
Transcriptomic and Physiological Analyses Reveal the Molecular Mechanism through Which Exogenous Melatonin Increases Drought Stress Tolerance in Chrysanthemum

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
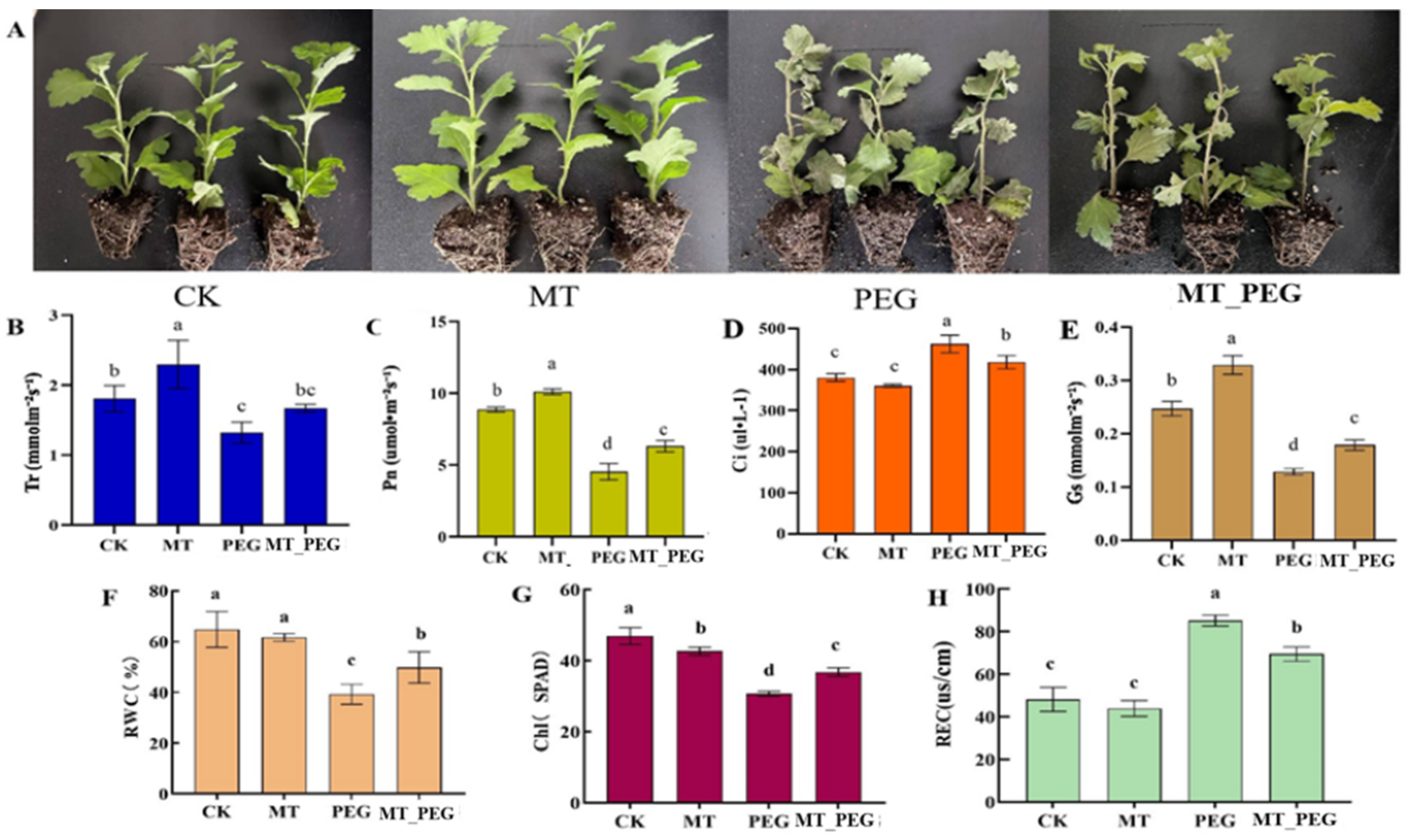
2.1. Effects of Melatonin on Chrysanthemum Seedlings under Drought Stress
2.2. Effects of Melatonin on Oxidative Damage and Antioxidant Activity under Drought Stress
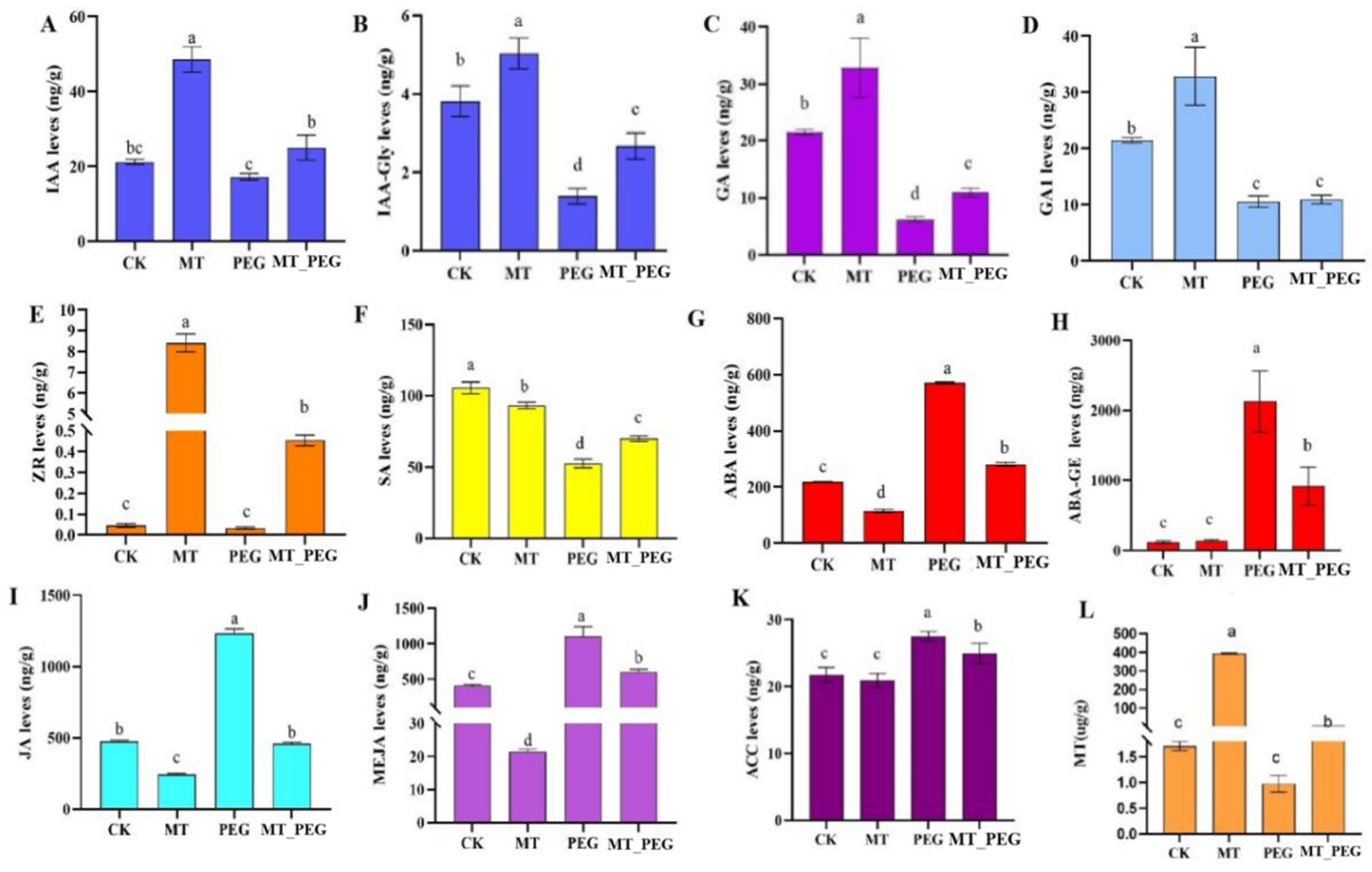
2.3. Effects of Melatonin on the Synthesis of Endogenous Hormones and the Melatonin Content of Chrysanthemum under Drought Stress
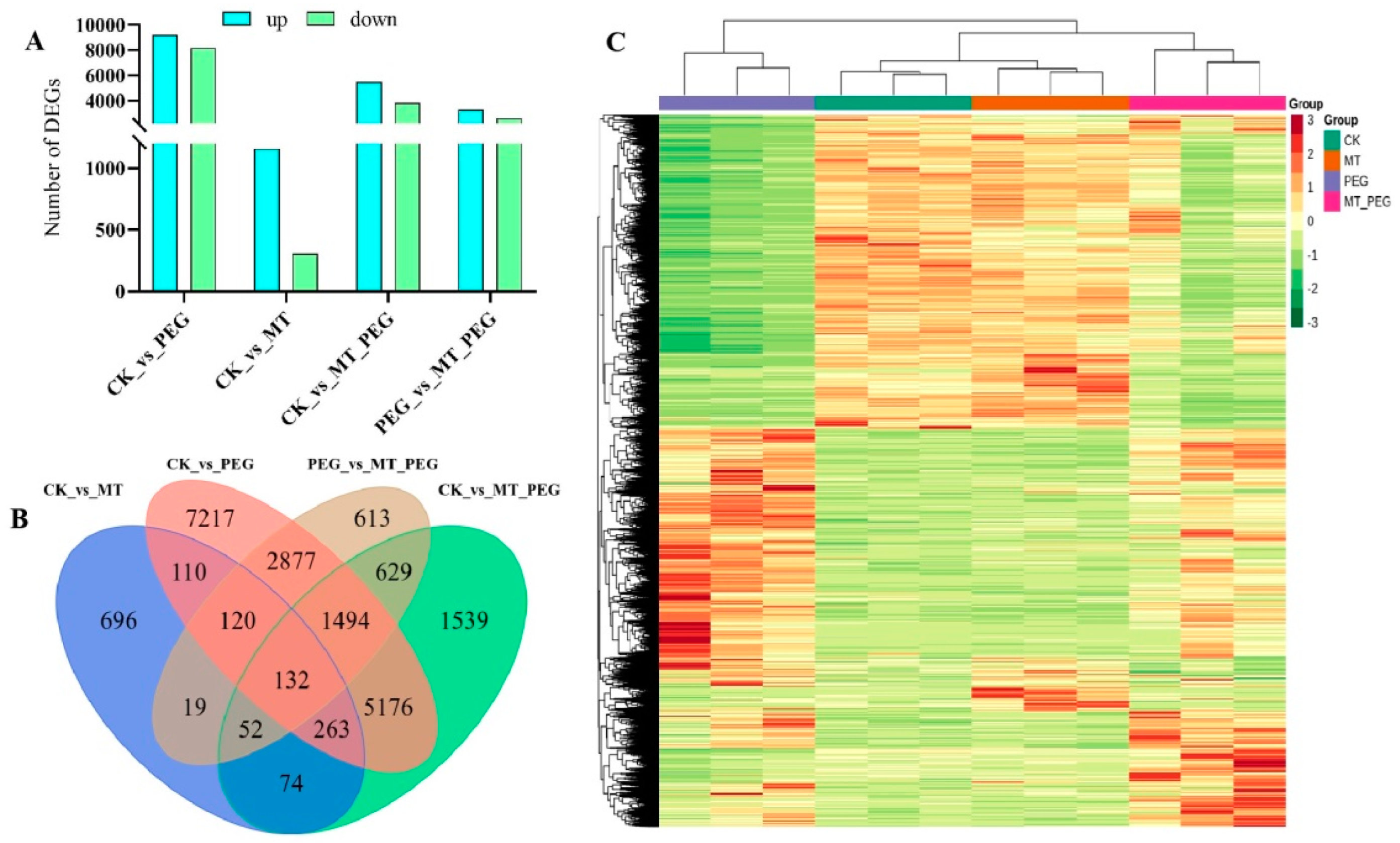
2.4. Transcriptome Analysis of Differential Gene Expression
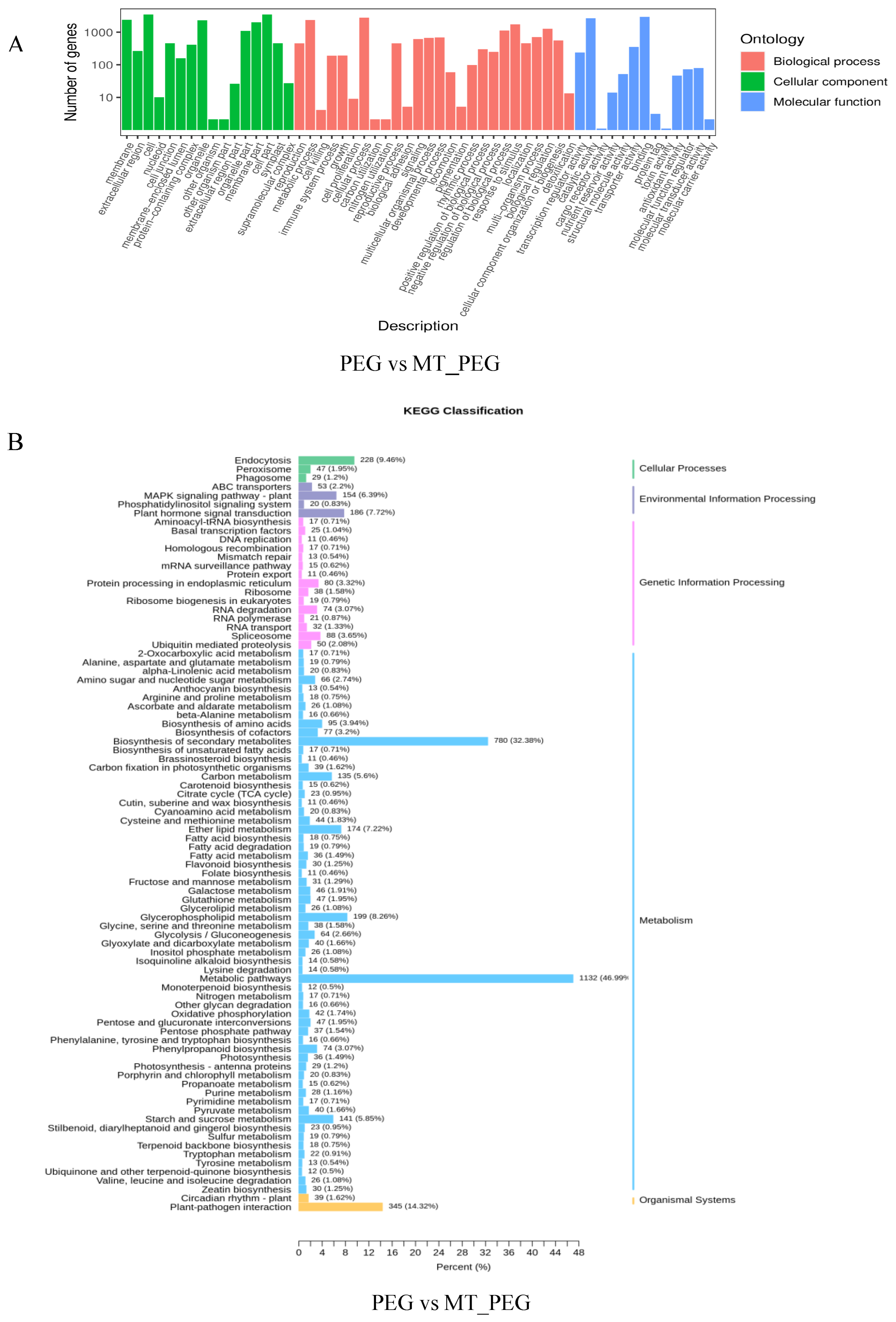
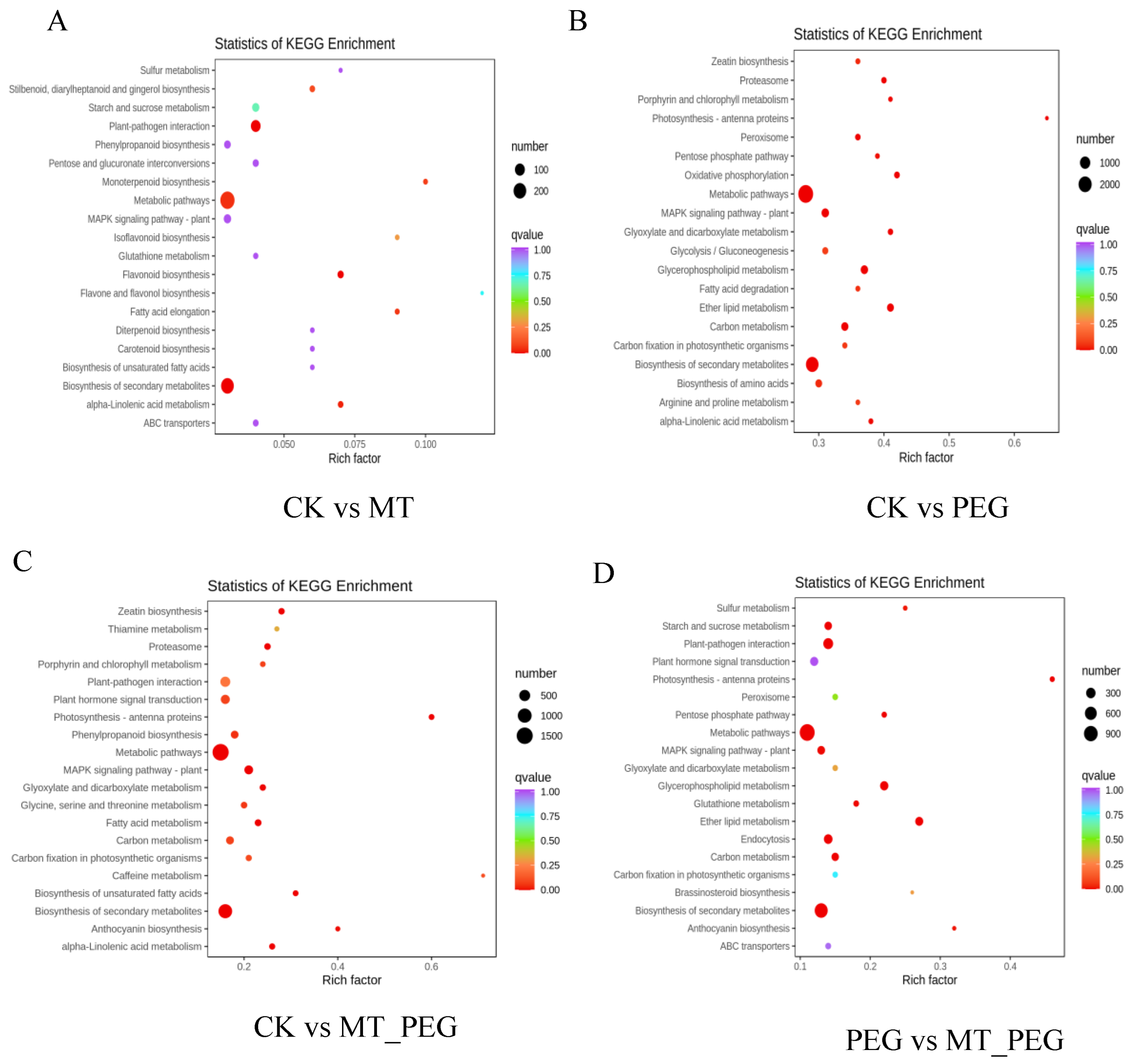
2.5. GO Classification and KEGG Analyses
2.6. KEGG Metabolic Pathway Analysis of DEGs
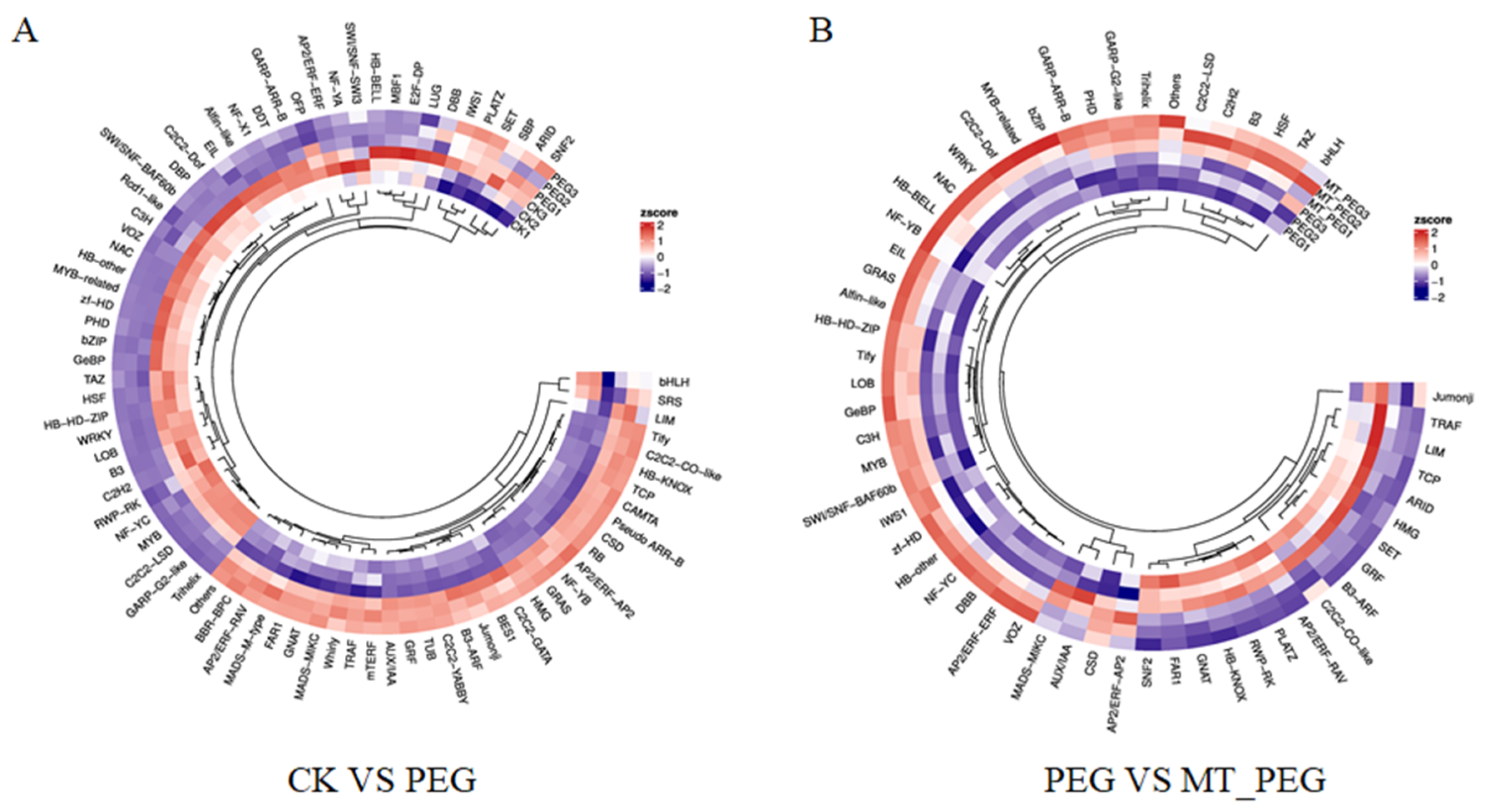
2.7. Effects of Exogenous MT on Transcription Factors in Chrysanthemum under Drought Stress
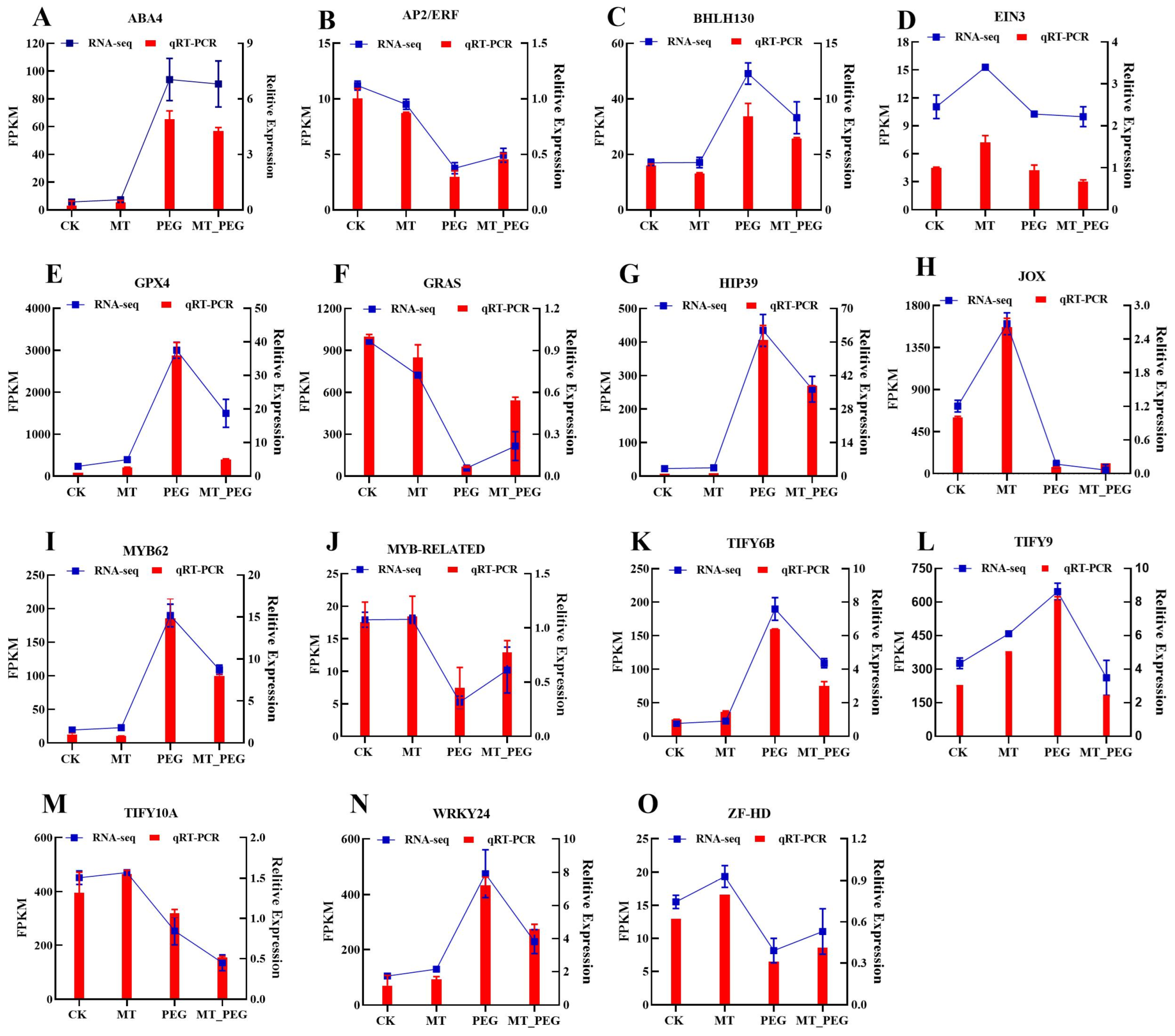
2.8. Validation of Sequencing Data by Quantitative Real-Time PCR (qPCR) Analysis
3. Discussion
4. Material and Methods
4.1. Plant Materials and Treatments
4.2. Determination of Physiological Indicators such as Light and Plant Characteristics and Chlorophyll and Antioxidant System Contents
4.3. Determination of Endogenous Hormone and Melatonin Contents
4.4. Transcriptome Sequencing and RNA-Seq Data Analysis
4.5. Quantitative Real-Time PCR (qPCR) Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gong, Z.Z.; Xiong, L.M.; Shi, H.Z.; Yang, S.H.; Herrera-Estrella, L.R.; Xu, G.H.; Chao, D.Y.; Li, J.R.; Wang, P.Y.; Qin, F.; et al. Plant abiotic stress response and nutrient use efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [PubMed]
- Gupta, A.; Rico-Medina, A.; Cano-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Socha, A.L.; Guerinot, M.L. Mn-euvering manganese: The role of transporter gene family members in manganese uptake and mobilization in plants. Front. Plant Sci. 2014, 5, 106. [Google Scholar] [CrossRef] [Green Version]
- Teixeira da Silva, J.A. Chrysanthemum: Advances in tissue culture, cryopreservation, postharvest technology, genetics and transgenic biotechnology. Biotechnol. Adv. 2003, 21, 715–766. [Google Scholar] [CrossRef]
- Bhattacharya, A.; da Silva, J.A.T. Molecular systematics in Chrysanthemum × grandiflorum (Ramat.) Kitamura. Sci. Hortic. 2006, 109, 379–384. [Google Scholar] [CrossRef]
- Chinese Pharmmacopoeia Editorial Committee. Pharmmacopoeia of the People’s Republic of China; Chemical Industry Press: Beijing, China, 2010. [Google Scholar]
- Liang, W.Y. Screening and Evaluation of Hybrid Progeny of Garden Chrysanthemum × ‘Huangju’. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2020. [Google Scholar] [CrossRef]
- Wang, T.L.; Wei, Q.; Wang, Z.L.; Liu, W.W.; Zhao, X.; Ma, C.; Gao, J.P.; Xu, Y.J.; Hong, B. CmNF-YB8 affects drought resistance in Chrysanthemum by altering stomatal status and leaf cuticle thickness. J. Integr. Plant Biol. 2022, 64, 741–755. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.J.; Zhao, X.; Aiwaili, P.; Mu, X.Y.; Zhao, M.; Zhao, J.; Cheng, L.N.; Ma, C.; Gao, J.P.; Hong, B. A zinc finger protein BBX19 interacts with ABF3 to affect drought tolerance negatively in Chrysanthemum. Plant J. 2020, 103, 1783–1795. [Google Scholar] [CrossRef]
- Hong, B.; Tong, Z.; Ma, N.; Li, J.K.; Kasuga, M.; Yamaguchi-Shinozaki, K.; Gao, J.P. Heterologous expression of the AtDREB1A gene in Chrysanthemum increases drought and salt stress tolerance. Sci. China Life Sci. 2006, 49, 436–445. [Google Scholar] [CrossRef]
- Chen, L.; Chen, Y.; Jiang, J.F.; Chen, S.M.; Chen, F.D.; Guan, Z.Y.; Fang, W.M. The constitutive expression of Chrysanthemum dichrum ICE1 in Chrysanthemum grandiflorum improves the level of low temperature, salinity and drought tolerance. Plant Cell Rep. 2012, 31, 1747–1758. [Google Scholar] [CrossRef]
- Yang, Y.J.; Ma, C.; Xu, Y.J.; Wei, Q.; Imtiaz, M.; Lan, H.B.; Gao, S.; Cheng, L.N.; Wang, M.Y.; Fei, Z.J. A zinc finger protein regulates flowering time and abiotic stress tolerance in Chrysanthemum by modulating gibberellin biosynthesis. Plant Cell 2014, 26, 2038–2054. [Google Scholar] [CrossRef] [Green Version]
- Jaffar, M.A.; Song, A.P.; Faheem, M.; Chen, S.M.; Jiang, J.F.; Liu, C.; Fan, Q.Q.; Chen, F.D. Involvement of CmWRKY10 in drought tolerance of Chrysanthemum through the ABA-signaling pathway. Int. J. Mol. Sci. 2016, 17, 693. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Zhong, M.; He, L.; Wang, B.; Liu, Q.L.; Pan, Y.Z.; Jiang, B.B.; Zhang, L. Overexpression of a Chrysanthemum transcription factor gene DgNAC1 improves drought tolerance in Chrysanthemum. Plant Cell Tissue Organ Cult. (PCTOC) 2018, 135, 119–132. [Google Scholar] [CrossRef]
- Liu, Y.N.; Chen, H.; Ping, Q.; Zhang, Z.; Guan, Z.Z.; Fang, W.M.; Chen, S.M.; Chen, F.D.; Jiang, J.F.; Zhang, F. The heterologous expression of CmBBX22 delays leaf senescence and improves drought tolerance in Arabidopsis. Plant Cell Rep. 2019, 38, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Kanwar, M.K.; Yu, J.Q.; Zhou, J. Phytomelatonin: Recent advances and future prospects. J. Pineal Res. 2018, 65, e12526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: A new plant hormone and/or a plant master regulator? Trends Plant Sci. 2019, 24, 38–48. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Lal, M.K.; Kumar, R.; Chourasia, K.N.; Naga, K.C.; Kumar, D.; Das, S.K.; Zinta, G. Mechanistic insights on melatonin-mediated drought stress mitigation in plants. Plant Physiol. 2021, 172, 1212–1226. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.P.; Reiter, R.J.; Chan, Z.L. Phytomelatonin: A universal abiotic stress regulator. J. Exp. Bot. 2018, 69, 963–974. [Google Scholar] [CrossRef] [Green Version]
- Han, Q.H.; Huang, B.; Ding, C.B.; Zhang, Z.W.; Chen, Y.E.; Hu, C.; Zhou, L.J.; Huang, Y.; Liao, J.Q.; Yuan, S.; et al. Effects of melatonin on anti-oxidative systems and photosystem II in cold-stressed rice seedlings. Front. Plant Sci. 2017, 8, 785. [Google Scholar] [CrossRef]
- Ke, Q.B.; Ye, J.; Wang, B.M.; Ren, J.H.; Yin, L.N.; Deng, X.P.; Wang, S.W. Melatonin mitigates salt stress in wheat seedlings by modulating polyamine metabolism. Front. Plant Sci. 2018, 9, 914. [Google Scholar] [CrossRef] [Green Version]
- Li, X.N.; Tan, D.X.; Jiang, D.; Liu, F.L. Melatonin enhances cold tolerance in drought-primed wild-type and abscisic acid-deficient mutant barley. J. Pineal Res. 2016, 61, 328–339. [Google Scholar] [CrossRef]
- Zhang, R.M.; Sun, Y.K.; Liu, Z.Y.; Jin, W.; Sun, Y. Effects of melatonin on seedling growth, mineral nutrition, and nitrogen metabolism in cucumber under nitrate stress. J. Pineal Res. 2017, 62. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Li, Q.T.; Chu, Y.N.; Reiter, R.J.; Yu, X.M.; Zhu, D.H.; Zhang, W.K.; Ma, B.; Lin, Q.; Zhang, J.S.; et al. Melatonin enhances plant growth and abiotic stress tolerance in soybean plants. J. Exp. Bot. 2015, 66, 695–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.L.; Xiang, G.Q.; Sun, Q.H.; Ni, Y.; Jin, Z.X.; Gao, S.W.; Yao, Y.X. Melatonin enhances salt tolerance by promoting MYB108A-mediated ethylene biosynthesis in grapevines. Hortic. Res. 2019, 6, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoniou, C.; Chatzimichail, G.; Xenofontos, R.; Pavlou, J.J.; Panagiotou, E.; Christou, A.; Fotopoulos, V. Melatonin systemically ameliorates drought stress-induced damage in Medicago sativa plants by modulating nitro-oxidative homeostasis and proline metabolism. J. Pineal Res. 2017, 62. [Google Scholar] [CrossRef]
- Luo, Y.; Hu, T.T.; Gao, X.F.; Zhao, Y.L.; Zhang, L.; Yan, R. Effects of exogenous melatonin on growth and physiology of garden chrysanthemum under drought stress. J. Gansu Agric. Univ. 2023, 1–11. Available online: https://kns.cnki.net/kcms2/article/abstract?v=3uoqIhG8C45S0n9fL2suRadTyEVl2pW9UrhTDCdPD65KmEWR38_iwZWXuAxF9_V3PXR4vLgakRGRrMEqji7aucQGfYrLODyA&uniplatform=NZKPT (accessed on 14 March 2023).
- Trebst, A.; Depka, B.; Holländer-Czytko, H. A specific role for tocopherol and of chemical singlet oxygen quenchers in the maintenance of photosystem II structure and function in Chlamydomonas reinhardtii. FEBS Lett. 2002, 516, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Ali, B. Brassinosteroids: The promising plant growth regulators in horticulture. In Brassinosteroids: Plant Growth and Development; Springer: Singapore, 2019; pp. 349–365. [Google Scholar]
- Zhang, N.; Zhao, B.; Zhang, H.J.; Weeda, S.; Yang, C.; Yang, Z.C.; Ren, S.X.; Guo, Y.D. Melatonin promotes water-stress tolerance, lateral root formation, and seed germination in cucumber (Cucumis sativus L.). J. Pineal Res. 2013, 54, 15–23. [Google Scholar] [CrossRef]
- Khan, I.; Awan, S.A.; Ikram, R.; Rizwan, M.; Akhtar, N.; Yasmin, H.; Sayyed, R.Z.; Ali Shafaqat Ilyas, N. Effects of 24-epibrassinolide on plant growth, antioxidants defense system, and endogenous hormones in two wheat varieties under drought stress. Physiol. Plant. 2021, 172, 696–706. [Google Scholar] [CrossRef]
- Hardeland, R.; Cardinali, D.P.; Srinivasan, V.; Spence, D.W.; Brown, G.M.; Pandi-Perumal, S.R. Melatonin—A pleiotropic, orchestrating regulator molecule. Prog. Neurobiol. 2011, 93, 350–384. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, M.; Tavakol, I.A.; Pouryousef, M.; Fard, E.M. Study the effect of 24-epibrassinolide application on the Cu/Zn-SOD expression and tolerance to drought stress in common bean. Physiol. Mol. Biol. Plants 2020, 26, 459–474. [Google Scholar] [CrossRef]
- Tan, D.X.; Reiter, R.J.; Manchester, L.C.; Yan, M.T.; El-Sawi, M.; Sainz, R.M.; Mayo, J.C.; Kohen, R.; Allegra, M.C.; Hardeland, R. Chemical and physical properties and potential mechanisms: Melatonin as a broad spectrum antioxidant and free radical scavenger. Curr. Top. Med. Chem. 2002, 2, 181–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.B. Physiological Response and Proteomic Analysis of Wheat Seedlings to Melatonin Treatment under Drought and PEG Stress. Ph.D. Thesis, Northwest A & F University, Xianyang, China, 2019. [Google Scholar]
- Zhang, H.J.; Zhang, N.A.; Yang, R.C.; Wang, L.; Sun, Q.Q.; Li, D.B.; Cao, Y.Y.; Weeda, S.; Zhao, B.; Ren, S.X.; et al. Melatonin promotes seed germination under high salinity by regulating antioxidant systems, ABA and GA4 interaction in cucumber (Cucumis sativus L.). J. Pineal Res. 2014, 57, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.X.; Zhang, T.; Zhang, P.; Wang, Z.Y. Melatonin attenuates postharvest physiological deterioration of cassava storage roots. J. Pineal Res. 2016, 60, 424–434. [Google Scholar] [CrossRef]
- Ye, J.; Wang, S.W.; Deng, X.P.; Yin, L.N.; Xiong, B.L.; Wang, X.Y. Melatonin increased maize (Zea mays L.) seedling drought tolerance by alleviating drought-induced photosynthetic inhibition and oxidative damage. Acta Physiol. Plant. 2016, 38. [Google Scholar] [CrossRef]
- Zhang, N.; Sun, Q.; Zhang, H.; Cao, Y.; Weeda, S.; Ren, S.; Guo, Y.D. Roles of melatonin in abiotic stress resistance in plants. J. Exp. Bot. 2015, 66, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Jiang, C.; Ye, T.T.; Tan, D.X.; Reiter, R.J.; Zhang, H.; Liu, R.Y.; Chan, Z.L. Comparative physiological, metabolomic, and transcriptomic analyses reveal mechanisms of improved abiotic stress resistance in bermudagrass [Cynodon dactylon (L). Pers.] by exogenous melatonin. J. Exp. Bot. 2015, 66, 681–694. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.; Kamran, M.; Ding, R.X.; Meng, X.P.; Wang, H.Q.; Ahmad, I.; Fahad, S.; Han, Q.F. Exogenous melatonin confers drought stress by promoting plant growth, photosynthetic capacity and antioxidant defense system of maize seedlings. Peer J. 2019, 7, e7793. [Google Scholar] [CrossRef] [Green Version]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Zhao, C.F.; Wang, C.G.; Li, H.J.; Zheng, X.H.; Yang, M.; Zhang, R.H. Effects of exogenous melatonin on photosynthesis of maize leaves under drought and rehydration conditions. Acta Ecol. Sin. 2021, 41, 1431–1439. [Google Scholar]
- Llanes, A.; Andrade, A.; Alemano, S.; Luna, V. Alterations of endogenous hormonal levels in plants under drought and salinit. Am. J. Plant Sci. 2016, 7, 1357–1371. [Google Scholar] [CrossRef] [Green Version]
- Pirasteh, A.H.; Emam, Y.; Pessarakli, M. Changes in Endogenous Hormonal Status in Corn (Zea mays) Hybrids Under Drought Stress. J. Plant Nutr. 2013, 36, 1695–1707. [Google Scholar] [CrossRef]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant. Boil. 2003, 6, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Liu, R.; Sui, N.; Shi, W.W.; Wang, L.; Tian, C.Y.; Song, J. Changes in endogenous hormones and seed-coat phenolics during seed storage of two Suaeda salsa populations. Aust. J. Bot. 2016, 64, 325–332. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, Y.; Wei, X.; Zhao, X.; Wang, B. Transcription Profiles of Genes Related to Hormonal Regulations Under Salt Stress in Sweet Sorghum. Plant. Mol. Biol. Rep. 2017, 35, 586–599. [Google Scholar] [CrossRef]
- Muhammad, A.; Shao, Y.H.; Liu, S.X.; Wang, F.; Gao, J.W.; Jiang, D.; Tian, Z.W.; Dai, T.B. Pre-drought priming sustains grain development under post-anthesis drought stress by regulating the growth hormones in winter wheat (Triticum aestivum L.). Planta 2017, 246, 509–524. [Google Scholar]
- Zhou, Q.; Li, Y.P.; Wang, X.J.; Yan, C.; Ma, C.M.; Liu, J.; Dong, S.K. Effects of Different Drought Degrees on Physiological Characteristics and Endogenous Hormones of Soybean. Plants 2022, 11, 2282. [Google Scholar] [CrossRef] [PubMed]
- Fleta-Soriano, E.; Día, L.; Bonet, E.; Munne-Bosch, S. Melatonin may exert a protective role against drought stress in maize. J. Agron. Crop Sci. 2017, 203, 286–294. [Google Scholar] [CrossRef]
- Bhusal, N.; Han, S.G.; Yoon, T.M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Zhou, Y.F.; Wang, D.Q.; Lu, Z.S.; Wang, N.; Wang, Y.T.; Li, F.X.; Xu, J.; Huang, R.D. Effects of drought stress on photosynthetic characteristics and endogenous hormones ABA and CTK content of stay-green sorghum. Chin. Agric. Sci. 2014, 47, 655–663. [Google Scholar]
- Elizabeth, A.M.; Sergi, M.B. Photo- and antioxidant protection and salicylic acid accumulation during post-anthesis leaf senescence in Salvia lanigera grown under Mediterranean climate. Physiol. Plant. 2007, 131, 590–598. [Google Scholar]
- Zhou, R.; Yu, X.Q.; Zhao, T.M.; Ottosen, C.O.; Rosenqvist, E.; Wu, Z. Physiological analysis and transcriptome sequencing reveal the effects of combined cold and drought on tomato leaf. BMC Plant Biol. 2019, 19, 377. [Google Scholar] [CrossRef]
- Debnath, B.; Li, M.; Liu, S.; Pan, T.F.; Ma, C.L.; Qiu, D.L. Melatonin-mediate acid rain stress tolerance mechanism through alteration of transcriptional factors and secondary metabolites gene expression in tomato. Ecotoxicol. Environ. Saf. 2020, 200, 110720. [Google Scholar] [CrossRef]
- Baillo, E.H.; Kimotho, R.N.; Zhang, Z.B.; Xu, P. Transcription factors associated with abiotic and biotic stress tolerance and their potential for crops improvement. Genes 2019, 10, 771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Wei, Y.X.; He, C.Z. Melatonin-induced CBF/DREB1s are essential for diurnal change of disease resistance and CCA1 expression in Arabidopsis. Plant Physiol. Biochem. 2016, 100, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Eulgem, T.; Rushton, P.J.; Robatzek, S.; Somssich, I.E. The WRKY superfamily of plant transcription factors. Trends Plant Sci. 2000, 5, 199–220. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Chen, K.; Wei, Y.; He, C. Fundamental issues of melatonin-mediated stress signaling in plants. Front. Plant Sci. 2016, 7, 1124. [Google Scholar] [CrossRef] [Green Version]
- Turk, H.; Erdal, S. Melatonin alleviates cold-induced oxidative damage in maize seedlings by up-regulating mineral elements and enhancing antioxidant activity. J. Plant Nutr. Soil Sci. 2015, 178, 433–439. [Google Scholar] [CrossRef]
- Gao, J.F. Plant Physiology Laboratory Guide; Higher Education Press: Beijing, China, 2006. [Google Scholar]
- Zhao, S.J.; Xu, C.C.; Zou, Q.; Meng, Q.W. Improvement of the method for determination of malondialdehyde in plant tissues. Plant Physiol. Lett. 1994, 03, 207–210. [Google Scholar]
- Li, H.S. Principles and Techniques of Plant Physiological and Biochemical Experiments; Higher Education Press: Beijing, China, 2000. [Google Scholar]
- Leng, F.; Sun, S.; Jing, Y.; Wang, F.; Wei, Q.; Wang, X. A rapid and sensitive method for determination of trace amounts of glucose by anthrone-sulfuric acid method. Bulg. Chem. Commun. 2016, 48, 109–113. [Google Scholar]
- Sedmak, J.J.; Grossberg, S.E. A rapid, sensitive, and versatile assay for protein using Coomassie brilliant blue G250. Anal. Biochem. 1977, 79, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for rna-seq data with deseq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varet, H.; Brillet-Guéguen, L.; Coppée, J.Y.; Dillies, M.A. SARTools: A DESeq2- and EdgeR-based R pipeline for comprehensive differential analysis of RNA-seq data. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adnan, M.; Morton, G.; Hadi, S. Analysis of rpoS and bolA gene expression under various stress-induced environments in planktonic and biofilm phase using 2−ΔΔCT method. Mol. Cell. Biochem. 2011, 357, 275–282. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Y.; Hu, T.; Huo, Y.; Wang, L.; Zhang, L.; Yan, R. Transcriptomic and Physiological Analyses Reveal the Molecular Mechanism through Which Exogenous Melatonin Increases Drought Stress Tolerance in Chrysanthemum. Plants 2023, 12, 1489. https://doi.org/10.3390/plants12071489
Luo Y, Hu T, Huo Y, Wang L, Zhang L, Yan R. Transcriptomic and Physiological Analyses Reveal the Molecular Mechanism through Which Exogenous Melatonin Increases Drought Stress Tolerance in Chrysanthemum. Plants. 2023; 12(7):1489. https://doi.org/10.3390/plants12071489
Chicago/Turabian StyleLuo, Yan, Taotao Hu, Yunyun Huo, Lingling Wang, Li Zhang, and Rui Yan. 2023. "Transcriptomic and Physiological Analyses Reveal the Molecular Mechanism through Which Exogenous Melatonin Increases Drought Stress Tolerance in Chrysanthemum" Plants 12, no. 7: 1489. https://doi.org/10.3390/plants12071489
APA StyleLuo, Y., Hu, T., Huo, Y., Wang, L., Zhang, L., & Yan, R. (2023). Transcriptomic and Physiological Analyses Reveal the Molecular Mechanism through Which Exogenous Melatonin Increases Drought Stress Tolerance in Chrysanthemum. Plants, 12(7), 1489. https://doi.org/10.3390/plants12071489