
Yellow Leaf Disease Resistance and Melanaphis sacchari Preference in Commercial Sugarcane Cultivars

 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. YLD Phenotyping
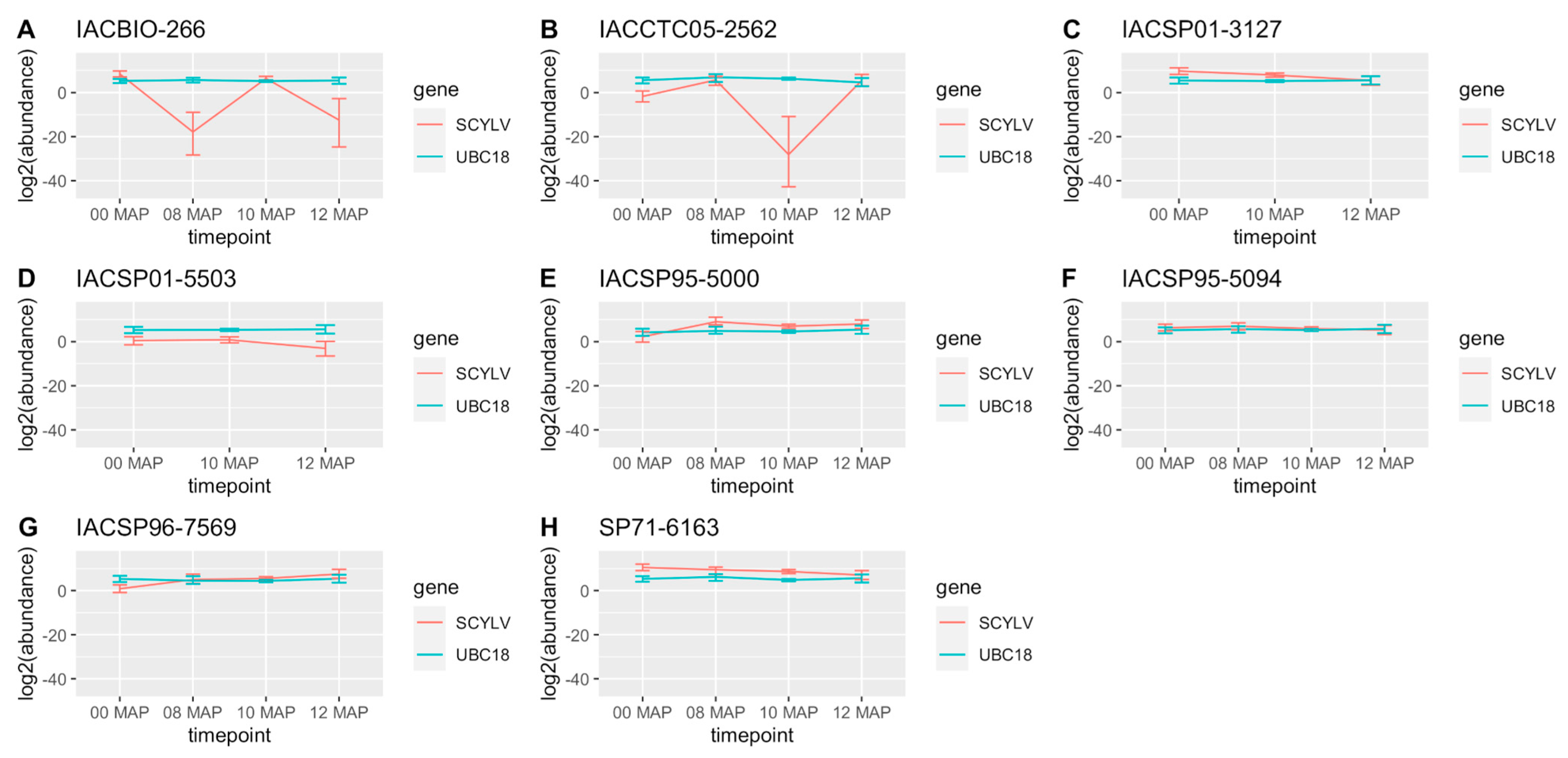
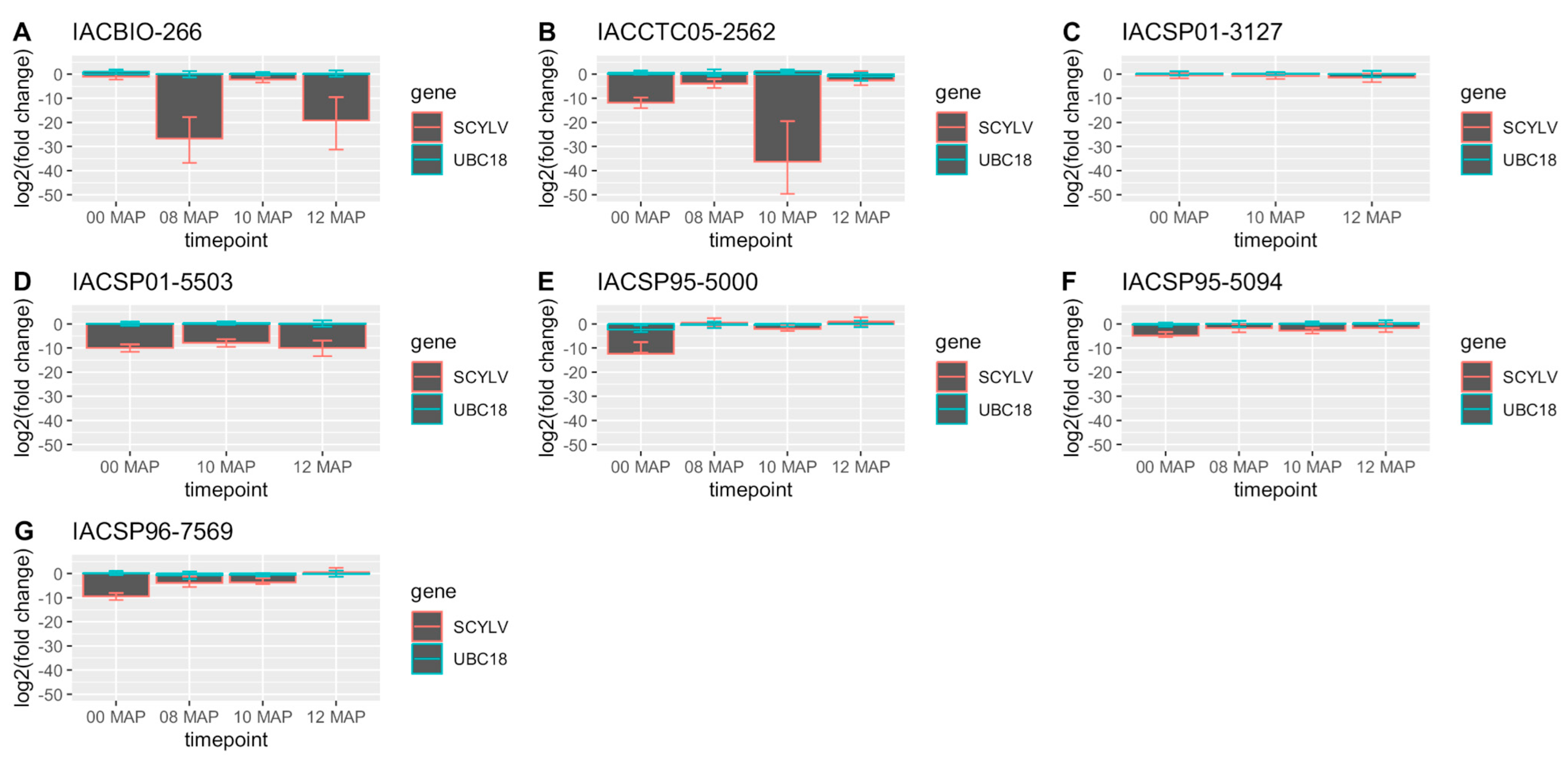
2.2. RT-qPCR and Virus Titer
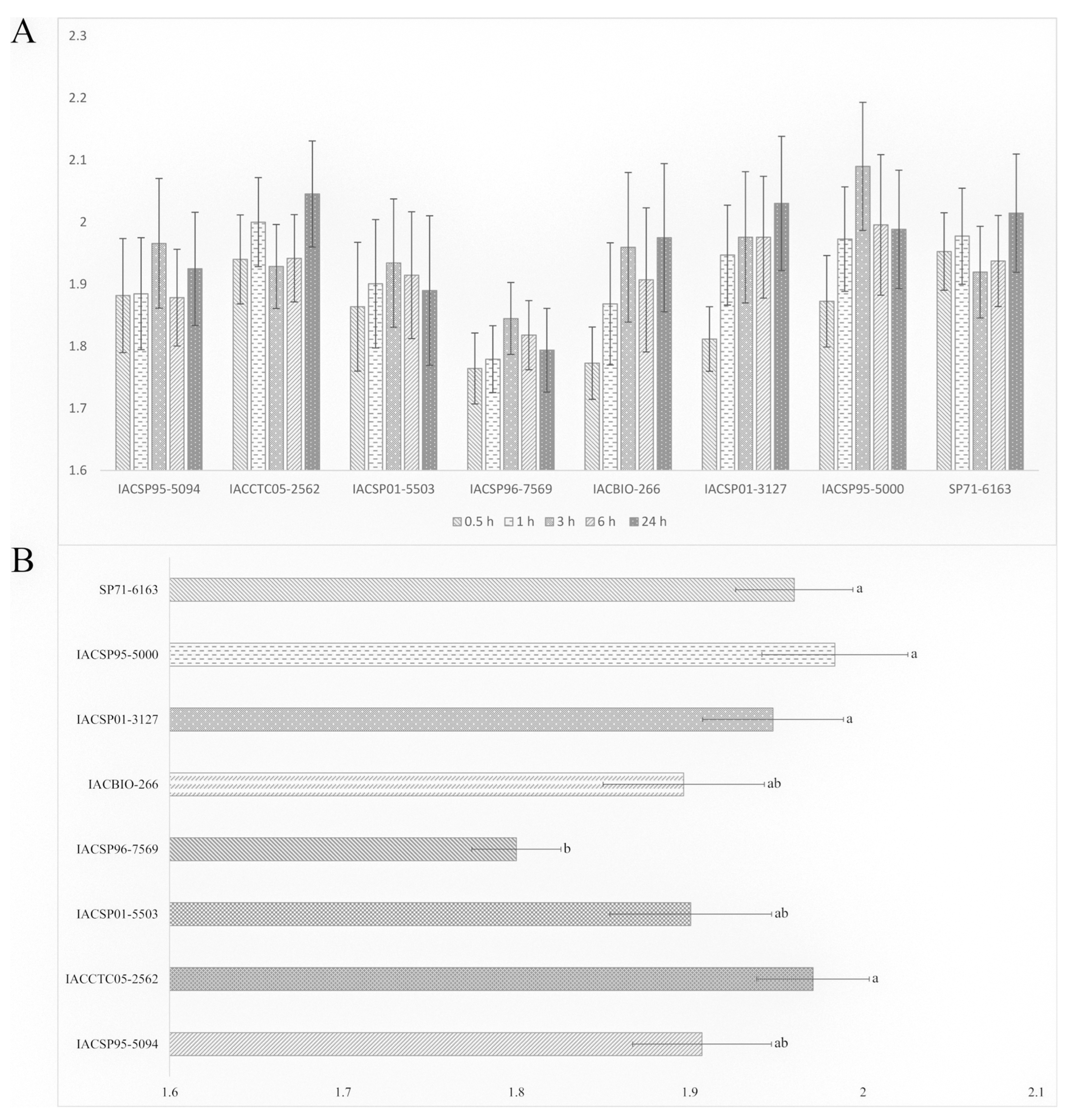
2.3. Aphid Preference
3. Discussion
4. Material and Methods
4.1. Plant Material
4.2. Aphid Rearing
4.3. YLD Phenotyping
4.4. RNA Extraction and RT-qPCR
4.5. Aphid Settling Behavior
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Waclawovsky, A.J.; Sato, P.M.; Lembke, C.G.; Moore, P.H.; Souza, G.M. Sugarcane for bioenergy production: An assessment of yield and regulation of sucrose content. Plant Biotechnol. J. 2010, 8, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Jackson, P.; Wei, X.; Ross, E.M.; Aitken, K.; Deomano, E.; Atkin, F.; Hayes, B.J.; Voss-Fels, K.P. Accelerating Genetic Gain in Sugarcane Breeding Using Genomic Selection. Agronomy 2020, 10, 585. [Google Scholar] [CrossRef]
- Carpio, L.G.; Souza, F.S. Optimal allocation of sugarcane bagasse for producing bioelectricity and second generation ethanol in Brazil: Scenarios of cost reductions. Renew. Energy 2017, 111, 771–780. [Google Scholar] [CrossRef]
- Reis, C.E.R.; Hu, B. Vinasse from sugarcane Ethanol production: Better treatment or better utilization? Front. Energy Res. 2017, 5, 7. [Google Scholar]
- Cheavegatti-Gianotto, A.; Abreu, H.M.C.; Arruda, P.; Filho, J.C.B.; Burnquist, W.L.; Creste, S.; Ciero, L.; Ferro, J.A.; Figueira, A.V.O.; Filgueiras, T.S.; et al. Sugarcane (Saccharum x officinarum): A reference study for the regulation of genetically modified cultivars in Brazil. Trop. Plant Biol. 2011, 4, 62–89. [Google Scholar] [CrossRef]
- Gonçalves, M.C.; Pinto, L.R.; Souza, S.C.; Landell, M.G.A. Virus Diseases of Sugarcane. A Constant Challenge to Sugarcane Breeding in Brazil. Funct. Plant Sci. Biotechnol. 2012, 6, 108–116. [Google Scholar]
- ElSayed, A.I.; Komor, E.; Boulila, M.; Viswanathan, R.; Odero, D.C. Biology and management of sugarcane yellow leaf vírus: An historical overview. Arch. Virol. 2015, 160, 2921–2934. [Google Scholar] [CrossRef]
- Holkar, S.K.; Balasubramaniam, P.; Kumar, A.; Kadirvel, N.; Shingote, P.R.; Chhabra, M.L.; Kumar, S.; Kumar, P.; Viswanathan, R.; Jain, R.K.; et al. Present status and future management strategies for sugarcane yellow leaf virus: A major constraint to the global sugarcane production. Plant Pathol. J. 2020, 36, 536–557. [Google Scholar] [CrossRef]
- Grisham, M.P.; Eggleston, G.; Hoy, J.W.; Viator, R.P. The effect of Sugarcane yellow leaf virus infection on yield of sugarcane in Louisiana. Sugar Cane Int. 2009, 27, 3–91. [Google Scholar]
- Lehrer, A.T.; Wu, K.K.; Komor, E. Impact of sugarcane yellow leaf virus (ScYLV) on growth and sugar yield of sugarcane. J. Gen. Plant Pathol. 2009, 75, 288–296. [Google Scholar] [CrossRef]
- Izaguirre-Mayoral, M.L.; Carballo, O.; Alceste, C.; Romano, M.; Nass, H.A. Physiological performance of asymptomatic and yellow leaf syndrome-affected sugarcanes in Venezuela. J. Phytopathol. 2002, 150, 13–19. [Google Scholar] [CrossRef]
- Boukari, W.; Kaye, C.; Wei, C.; Hincapie, M.; LaBorde, C.; Irey, M.; Rott, P. Field infection of virus-free sugarcane by Sugarcane yellow leaf virus and effect of yellow leaf on sugarcane grown on organic and on mineral soils in Florida. Plant Dis. 2019, 103, 2367–2373. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, R.; Balamuralikrishnan, M.; Karuppaiah, R. Yellow leaf disease of sugarcane: Occurrence and impact of infected sets on disease severity and yield. Proc. Sugar Cancer Technol. Assoc. India 2006, 67, 74–89. [Google Scholar]
- Gray, S.; Cilia, M.; Ghanim, M. Circulative, “nonpropagative” virus transmission: An orchestra of virus, insect, and plant-derived instruments. Adv. Virus Res. 2014, 89, 141–199. [Google Scholar] [PubMed]
- Rott, P.; Mirkov, T.E.; Schenck, S.; Girard, J.C. Recent advances in research on sugarcane yellow leaf vírus, the causal agents of sugarcane yellow leaf. Sugar Cane Int. 2008, 26, 18–22. [Google Scholar]
- Ahmad, A.Y.; Costet, L.; Daugrois, J.H.; Nibouche, S.; Letourmy, P.; Girard, J.C.; Rott, P. Variation in infection capacity and in virulence exists between genotypes of Sugarcane yellow leaf virus. Plant Dis. 2007, 91, 253–259. [Google Scholar] [CrossRef]
- Lehrer, A.T.; Schenck, S.; Yan, S.-L.; Komor, E. Movement of aphid-transmitted Sugarcane yellow leaf vírus (ScYLV) within and between sugarcane plants. Plant Pathol. 2007, 56, 711–717. [Google Scholar] [CrossRef]
- Chinnaraja, C.; Viswanathan, R. Quantification of sugarcane yellow leaf virus in sugarcane following transmission through aphid vector Melanaphis sacchari. Virus Dis. 2015, 26, 237–242. [Google Scholar] [CrossRef]
- Gonçalves, M.C.; Vega, J.; Oliveira, J.G.; Gomes, M.M.A. Sugarcane yellow leaf virus infection leads to alterations in photosynthetic efficiency and carbohydrate accumulation in sugarcane leaves. Fitop. Brasil. 2005, 30, 10–16. [Google Scholar] [CrossRef]
- Lehrer, A.T.; Komor, E. Symptom expression of yellow leaf disease in sugarcane cultivars with different degrees of infection by Sugarcane yellow leaf vírus. Plant Pathol. 2008, 57, 178–189. [Google Scholar] [CrossRef]
- Viswanathan, R. Impact of yellow leaf disease in sugarcane and its successful disease management to sustain crop production. Indian Phytopathol. 2021, 74, 573–586. [Google Scholar] [CrossRef]
- Rassaby, L.; Girard, J.C.; Letourmy, P.; Chaume, J.; Irey, M.S.; Lockhart, B.E.; Kodja, H.; Rott, P. Impact of sugarcane yellow leaf virus on sugarcane yield and juice quality in Reunion Island. Eur. J. Plant Pathol. 2003, 109, 459–466. [Google Scholar] [CrossRef]
- Bertani, R.P.; Perera, M.F.; Arias, M.E.; Luque, C.; Funes, C.; González, V.; Cuenya, M.I.; Ploper, L.D.; Welin, B.; Castagnaro, A.P. A study of the sugarcane yellow leaf disease in Argentina. Plant Dis. J. 2014, 98, 1036–1042. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, R.; Karuppaiah, R.; Malathi, P.; Ganesh Kumar, V.; Chinnaraja, C. Diagnosis of sugarcane yellow leaf virus in asymptomatic sugarcane by RT-PCR. Sugar Tech. 2009, 11, 368–372. [Google Scholar] [CrossRef]
- Comstock, J.C.; Irey, M.S.; Lockhart, B.E.L.; Wang, Z.K. Incidence of yellow leaf syndrome in CP cultivars based on polymerase chain reaction and serological techniques. Sugar Cane 1998, 4, 21–24. [Google Scholar]
- Gao, S.J.; Lin, Y.H.; Pan, Y.B.; Damaj, M.B.; Wang, Q.N.; Mirkov, T.E.; Chen, R.K. Molecular characterization and phylogenetic analysis of Sugarcane yellow leaf virus isolates from China. Virus Genes 2012, 45, 340–349. [Google Scholar] [CrossRef]
- Grisham, M.P.; Comstock, J.C.; Flynn, J.L. Comparing the natural spread of Sugarcane yellow leaf virus in Florida to its natural spread in Louisiana. J. Am. Soc. Sugar Cane Technol. 2010, 30, 151. [Google Scholar]
- Zhu, Y.J.; Lim, S.T.; Schenck, S.; Arcinas, A.; Komor, E. RT-PCR and quantitative real-time RT-PCR detection of Sugarcane yellow leaf virus (SCYLV) in symptomatic and asymptomatic plants of Hawaiian sugarcane cultivars and the correlation of SCYLV titre to yield. Eur. J. Plant Pathol. 2010, 127, 263–273. [Google Scholar] [CrossRef]
- ElSayed, A.I.; Boulila, M.; Rott, P. Molecular evolutionary history of Sugarcane yellow leaf vírus based on sequence analysis of RNA-dependent RNA polymerase and putative aphid transmission factor-coding genes. J. Mol. Evol. 2014, 78, 349–365. [Google Scholar] [CrossRef]
- Gonçalves, M.C.; Klerks, M.M.; Verbeek, M.; Veja, J.; Van den Heuvel, J.F.J.M. The use of molecular beacons combined with NASBA for the sensitive detection of sugarcane yellow leaf virus. Eur. J. Plant Pathol. 2002, 108, 401–407. [Google Scholar] [CrossRef]
- Amata, R.L.; Fernandez, E.; Filloux, D.; Martin, D.P.; Rott, P.; Roumagnac, P. Prevalence of Sugarcane yellow leaf virus in sugarcane producing regions in Kenya revealed by reverse transcription loop-mediated isothermal amplification method. Plant Dis. 2015, 100, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Scagliusi, S.M.; Lockhart, B.E.L. Transmission, characterization, and serology of a luteovirus associated with yellow leaf syndrome of sugarcane. Phytopathology 2000, 90, 120–124. [Google Scholar] [CrossRef]
- Chatenet, M.; Delage, C.; Ripolles, M.; Irey, M.S.; Lockhart, B.E.L.; Rott, P. Detection of Sugarcane yellow leaf vírus in quarantine and production of virus-free sugarcane by apical meristem culture. Plant Dis. 2001, 85, 1177–1180. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, R.; Balamuralikrishnan, M. Detection of sugarcane yellow leaf virus, the causal agent of yellow leaf syndrome in sugarcane by DAS-ELISA Arch. Phytopathol. Plant Protect 2004, 37, 169–176. [Google Scholar] [CrossRef]
- Schenck, S.; Hu, J.S.; Lockhart, B.E. Use of a tissue blot immunoassay to determine the distribution of sugarcane yellow leaf virus in Hawaii. Sugar Cane 1997, 4, 5–8. [Google Scholar]
- Parakh, D.R.; Shamloul, A.M.; Hadidi, A.; Waterworth, H.E.; Scott, S.W.; Howell, H.E.; Mink, G.I. Detection of Prune dwarf ilarvirus from infected stone fruits using reverse transcription polymerase chain reaction. Acta Hortic. 1995, 386, 421–430. [Google Scholar] [CrossRef]
- Nassuth, A.; Pollari, E.; Helmeczy, K.; Stewart, S.; Ko-falvi, A. Improved RNA extraction and one-tube RT-PCR assay for simultaneous detection of control plantRNA plus several viruses in plant extracts. J. Virol. Methods 2000, 90, 37–49. [Google Scholar] [CrossRef]
- Burbano, R.C.; Gonçalves, M.C.; Nobile, P.M.; dos Anjos, I.A.; da Silva, M.F.; Perecin, D.; Anjos, L.O.S.; Fernandes, V.B.V.R.; Pinto, L.R. Screening of Saccharum spp. genotypes for sugarcane yellow leaf virus resistance by combining symptom phenotyping and highly precise virus titration. Crop Prot. 2021, 144, 105577. [Google Scholar] [CrossRef]
- Chinnaraja, C.; Viswanathan, R.; Sathyabhama, M.; Parameswari, B.; Bagyalakshmi, K.; Malathi, P.; Neelamathi, D. Quantification of sugarcane yellow leaf virus in in vitro plantlets and asymptomatic plants of sugarcane by RT-qPCR. Curr. Sci. 2014, 106, 729–734. [Google Scholar]
- Carr, J.P.; Murphy, A.M.; Tungadi, T.; Yoon, J.Y. Plant defense signals: Players and pawns in plant-virus-vector interactions. Plant Sci. 2019, 279, 87–95. [Google Scholar] [CrossRef]
- Ray, S.; Casteel, C.L. Effector-mediated plant–virus–vector interactions. The Plant Cell 2022, 34, 1514–1531. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Mescher, M.C.; Wang, S.; Chen, G.; Xie, W.; Wu, Q.; Wang, W.; Zhang, Y. Tomato yellow leaf curl virus differentially influences plant defense responses to a vector and a non-vector herbivore. Plant Cell Environ. 2016, 39, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Ziebell, H.; Murphy, A.M.; Groen, S.C.; Tungadi, T.; Westwood, J.H.; Lewsey, M.G.; Moulin, M.; Kleczkowski, A.; Smith, A.G.; Stevens, M.; et al. Cucumber mosaic virus and its 2b RNA silencing suppressor modify plant aphid interactions in Tobacco. Sci. Rep. 2011, 1, 187. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Weldegergis, B.T.; Li, J.; Jung, C.; Qu, J.; Sun, Y.; Qian, H.; Tee, C.; Van Loon, J.J.; Dicke, M.; et al. Virulence factors of geminivirus interact with MYC2 to subvert plant resistance and promote vector performance. Plant Cell 2014, 26, 4991–5008. [Google Scholar] [CrossRef]
- Casteel, C.L.; Falk, B.W. Plant virus-vector interactions: More than just for virus transmission. In Current Research Topics in Plant Virology; Wang, A., Zhou, X., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 217–240. [Google Scholar]
- Mauck, K.E.; Chesnais, Q.; Shapiro, L.R. Evolutionary determinants of host and vector manipulation by plant viruses. In Virology and Virus Ecology; Malmstrom, C., Ed.; Environmental Academic Press: Cambridge, MA, USA, 2018; pp. 189–250. [Google Scholar]
- Patton, M.F.; Bak, A.; Sayre, J.M.; Heck, M.L.; Casteel, C.L. A polerovirus, Potato leafroll virus, alters plant–vector interactions using three viral proteins. Plant Cell Environ. 2020, 43, 387–399. [Google Scholar] [CrossRef]
- Madden, L.V.; Jeger, M.J.; van den Bosch, F. A theoretical assessment of the effects of vector-virus transmission mechanism on plant virus disease epidemics. Phytopathology 2000, 90, 576–594. [Google Scholar] [CrossRef]
- Fortes, I.M.; Muñoz, R.F.; Moriones, E. Host plant resistance to Bemisia tabaci to control damage caused in tomato plants by the emerging crinivirus tomato chlorosis vírus. Front. Plant. Sci. 2020, 11, 585510. [Google Scholar] [CrossRef]
- Painter, R.H. Insect Resistance in Crop Plants; The Macmillan Company: New York, NY, USA, 1951; Volume 72, p. 481. [Google Scholar]
- Fartek, B.; Nibouche, S.; Turpin, P.; Costet, B.R. Resistance to Melanaphis sacchari in the sugarcane cultivar R 365. Entomol. Exp. Appl. 2012, 144, 270–278. [Google Scholar] [CrossRef]
- Van Emden, H.F.; Harrington, R. Aphids as Crop Pests; Cabi: Wallingford, UK, 2007. [Google Scholar]
- Williams, W.; Kennedy, G.G.; Yamamoto, R.T.; Thacker, J.; Bordner, J. 2-Tridecanone: A naturally occurring insecticide from the wild tomato Lycopersicon hirsutum f. glabratum. Science 1980, 207, 888–889. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.M. Plant Resistance to Arthropods: Molecular and Conventional Approaches; Springer Science & Business Media: Berlin, Germany, 2005. [Google Scholar]
- Fartek, B.; Nibouche, S.; Costet, L.; Reynaud, B. Characterization of Sugarcane Resistance to Melanaphis sacchari Using Biological Tests and Electrical Penetration Graph (DC-EPG); Symposium on Plant Interactions with Aphids: Wageningen, The Netherlands, 2008; p. 1. [Google Scholar]
- Akbar, W.; Showler, A.T.; Reagan, T.E.; Davis, J.A.; Beuzelin, J.M. Feeding by sugarcane aphid, Melanaphis sacchari, on sugarcane cultivars with differential susceptibility and potential mechanism of resistance. Entomol. Exp. Appl. 2014, 150, 32–44. [Google Scholar] [CrossRef]
- Bertasello, L.E.T.; Sousa, M.C.; Maluta, N.K.P.; Pinto, L.R.; Lopes, J.R.S.; Gonçalves, M.C. Effect of sugarcane cultivars infected with sugarcane yellow leaf vírus (ScYLV) on feeding behavior and biological performance of Melanaphis sacchari (Hemiptera: Aphididae). Plants 2021, 10, 2122. [Google Scholar] [CrossRef]
- Ramakers, C.; Ruijter, J.M.; Deprez, R.H.L.; Moorman, A.F.M. Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci. Lett. 2003, 339, 62–66. [Google Scholar] [CrossRef]
- Gonçalves, M.C. Doenças causadas por vírus. In Cana-de-Açúcar; Dinardo-Miranda, L.L., Vasconcelos, A.C.M., Landell, M.G.A., Eds.; Instituto Agronômico de Campinas: São Paulo, Brasil, 2008; Volume 1, pp. 150–165. [Google Scholar]
- Zhu, Y.J.; McCafferty, H.; Osterman, G.; Lim, S.; Agbayani, R.; Lehrer, A.; Schenck, S.; Komor, E. Genetic transformation with untranslatable coat protein gene of sugarcane yellow leaf virus reduces virus titers in sugarcane. Transgenic Res. 2011, 20, 503–512. [Google Scholar] [CrossRef]
- Bagyalakshmi, K.; Viswanathan, R.; Ravichandran, V. Impact of the viruses associated with mosaic and yellow leaf disease on varietal degeneration in sugarcane. Phytoparasitica 2019, 47, 591–604. [Google Scholar] [CrossRef]
- Pimenta, R.J.G.; Aono, A.H.; Burbano, R.C.V.; Coutinho, A.E.; Silva, C.C.; Dos Anjos, I.A.; Perecin, D.; Landell, M.G.A.; Gonçalves, M.C.; Pinto, L.R.; et al. Genome-wide approaches for the identification of markers and genes associated with sugarcane yellow leaf vírus resistance. Sci. Rep. 2021, 11, 15730. [Google Scholar] [CrossRef]
- Yang, X.; Sood, S.; Luo, Z.; Todd, J.; Wang, L. Genome-wide association studies identified resistance loci to orange rust and yellow leaf virus diseases in sugarcane (Saccharum spp.). Phytopathology 2019, 109, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Akbar, W.; Showier, A.T.; White, W.H.; Reagan, T.E. Categorizing sugarcane cultivars for resistance to the sugarcane aphid and yellow sugarcane aphid (Hemiptera: Aphididae). J. Econ. Entomol. 2010, 103, 1431–1437. [Google Scholar] [CrossRef]
- Akbar, W.; Showler, A.T.; Beuzelin, J.M.; Reagan, T.E.; Gravois, K.A. Evaluation of Aphid Resistance Among Sugarcane Cultivars in Louisiana. Ann. Entomol. Soc. Am. 2011, 104, 699–704. [Google Scholar] [CrossRef]
- Fartek, B.; Nibouche, S.; Atiama-Nurbel, T.; Reynaud Costet, B.L. Genotypic variability of sugarcane resistance to the aphid Melanaphis sacchari, vector of the Sugarcane yellow leaf virus. Plant Breed. 2014, 133, 771–776. [Google Scholar] [CrossRef]
- White, W.H. Yellow sugarcane aphid (Homoptera: Aphididae) resistance mechanisms in selected cultivars. J. Econ. Entomol. 1990, 83, 2111–2114. [Google Scholar] [CrossRef]
- Atiri, G.I.; Ekpot, E.J.A.; Thottappilly, G. The effect of aphid-resistance in cowpea on infestation and development of Aphis cracivora and the transmission of cowpea aphid-borne mosaic virus. Ann. Appl. Biol. 1984, 104, 339–346. [Google Scholar] [CrossRef]
- Rodríguez-López, M.J.; Garzo, E.; Bonani, J.P.; Fereres, A.; Fernández-Muñoz, R.; Moriones, E. Whitefly resistance traits derived from the wild tomato Solanum pimpinellifolium affect the preference and feeding behavior of Bemisia tabaci and reduce the spread of Tomato yellow leaf curl virus. Phytopathology 2011, 101, 1191–1201. [Google Scholar] [CrossRef]
- Viswanathan, R.; Ramasubramanian, T.; Chinnaraja, C.; Selvakumar, R.; Lakshmi Pathy, T.; Manivannan, K.; Nithyanantham, R. Population dynamics of Melanaphis sacchari (Zehntner), the aphid vector of sugarcane yellow leaf virus under tropical conditions in India. Trop Plant Pathol. 2022, 47, 260–277. [Google Scholar] [CrossRef]
- SAS Institute. SAS/STAT User’s Guide, Version 9.3; SAS: Cary, NC, USA, 2013. [Google Scholar]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.M.; Karlen, Y.; Bakker, O.; van den Hoff, M.J.; Moorman, A.F. Amplification efficiency: Linking baseline and bias in the analysis of quantitative PCR data. Nucleic Acids Res. 2009, 37, e45. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef]
- Ririe, K.M.; Rasmussen, R.P.; Wittwer, C.T. Product differentiation by analysis of DNA melting curves during the polymerase chain reaction. Anal. Biochem. 1997, 245, 154–160. [Google Scholar] [CrossRef]
- Matz, M.V.; Wright, R.M.; Scott, J.G. No control genes requires: Bayesian analysis of qRT-PCR data. PLoS ONE 2013, 8, e71448. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: https://www.R-project.org (accessed on 6 February 2023).
- Ramos, A.; Esteves, M.B.; Cortés, M.T.B.; Lopes, J.R.S. Maize Bushy Stunt Phytoplasma Favors Its Spread by Changing Host Preference of the Insect Vector. Insects 2020, 11, 600. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Genotypes | Symptom Evaluations | |||||||
|---|---|---|---|---|---|---|---|---|
| 6 MAP | 7 MAP | 8 MAP | 9 MAP | 10 MAP | 11 MAP | Mean | Class | |
| IACSP95-5094 | 1.07 c | 1.11 b | 1.28 cd | 1.78 cd | 1.84 c | 1.79 d | 1.48 d | MR |
| IACCTC05-2562 | 1.00 c | 1.00 b | 1.03 d | 1.43 de | 1.94 c | 1.78 d | 1.36 d | MR |
| IACSP01-5503 | 1.21 b | 1.17 b | 1.17 cd | 1.08 ef | 1.25 d | 1.35 e | 1.20 e | MR |
| IACSP96-7569 | 1.00 c | 1.15 b | 1.53 bc | 2.00 bc | 1.93 c | 2.1 c | 1.62 c | MR |
| IACBIO-266 | 1.00 c | 1.00 b | 1.00 d | 1.00 f | 1.00 d | 1.00 f | 1.00 f | R |
| IACSP01-3127 | 1.00 c | 1.00 b | 1.10 cd | 1.13 ef | 1.18 d | 1.38 e | 1.13 e | MR |
| IACSP95-5000 | 1.00 c | 1.00 b | 1.78 b | 2.26 b | 2.93 b | 3.12 b | 2.01 b | MS |
| SP71-6163 | 1.98 a | 2.46 a | 3.29 a | 3.98 a | 4.00 a | 4.00 a | 3.29 a | S |
| Source | DF | Sum of Squares | Mean Square | F Value | Pr > F |
| Model | 39 | 3.64844257 | 0.09354981 | 0.74 | 0.8748 |
| Error | 600 | 75.59045779 | 0.12598410 | ||
| Source | DF | Type III SS | Mean Square | F Value | Pr > F |
| Cultivar | 7 | 1.96570427 | 0.28081490 | 2.23 | 0.0305 |
| Time after release | 4 | 0.81747984 | 0.20436996 | 1.62 | 0.1670 |
| Cultivar x time | 28 | 0.86525846 | 0.03090209 | 0.25 | 1.0000 |
| CV (%) | 18.47719 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertasello, L.E.T.; da Silva, M.F.; Pinto, L.R.; Nóbile, P.M.; Carmo-Sousa, M.; dos Anjos, I.A.; Perecin, D.; Spotti Lopes, J.R.; Gonçalves, M.C. Yellow Leaf Disease Resistance and Melanaphis sacchari Preference in Commercial Sugarcane Cultivars. Plants 2023, 12, 3079. https://doi.org/10.3390/plants12173079
Bertasello LET, da Silva MF, Pinto LR, Nóbile PM, Carmo-Sousa M, dos Anjos IA, Perecin D, Spotti Lopes JR, Gonçalves MC. Yellow Leaf Disease Resistance and Melanaphis sacchari Preference in Commercial Sugarcane Cultivars. Plants. 2023; 12(17):3079. https://doi.org/10.3390/plants12173079
Chicago/Turabian StyleBertasello, Luiz Eduardo Tilhaqui, Marcel Fernando da Silva, Luciana Rossini Pinto, Paula Macedo Nóbile, Michele Carmo-Sousa, Ivan Antônio dos Anjos, Dilermando Perecin, João Roberto Spotti Lopes, and Marcos Cesar Gonçalves. 2023. "Yellow Leaf Disease Resistance and Melanaphis sacchari Preference in Commercial Sugarcane Cultivars" Plants 12, no. 17: 3079. https://doi.org/10.3390/plants12173079
APA StyleBertasello, L. E. T., da Silva, M. F., Pinto, L. R., Nóbile, P. M., Carmo-Sousa, M., dos Anjos, I. A., Perecin, D., Spotti Lopes, J. R., & Gonçalves, M. C. (2023). Yellow Leaf Disease Resistance and Melanaphis sacchari Preference in Commercial Sugarcane Cultivars. Plants, 12(17), 3079. https://doi.org/10.3390/plants12173079