and prime editing: The current status and future perspectives. J. Integr. Plant Biol. 2022, 65, 444–467. [Google Scholar] [CrossRef]
- Mishra, R.; Joshi, R.K.; Zhao, K.J. Base editing in crops: Current advances, limitations and future implications. Plant Biotechnol. J. 2020, 18, 20–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.S.; Ma, R.; Liu, D.; Wang, M.Y.; Zhu, T.; Deng, Y.X. A straightforward plant prime editing system enabled highly efficient precise editing of rice Waxy gene. Plant Sci. 2022, 323, 111400. [Google Scholar] [CrossRef]
- Gaillochet, C.; Pena Fernandez, A.; Goossens, V.; D’Halluin, K.; Drozdzecki, A.; Shafie, M.; Van Duyse, J.; Van Isterdael, G.; Gonzalez, C.; Vermeersch, M.; et al. Systematic optimization of Cas12a base editors in wheat and maize using the ITER platform. Genome Biol. 2023, 24, 6. [Google Scholar] [CrossRef] [PubMed]
- Pausch, P.; Al-Shayeb, B.; Bisom-Rapp, E.; Tsuchida, C.A.; Li, Z.; Cress, B.F.; Knott, G.J.; Jacobsen, S.E.; Banfield, J.F.; Doudna, J.A. CRISPR-Cas Phi from huge phages is a hypercompact genome editor. Science 2020, 369, 333. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Sretenovic, S.; Fan, T.; Cheng, Y.; Li, G.; Qi, A.; Tang, X.; Xu, Y.; Guo, W.; Zhong, Z.; et al. Hypercompact CRISPR-Cas12j2 (CasPhi) enables genome editing, gene activation, and epigenome editing in plants. Plant Commun. 2022, 3, 100453. [Google Scholar] [CrossRef]
- Pausch, P.; Soczek, K.M.; Herbst, D.A.; Tsuchida, C.A.; Al-Shayeb, B.; Banfield, J.F.; Nogales, E.; Doudna, J.A. DNA interference states of the hypercompact CRISPR-Cas phi effector. Nat. Struct. Mol. Biol. 2021, 28, 652. [Google Scholar] [CrossRef]
- Anonymous. CRISPR-Csm for eukaryotic RNA knockdown and imaging without toxicity. Nat. Biotechnol. 2023. [Google Scholar] [CrossRef]
- Colognori, D.; Trinidad, M.; Doudna, J.A. Precise transcript targeting by CRISPR-Csm complexes. Nat. Biotechnol. 2023. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Luan, X.; Liu, Y.; Wang, L.; Wang, J.; Yang, S.; Liu, S.; Zhang, J.; Liu, H.; Yao, D. Strategies and Methods for Improving the Efficiency of CRISPR/Cas9 Gene Editing in Plant Molecular Breeding. Plants 2023, 12, 1478. https://doi.org/10.3390/plants12071478
Zhou J, Luan X, Liu Y, Wang L, Wang J, Yang S, Liu S, Zhang J, Liu H, Yao D. Strategies and Methods for Improving the Efficiency of CRISPR/Cas9 Gene Editing in Plant Molecular Breeding. Plants. 2023; 12(7):1478. https://doi.org/10.3390/plants12071478
Chicago/Turabian StyleZhou, Junming, Xinchao Luan, Yixuan Liu, Lixue Wang, Jiaxin Wang, Songnan Yang, Shuying Liu, Jun Zhang, Huijing Liu, and Dan Yao. 2023. "Strategies and Methods for Improving the Efficiency of CRISPR/Cas9 Gene Editing in Plant Molecular Breeding" Plants 12, no. 7: 1478. https://doi.org/10.3390/plants12071478
APA StyleZhou, J., Luan, X., Liu, Y., Wang, L., Wang, J., Yang, S., Liu, S., Zhang, J., Liu, H., & Yao, D. (2023). Strategies and Methods for Improving the Efficiency of CRISPR/Cas9 Gene Editing in Plant Molecular Breeding. Plants, 12(7), 1478. https://doi.org/10.3390/plants12071478