
Development of an In-Field Real-Time LAMP Assay for Rapid Detection of Tomato Leaf Curl New Delhi Virus

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of Viral Material
2.2. AV1 Gene Amplification by PCR
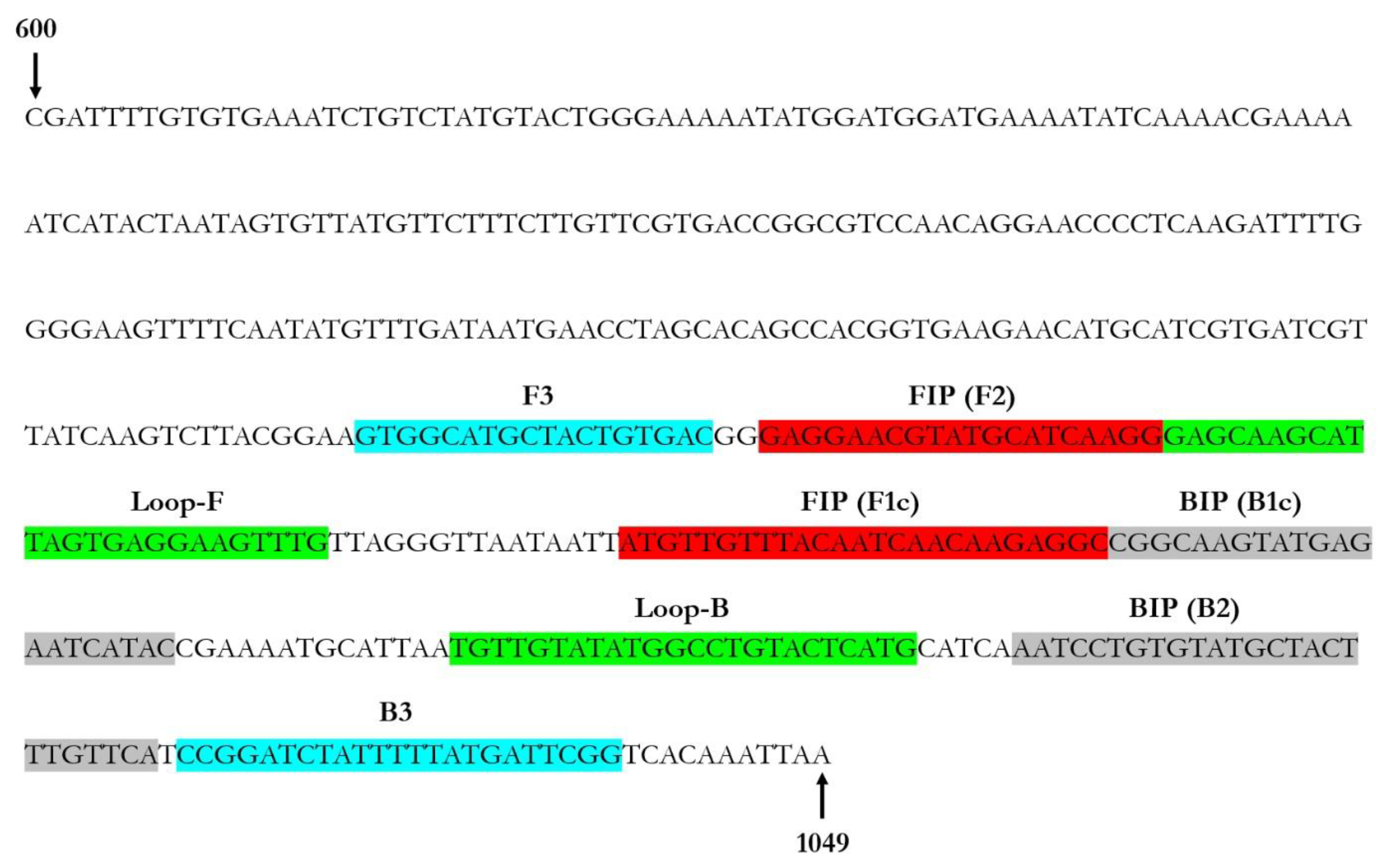
2.3. Primer Design for LAMP Assay Set-Up
2.4. ToLCNDV Real-Time LAMP Assay Optimisation
2.5. Sensitivity, Reaction Time, and Specificity of Real-Time LAMP Assay and Comparison to End-Point PCR
2.6. Rapid Sample Preparation Method Suitable for the ToLCNDV Real-Time LAMP Assay
2.7. Direct on Field Analysis of Tomato Leaf Curl New Delhi Virus
3. Results
3.1. AV1 Gene Amplification by PCR
3.2. Primers Design for LAMP Assay Set-Up
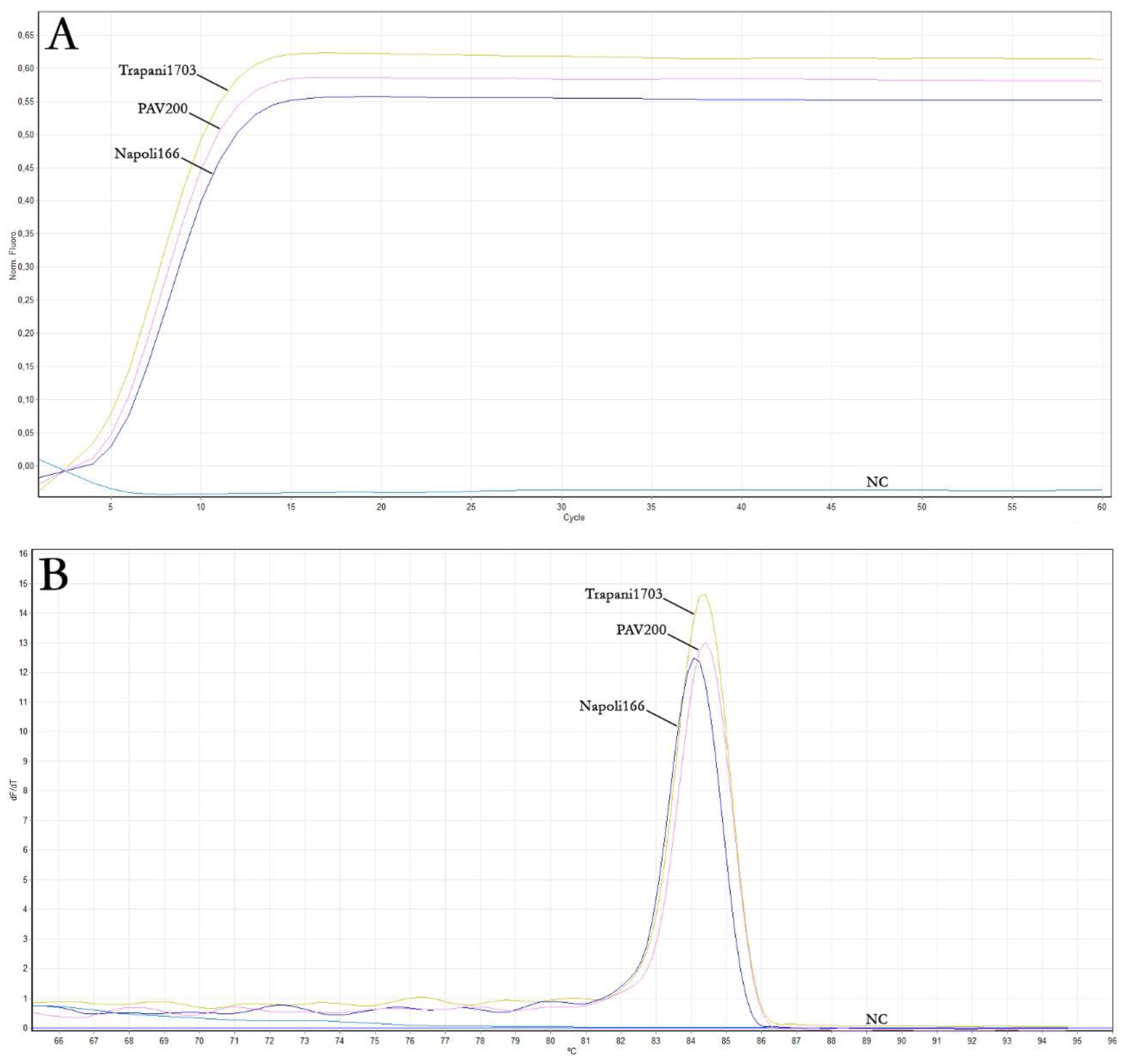
3.3. ToLCNDV Real-Time LAMP Assay Optimisation
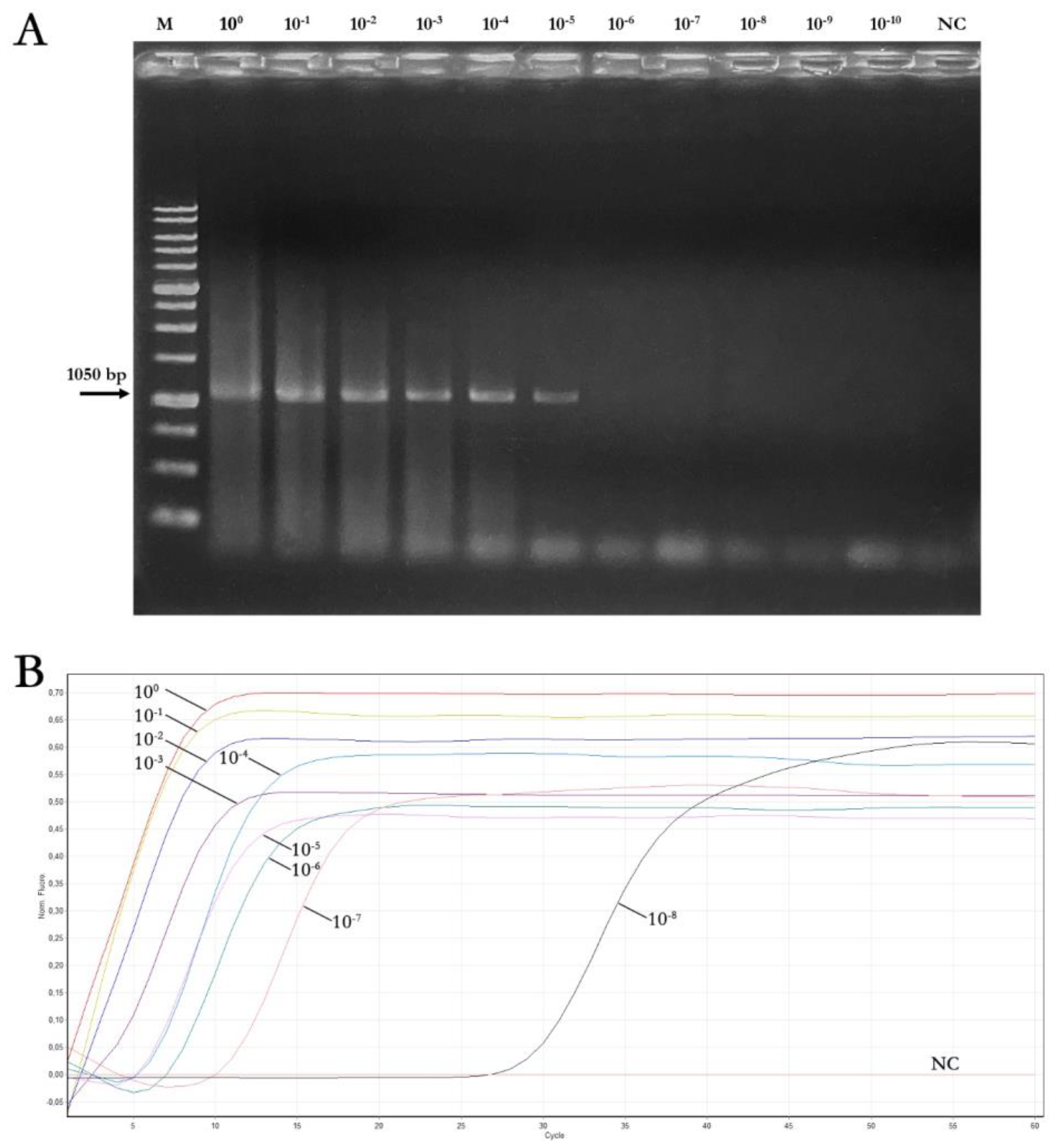
3.4. Sensitivity, Reaction Time, and Specificity of Real-Time LAMP Assay and Comparison to End-Point PCR
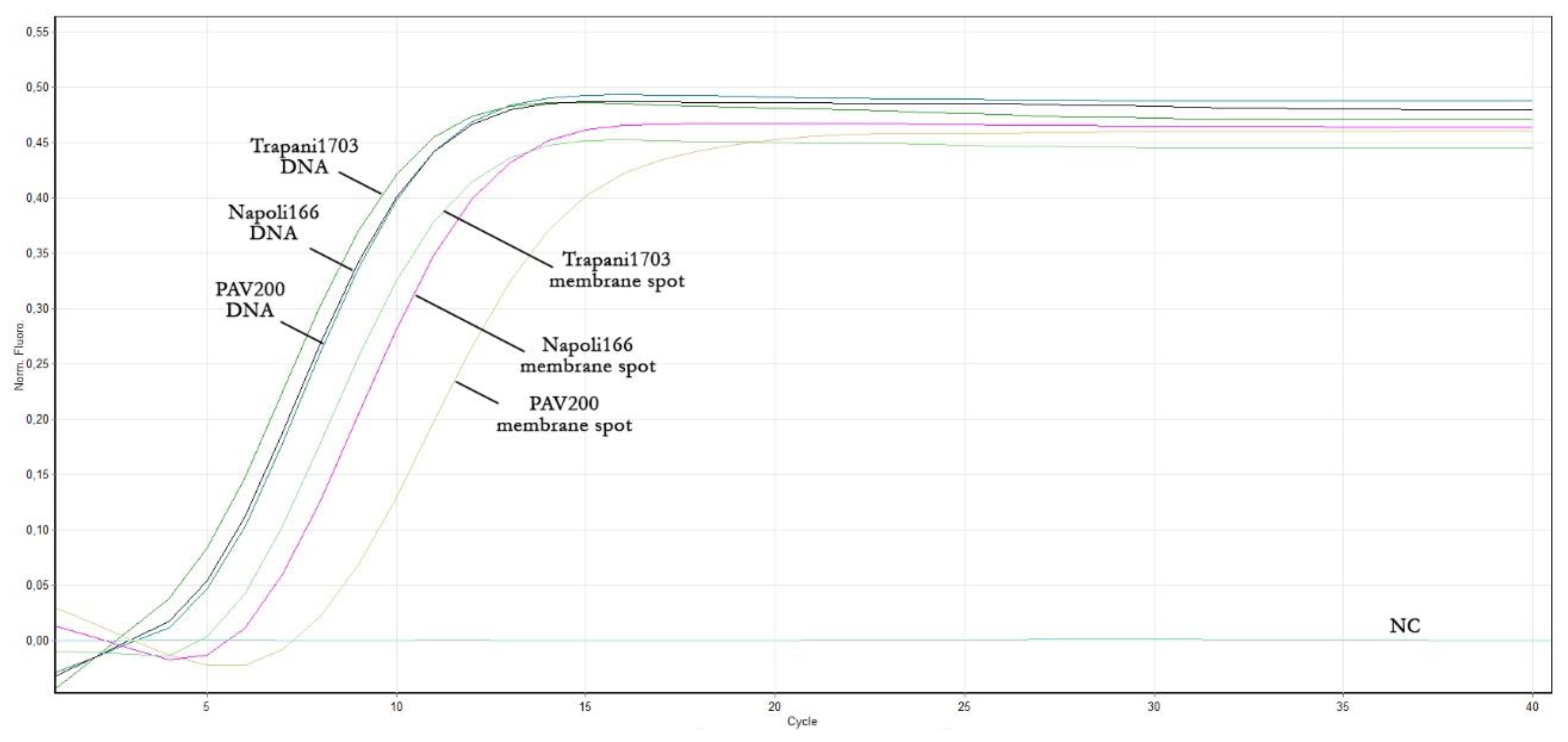
3.5. Rapid Sample Preparation Method Suitable for the ToLCNDV Real-Time LAMP Assay
3.6. Direct In-Field Analysis of Tomato Leaf Curl New Delhi Virus
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- King, A.M.; Lefkowitz, E.; Adams, M.J.; Carstens, E.B. Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier: Berkeley, CA, USA, 2011. [Google Scholar]
- Panno, S.; Caruso, A.G.; Troiano, E.; Luigi, M.; Manglli, A.; Vatrano, T.; Iacono, G.; Marchione, S.; Bertin, S.; Tomassoli, L.; et al. Emergence of tomato leaf curl New Delhi virus in Italy: Estimation of incidence and genetic diversity. Plant Pathol. 2019, 68, 601–608. [Google Scholar] [CrossRef]
- EPPO—European and Mediterranean Plant Protection Organization. Tomato Leaf Curl New Delhi Virus Host Plant Lists. 2022. Available online: https://gd.eppo.int/taxon/TOLCND/hosts (accessed on 10 December 2022).
- Istat—Istituto Nazionale di Statistica. 2021. Available online: http://dati.istat.it (accessed on 10 December 2022).
- Padidam, M.; Beachy, R.N.; Fauquet, C.M. Tomato leaf curl geminivirus from India has a bipartite genome and coat protein is not essential for infectivity. J. Gen. Virol. 1995, 76, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Luigi, M.; Bertin, S.; Manglli, A.; Troiano, E.; Davino, S.; Tomassoli, L.; Parrella, G. First report of tomato leaf curl New Delhi virus causing yellow leaf curl of pepper in Europe. Plant Dis. 2019, 103, 2970. [Google Scholar] [CrossRef]
- Kil, E.J.; Vo, T.T.B.; Fadhila, C.; Ho, P.T.; Lal, A.; Troiano, E.; Parrella, G.; Lee, S. Seed Transmission of Tomato Leaf Curl New Delhi Virus from Zucchini Squash in Italy. Plants 2020, 9, 563. [Google Scholar] [CrossRef] [PubMed]
- Juárez, M.; Tovar, R.; Fiallo-Olivé, E.; Aranda, M.A.; Gosálvez, B.; Castillo, P.; Moriones, E.; Navas-Castillo, J. First detection of Tomato leaf curl New Delhi virus infecting zucchini in Spain. Plant Dis. 2014, 98, 857. [Google Scholar] [CrossRef] [PubMed]
- Panno, S.; Iacono, G.; Davino, M.; Marchione, S.; Zappardo, V.; Bella, P.; Tomassoli, L.; Accotto, G.P.; Davino, S. First report of Tomato leaf curl New Delhi virus affecting zucchini squash in an important horticultural area of southern Italy. New Dis. Rep. 2016, 33, 2044-0588. [Google Scholar] [CrossRef] [Green Version]
- Mnari-Hattab, M.; Zammouri, S.; Belkadhi, M.S.; Bellon Doña, D.; Ben Nahia, E.; Hajlaoui, M.R. First report of Tomato leaf curl New Delhi virus infecting cucurbits in Tunisia. New Dis. Rep. 2015, 31, 21. [Google Scholar] [CrossRef] [Green Version]
- Radouane, N.; Tahiri, A.; El Ghadraoui, L.; Al Figuigui, J.; Lahlali, R. First report of Tomato leaf curl New Delhi virus in Morocco. New Dis. Rep. 2018, 37, 2044-0588. [Google Scholar] [CrossRef] [Green Version]
- Kheireddine, A.; Sifres, A.; Sáez, C.; Picó, B.; López, C. First Report of Tomato Leaf Curl New Delhi Virus Infecting Cucurbit Plants in Algeria. Plant Dis. 2019, 103, 3291. [Google Scholar] [CrossRef]
- Orfanidou, C.G.; Malandraki, I.; Beris, D.; Kektsidou, O.; Vassilakos, N.; Varveri, C.; Katis, N.I.; Maliogka, V.I. First report of tomato leaf curl New Delhi virus in zucchini crops in Greece. J. Plant Pathol. 2019, 101, 799. [Google Scholar] [CrossRef] [Green Version]
- Desbiez, C.; Gentit, P.; Cousseau-Suhard, P.; Renaudin, I.; Verdin, E. First report of Tomato leaf curl New Delhi virus infecting courgette in France. New Dis. Rep. 2021, 42, e12006. [Google Scholar] [CrossRef]
- Luigi, M.; Manglli, A.; Valdes, M.; Sitzia, M.; Davino, S.; Tomassoli, L. Occurrence of Tomato leaf curl New Delhi virus infecting zucchini in Sardinia (Italy). J. Plant Pathol. 2016, 98, 695. [Google Scholar]
- Bertin, S.; Luigi, M.; Parella, G.; Giorgini, M.; Davino, S.; Tomassoli, L. Survey of the distribution of Bemisia tabaci (Hemiptera: Aleyrodidae) in Lazio region (Central Italy): A threat for the northward expansion of Tomato leaf curl New Delhi virus (Begomovirus: Geminiviridae) infection. Phytoparasitica 2018, 46, 171–182. [Google Scholar] [CrossRef]
- Trisciuzzi, N.; Silletti, M.R.; Gallitelli, D.; Spanò, R. First detection of tomato leaf curl New Delhi virus in melon and zucchini squash in southern Italy. J. Plant Pathol. 2018, 100, 149. [Google Scholar] [CrossRef] [Green Version]
- EPPO—European and Mediterranean Plant Protection Organization. 2022. Available online: https://www.eppo.int (accessed on 10 December 2022).
- EUR-Lex—Access to European Union Law. 2022. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32019R2072&from=it (accessed on 10 December 2022).
- Rogers, S.G.; Bisaro, D.M.; Horsch, R.B.; Fraley, R.T.; Hoffmann, N.L.; Brand, L.; Scott Elmer, J. Tomato golden mosaic virus A component DNA replicates autonomously in transgenic plants. Cell 1986, 45, 593–600. [Google Scholar] [CrossRef]
- Sanderfoot, A.A.; Lazarowitz, S.G. Getting it together in plant virus movement: Cooperative interactions between bipartite geminivirus movement proteins. Trends Cell Biol. 1996, 6, 353–358. [Google Scholar] [CrossRef]
- Ruiz, M.L.; Simón, A.; Velasco, L.; García, M.C.; Janssen, D. First report of Tomato leaf curl New Delhi virus infecting tomato in Spain. Plant Dis. 2015, 99, 894. [Google Scholar] [CrossRef]
- Sharma, N.; Prasad, M. An insight into plant–Tomato leaf curl New Delhi virus interaction. Nucleus 2017, 60, 335–348. [Google Scholar] [CrossRef]
- Rosen, R.; Kanakala, S.; Kliot, A.; Pakkianathan, B.C.; Farich, B.A.; Santana-Magal, N.; Elimelech, M.; Kontsedalov, S.; Lebedev, G.; Cilia, M.; et al. Persistent, circulative transmission of begomoviruses by whitefly vectors. Curr. Opin. Virol. 2015, 15, 1–8. [Google Scholar] [CrossRef]
- López, C.; Ferriol, M.; Picó, M.B. Mechanical transmission of Tomato leaf curl New Delhi virus to cucurbit germplasm: Selection of tolerance sources in Cucumis melo. Euphytica 2015, 204, 679–691. [Google Scholar] [CrossRef]
- Hanssen, I.M.; Lapidot, M.; Thomma, B.P. Emerging viral diseases of tomato crops. Mol. Plant-Microbe Interact. 2010, 23, 539–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapisarda, C.; Cascone, G.; Colombo, A.; Tropea, G.; Mazzarella, R.; Serges, T. Physical control possibilities of Bemisia tabaci (Gennadius) and Tomato yellow leaf curl disease (TYLCD) in protected crops. Colt. Protette 2005, 34, 53–64. [Google Scholar]
- Figàs, M.R.; Alfaro-Fernández, A.; Font, M.I.; Borràs, D.; Casanova, C.; Hurtado, M.; Plazas, M.; Prohens, J.; Soler, S. Inoculation of cucumber, melon and zucchini varieties with Tomato leaf curl New Delhi virus and evaluation of infection using different detection methods. Ann. Appl. Biol. 2017, 170, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Jeevalatha, A.; Singh, B.P.; Kaundal, P.; Kumar, R.; Raigond, B. RCA-PCR: A robust technique for the detection of Tomato leaf curl New Delhi virus-[potato] at ultra-low virus titre. Potato J. 2014, 41, 76–80. [Google Scholar]
- Luigi, M.; Manglli, A.; Bertin, S.; Donati, L.; Tomassoli, L.; Ferretti, L.; Faggioli, F. Development and validation of a specific real-time PCR protocol for the detection of tomato leaf curl New Delhi virus. Eur. J. Plant Pathol. 2020, 157, 969–974. [Google Scholar] [CrossRef]
- Alfaro-Fernández, A.; Sánchez-Navarro, J.A.; Landeira, M.; Font, M.I.; Hernández-Llópis, D.; Pallás, V. Evaluation of PCR and non-radioactive molecular hybridization techniques for the routine diagnosis of Tomato leaf curl New Delhi virus, Tomato yellow leaf curl virus and Tomato yellow leaf curl Sardinia virus. J. Plant Pathol. 2016, 98, 245–254. [Google Scholar]
- Naganur, P.; Premchand, U.; Shankarappa, K.S.; Mesta, R.K.; Manjunatha, C.; Patil, C.V. Development of a loop-mediated isothermal amplification assay for detection of Tomato leaf curl New Delhi virus in ridge gourd [Luffa acutangula (L.) Roxb.]. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 2282–2295. [Google Scholar] [CrossRef]
- Mahas, A.; Hassan, N.; Aman, R.; Marsic, T.; Wang, Q.; Ali, Z.; Mahfouz, M.M. LAMP-coupled CRISPR–Cas12a module for rapid and sensitive detection of plant DNA viruses. Viruses 2021, 13, 466. [Google Scholar] [CrossRef]
- Davino, S.; Panno, S.; Iacono, G.; Sabatino, L.; D’Anna, F.; Iapichino, G.; Olmos, A.; Scuderi, G.; Rubio, L.; Tomassoli, L.; et al. Genetic variation and evolutionary analysis of Pepino mosaic virus in Sicily: Insights into the dispersion and epidemiology. Plant Pathol. 2017, 66, 368–375. [Google Scholar] [CrossRef] [Green Version]
- Davino, S.; Panno, S.; Rangel, E.A.; Davino, M.; Bellardi, M.G.; Rubio, L. Population genetics of cucumber mosaic virus infecting medicinal, aromatic and ornamental plants from northern Italy. Arch. Virol. 2012, 157, 739–745. [Google Scholar] [CrossRef]
- Panno, S.; Matić, S.; Tiberini, A.; Caruso, A.G.; Bella, P.; Torta, L.; Stassi, R.; Davino, S. Loop mediated isothermal amplification: Principles and applications in plant virology. Plants 2020, 9, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagamine, K.; Hase, T.; Notomi, T. Accelerated reaction by loop-mediated isothermal amplification using loop primers. Mol. Cell Probes 2002, 16, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Bertacca, S.; Caruso, A.G.; Trippa, D.; Marchese, A.; Giovino, A.; Matic, S.; Noris, E.; Font, M.I.; Alfaro, A.; Panno, S.; et al. Development of a Real-Time Loop-Mediated Isothermal Amplification Assay for the Rapid Detection of Olea Europaea Geminivirus. Plants 2022, 11, 660. [Google Scholar] [CrossRef] [PubMed]
- Davino, S.; Panno, S.; Arrigo, M.; La Rocca, M.; Caruso, A.G.; Bosco, G.L. Planthology: An application system for plant diseases management. Chem. Eng. Trans. 2017, 58, 619–624. [Google Scholar] [CrossRef]
- Acosta-Leal, R.; Duffy, S.; Xiong, Z.; Hammond, R.; Elena, S.F. Advances in plant virus evolution: Translating evolutionary insights into better disease management. Phytopathology 2011, 101, 1136–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferriol, I.; Rubio, L.; Pérez-Panadés, J.; Carbonell, E.A.; Davino, S.; Belliure, B. Transmissibility of Broad bean wilt virus 1 by aphids: Influence of virus accumulation in plants, virus genotype and aphid species. Ann. Appl. Biol. 2013, 162, 71–79. [Google Scholar] [CrossRef]
- Panno, S.; Davino, S.; Caruso, A.G.; Bertacca, S.; Crnogorac, A.; Mandić, A.; Noris, E.; Matić, S. A review of the most common and economically important diseases that undermine the cultivation of tomato crop in the Mediterranean basin. Agronomy 2021, 11, 2188. [Google Scholar] [CrossRef]
- Davino, S.; Davino, M.; Accotto, G.P. A single-tube PCR assay for detecting viruses and their recombinants that cause tomato yellow leaf curl disease in the Mediterranean basin. J. Virol. Methods 2008, 147, 93–98. [Google Scholar] [CrossRef]
- Panno, S.; Caruso, A.G.; Barone, S.; Lo Bosco, G.; Rangel, E.A.; Davino, S. Spread of tomato brown rugose fruit virus in Sicily and evaluation of the spatiotemporal dispersion in experimental conditions. Agronomy 2020, 10, 834. [Google Scholar] [CrossRef]
- Roy, B.; Chakraborty, P.; Ghosh, A. How many begomovirus copies are acquired and inoculated by its vector, whitefly (Bemisia tabaci) during feeding? PLoS ONE 2021, 16, e0258933. [Google Scholar] [CrossRef]
- Rodríguez, E.; Téllez, M.M.; Janssen, D. Whitefly control strategies against tomato leaf curl New Delhi virus in greenhouse zucchini. Int. J. Environ. Res. Public Health 2019, 16, 2673. [Google Scholar] [CrossRef] [Green Version]
- Clark, M.F.; Adams, A. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol. 1977, 34, 475. [Google Scholar] [CrossRef] [PubMed]
- Ferriol, I.; Rangel, E.A.; Panno, S.; Davino, S.; Han, C.G.; Olmos, A.; Rubio, L. Rapid detection and discrimination of fabaviruses by flow-through hybridisation with genus-and species-specific riboprobes. Ann. Appl. Biol. 2015, 167, 26–35. [Google Scholar] [CrossRef]
- Panno, S.; Caruso, A.G.; Bertacca, S.; Matić, S.; Davino, S.; Parrella, G. Detection of parietaria mottle virus by RT-qPCR: An emerging virus native of Mediterranean area that undermine tomato and pepper production in Southern Italy. Front. Plant Sci. 2021, 12, 698573. [Google Scholar] [CrossRef]
- Schrader, C.; Schielke, A.; Ellerbroek, L.; Johne, R. PCR inhibitors–occurrence, properties and removal. J. Appl. Microbial. 2012, 113, 1014–1026. [Google Scholar] [CrossRef] [PubMed]
- Boonham, N.; Glover, R.; Tomlinson, J.; Mumford, R. Exploiting generic platform technologies for the detection and identification of plant pathogens. Sustain. Dis. Manag. Eur. Context 2008, 121, 355–363. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Quantity |
|---|---|
| Sample extraction bag with 2 mL extraction buffer | 20 |
| Single hole puncher | 1 |
| 2 mL tubes with ≈0.2 cm2 hybridisation membrane | 20 |
| Hand homogeniser | 1 |
| Glycine buffer | 6 mL |
| LAMP reaction mixture | 550 µL |
| Positive control (PC) | 1 |
| Negative control (NC) | 1 |
| bCube2 thermal cycler | 1 |
| 16-well bCube cartridge | 1 |
| bCube cartridge aluminium foil | 1 |
| 12 Volt rechargeable battery | 1 |
| Smartphone/laptop | 1 |
| 100–1000 µL micropipette | 1 |
| 10–100 µL micropipette | 1 |
| 0.5–10 µL micropipette | 1 |
| 100–1000 µL micropipette tips | 1 rack (96 tips) |
| 10–100 µL micropipette tips | 1 rack (96 tips) |
| 0.5–10 µL micropipette tips | 1 rack (96 tips) |
| Primer Name | Length (nt) | Sequence 5′-3′ | Nucleotide Position (nt) | Amplicon Size (bp) | Reference Sequence (Acc. No.) |
|---|---|---|---|---|---|
| ToLCNDV-F3 | 18 | GTGGCATGCTACTGTGAC | 821–838 | 218 | MK732932 |
| ToLCNDV-B3 | 23 | CCGAATCATAAAAATAGATCCGG | 1016–1038 | ||
| ToLCNDV-BIP | 46 | CGGCAAGTATGAGAATCATACTGAACAAAGTAGCATACACAGGATT | 926–971 | – | |
| ToLCNDV-FIP | 45 | GCCTCTTGTTGATTGTAAACAACATGAGGAACGTATGCATCAAGG | 881–925 | ||
| ToLCNDV-LF | 25 | CAAACTTCCTAACTAATGCTTGCTC | 861–885 | – | |
| ToLCNDV-LB | 24 | TGTTGTATATGGCCTGTACTCATG | 961–984 |
| Sample | Real-Time LAMP Reaction Time (min) | ||
|---|---|---|---|
| Assay #1 | Assay #2 | Assay #3 | |
| ToLCNDV Trapani1703 | 14 | 14.8 | 14.2 |
| ToLCNDV Napoli166 | 16 | 15.4 | 14.9 |
| ToLCNDV PAV200 | 15 | 15.8 | 15.3 |
| Negative control (NC) | - | - | - |
| Starting DNA Concentration (50 ng/μL) (4.41 × 1010 Copies) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Assay | 100 | 10−1 | 10−2 | 10−3 | 10−4 | 10−5 | 10−6 | 10−7 | 10−8 | 10−9 | 10−10 |
| End-point PCR | + | + | + | + | + | + | − | − | − | − | − |
| LAMP reaction time plateau (min) (mean values ± SD) | 10.3 ± 0.2 | 10.3 ± 0.2 | 11 ± 0.2 | 14 ± 0.3 | 16.2 ± 0.3 | 15.9 ± 0.5 | 19.1 ± 0.2 | 24.9 ± 0.1 | 45 ± 0.2 | − | − |
| Sample | Reaction Time Plateau (min) (Mean Values ± SD) | |
|---|---|---|
| Total DNA (Commercial Kit) | Membrane Spot Crude Extract | |
| ToLCNDV Trapani1703 | 13.1 ± 0.3 | 15.2 ± 0.3 |
| ToLCNDV Napoli166 | 13.2 ± 0.3 | 16.2 ± 0.5 |
| ToLCNDV PAV200 | 12.9 ± 0.2 | 20.1 ±0.3 |
| Negative control (NC) | - | - |
| Sample Source | ID Sample | Symptomatic | In-Field Conditions | Laboratory Conditions | |
|---|---|---|---|---|---|
| Real-Time LAMP Results | End Point PCR Results | Real-Time LAMP Results (Total DNA/Membrane Spot Crude Extract) | |||
| Zucchini squash | ZS-01 | + | + | + | +/+ |
| ZS-02 | − | + | − | +/+ | |
| ZS-03 | − | + | − | +/+ | |
| ZS-04 | + | + | + | +/+ | |
| ZS-05 | + | + | + | +/+ | |
| ZS-06 | + | + | + | +/+ | |
| ZS-07 | − | − | − | − | |
| ZS-08 | + | + | + | +/+ | |
| ZS-09 | − | − | − | − | |
| ZS-10 | + | + | + | +/+ | |
| ZS-11 | + | + | + | +/+ | |
| ZS-12 | + | + | + | +/+ | |
| ZS-13 | + | + | + | +/+ | |
| ZS-14 | + | + | + | +/+ | |
| ZS-15 | − | − | − | − | |
| ZS-16 | + | + | + | +/+ | |
| ZS-17 | + | + | + | +/+ | |
| ZS-18 | + | + | + | +/+ | |
| ZS-19 | + | + | + | +/+ | |
| ZS-20 | + | + | + | +/+ | |
| ZS-NC | − | − | − | − | |
| Tomato | Tom-01 | − | + | − | +/+ |
| Tom-02 | − | − | − | − | |
| Tom-03 | + | + | + | +/+ | |
| Tom-04 | − | − | − | − | |
| Tom-05 | + | + | + | +/+ | |
| Tom-06 | + | + | + | +/+ | |
| Tom-07 | − | + | − | +/+ | |
| Tom-08 | + | + | + | +/+ | |
| Tom-09 | + | + | + | +/+ | |
| Tom-10 | − | + | − | +/+ | |
| Tom-11 | + | + | + | +/+ | |
| Tom-12 | + | + | + | +/+ | |
| Tom-13 | + | + | + | +/+ | |
| Tom-14 | + | + | + | +/+ | |
| Tom-15 | + | + | + | +/+ | |
| Tom-16 | − | − | − | − | |
| Tom-17 | − | − | − | − | |
| Tom-18 | − | + | − | +/+ | |
| Tom-19 | + | + | + | +/+ | |
| Tom-20 | + | + | + | +/+ | |
| Tom-NC | − | − | − | − | |
| Pepper | Pep-01 | + | + | + | +/+ |
| Pep-02 | + | + | + | +/+ | |
| Pep-03 | + | + | + | +/+ | |
| Pep-04 | + | + | + | +/+ | |
| Pep-05 | − | + | − | +/+ | |
| Pep-06 | + | + | + | +/+ | |
| Pep-07 | + | + | + | +/+ | |
| Pep-08 | + | + | + | +/+ | |
| Pep-09 | − | − | − | − | |
| Pep-10 | − | − | − | − | |
| Pep-11 | − | − | − | − | |
| Pep-12 | + | + | + | +/+ | |
| Pep-13 | + | + | + | +/+ | |
| Pep-14 | − | + | + | +/+ | |
| Pep-15 | − | − | − | − | |
| Pep-16 | + | + | + | +/+ | |
| Pep-17 | − | − | − | − | |
| Pep-18 | + | + | + | +/+ | |
| Pep-19 | + | + | + | +/+ | |
| Pep-20 | − | − | − | − | |
| Pep-NC | − | − | − | − | |
| ToLCNDV-positive control | PC | / | + | + | +/+ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caruso, A.G.; Ragona, A.; Bertacca, S.; Montoya, M.A.M.; Panno, S.; Davino, S. Development of an In-Field Real-Time LAMP Assay for Rapid Detection of Tomato Leaf Curl New Delhi Virus. Plants 2023, 12, 1487. https://doi.org/10.3390/plants12071487
Caruso AG, Ragona A, Bertacca S, Montoya MAM, Panno S, Davino S. Development of an In-Field Real-Time LAMP Assay for Rapid Detection of Tomato Leaf Curl New Delhi Virus. Plants. 2023; 12(7):1487. https://doi.org/10.3390/plants12071487
Chicago/Turabian StyleCaruso, Andrea Giovanni, Arianna Ragona, Sofia Bertacca, Mauricio Alejandro Marin Montoya, Stefano Panno, and Salvatore Davino. 2023. "Development of an In-Field Real-Time LAMP Assay for Rapid Detection of Tomato Leaf Curl New Delhi Virus" Plants 12, no. 7: 1487. https://doi.org/10.3390/plants12071487
APA StyleCaruso, A. G., Ragona, A., Bertacca, S., Montoya, M. A. M., Panno, S., & Davino, S. (2023). Development of an In-Field Real-Time LAMP Assay for Rapid Detection of Tomato Leaf Curl New Delhi Virus. Plants, 12(7), 1487. https://doi.org/10.3390/plants12071487