
Meta-Analysis of the Effect of Nitric Oxide Application on Heavy Metal Stress Tolerance in Plants





Abstract
:1. Introduction
2. Results
2.1. Dataset
2.2. Plant Growth and Chlorophyll Concentration
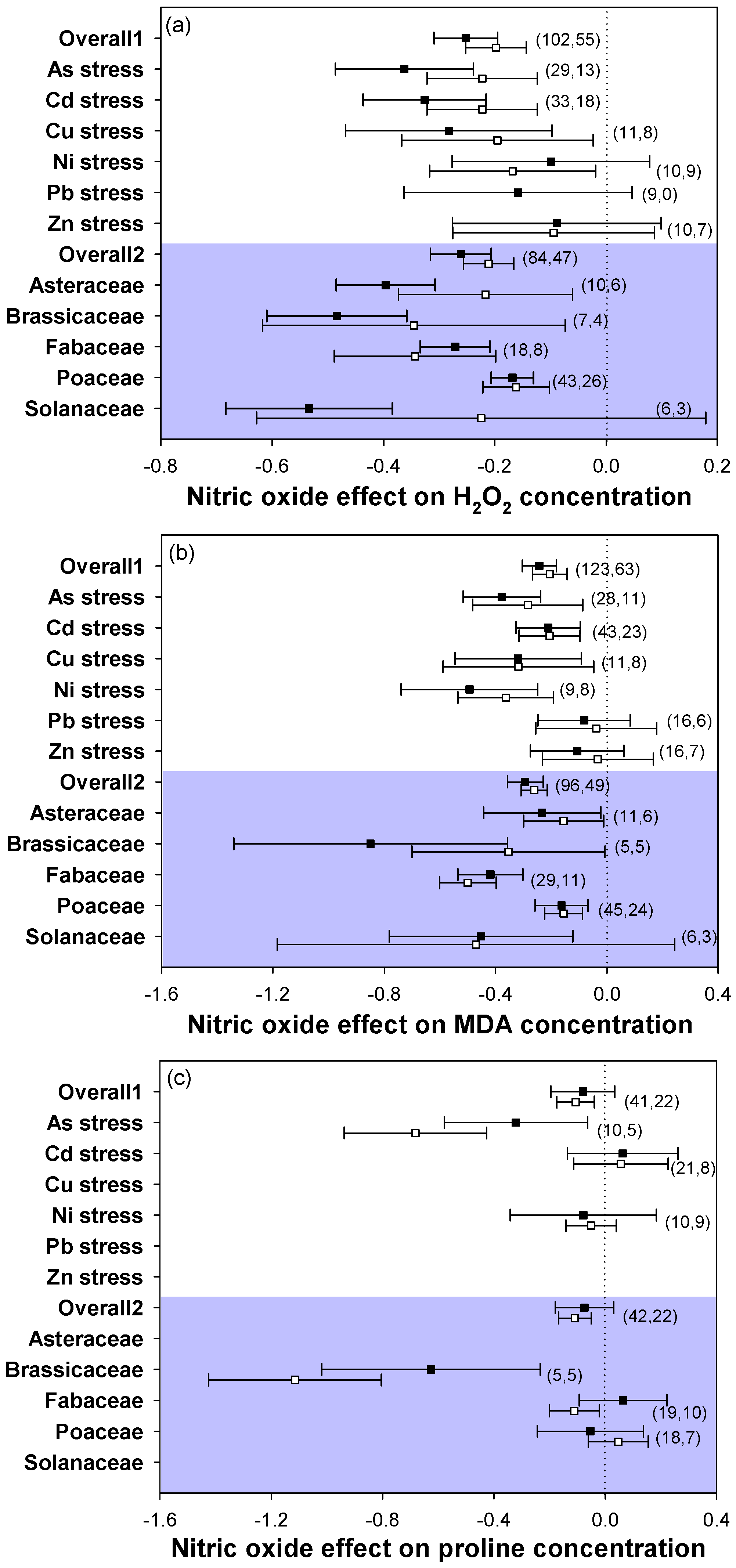
2.3. Oxidative Stress Markers
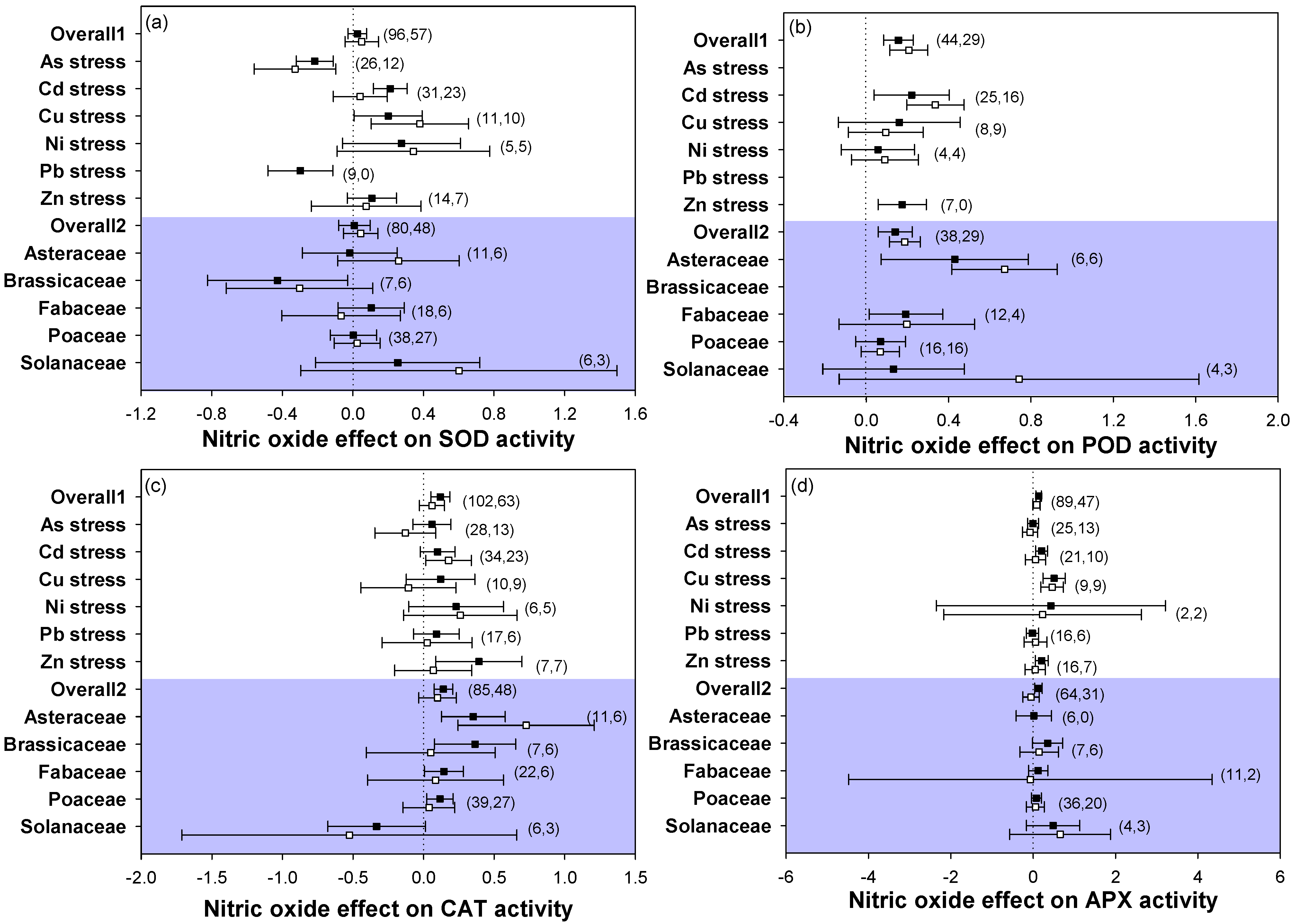
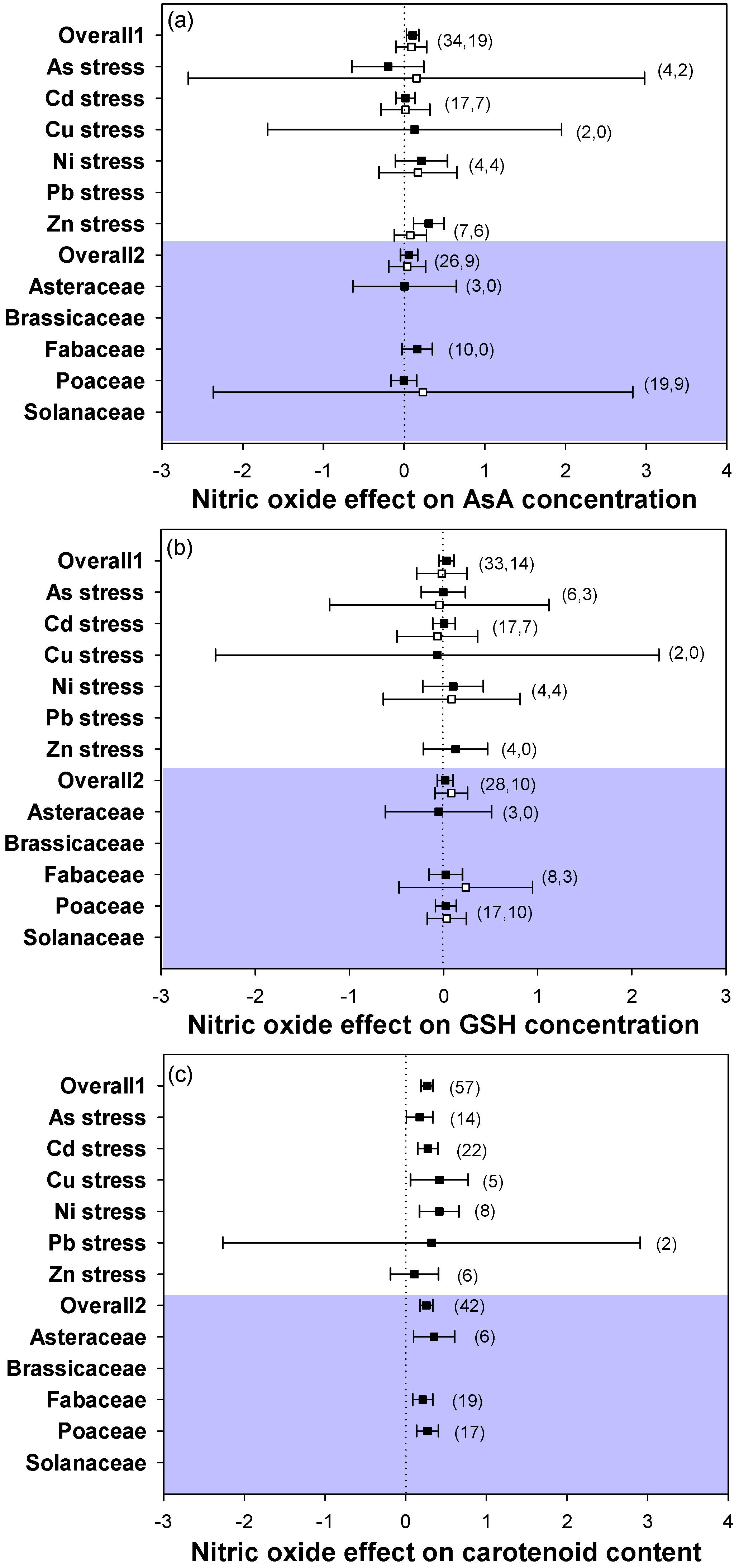
2.4. Antioxidant Responses
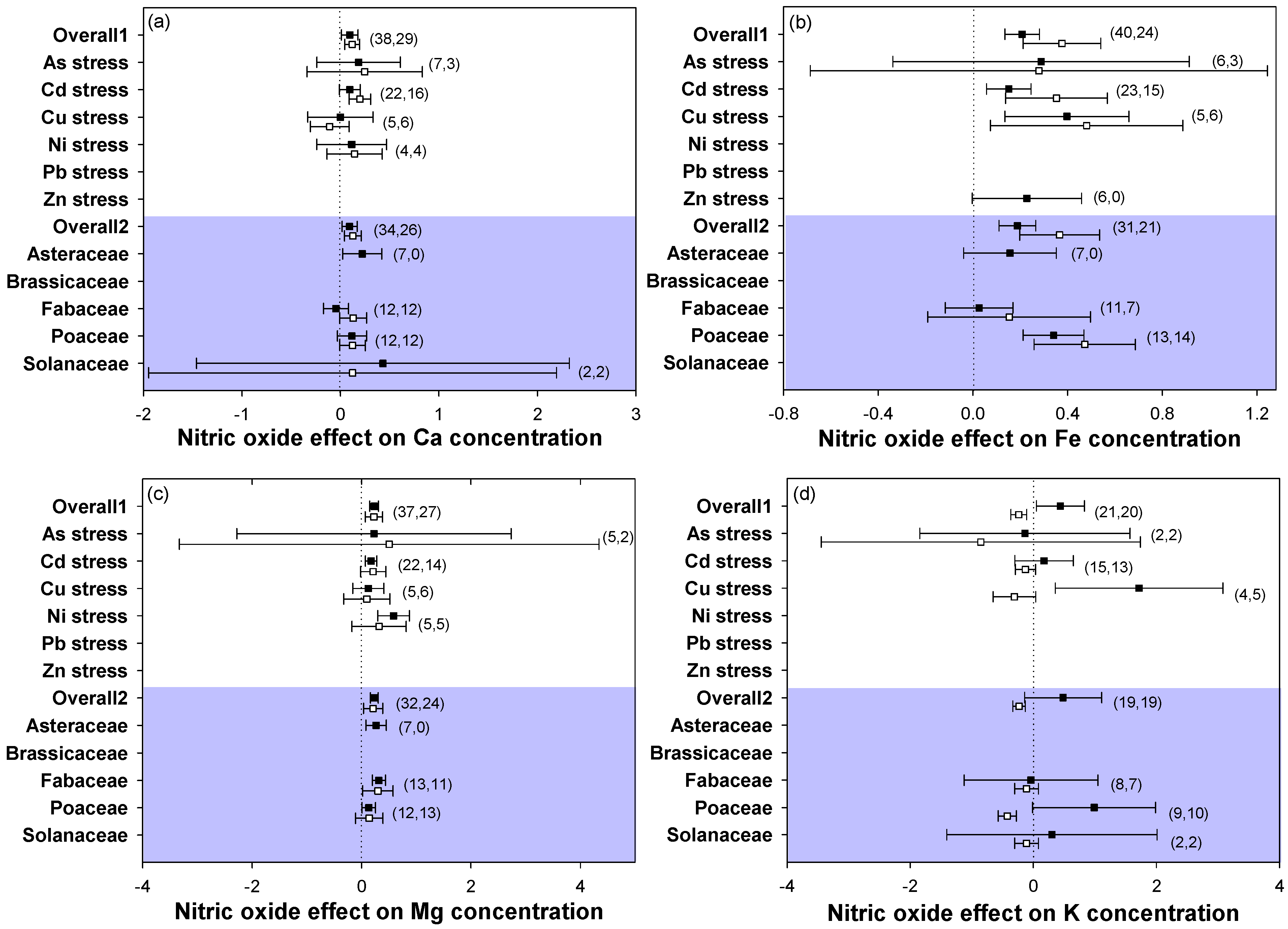
2.5. Element Content
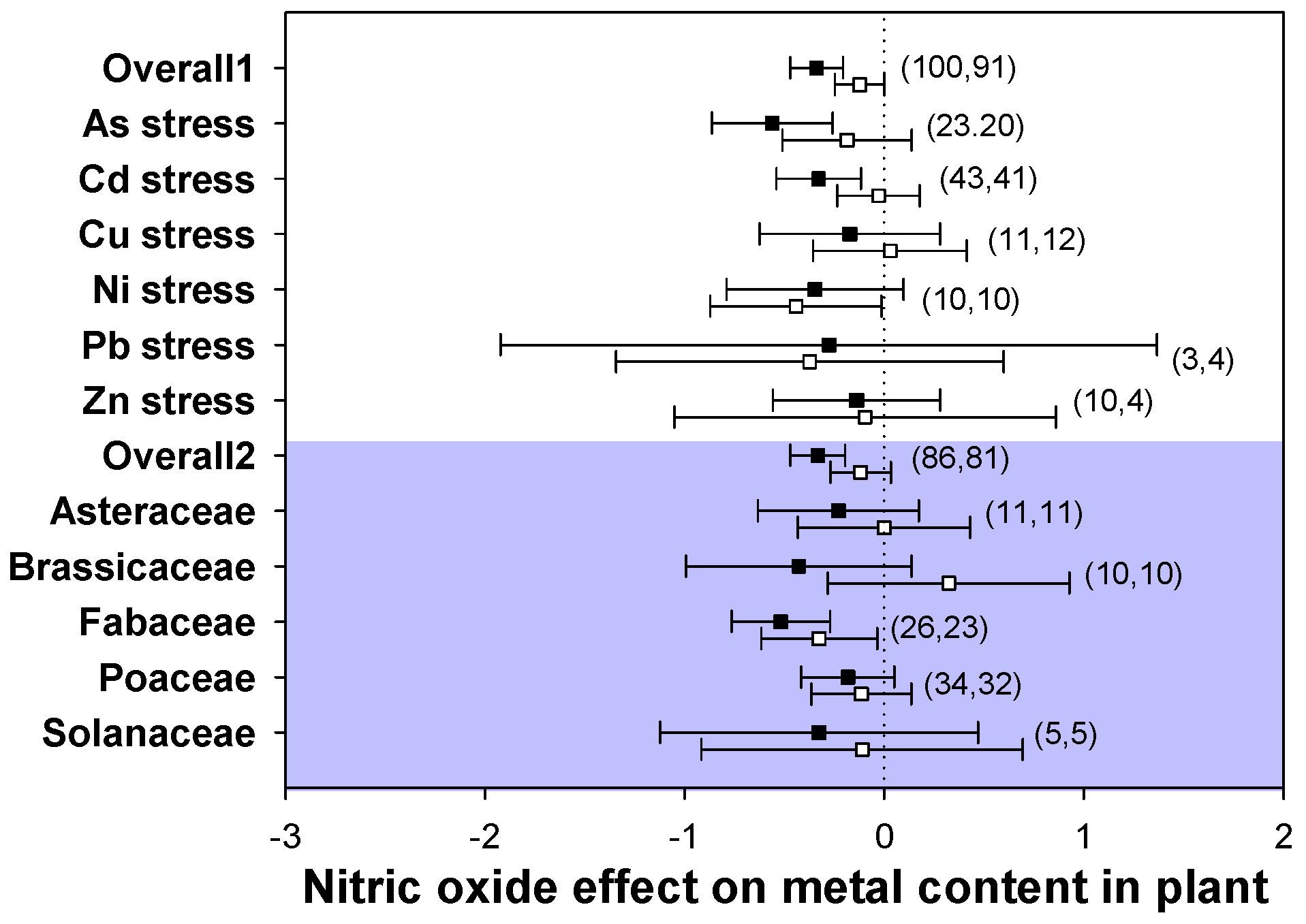
2.6. Heavy Metal Accumulation
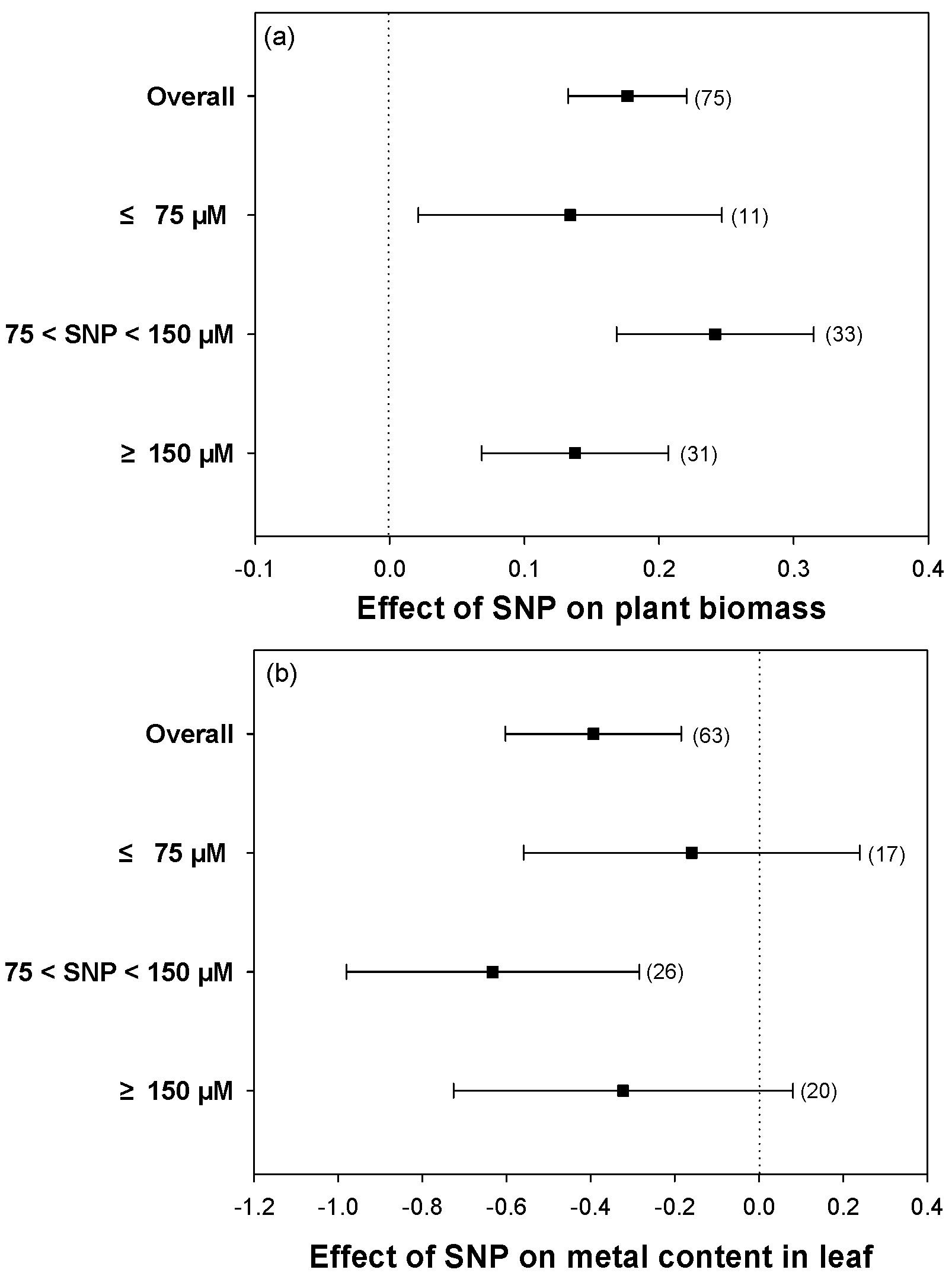
2.7. The Best Concentration of SNP for Hydroponic Experiments
3. Discussion
3.1. Application of NO Improved Plant Growth against Heavy Metal Stress
3.2. Application of NO Reduced Oxidative Damage under Heavy Metal Stress
3.3. Application of NO Increased Plant Antioxidant Ability under Heavy Metal Stress
3.4. Application of NO Increased the Contents of Essential Elements While Decreasing the Contents of Heavy Metal Elements under Heavy Metal Stress
3.5. Application of NO Is a Useful Approach in Alleviating Heavy Metal Stress
4. Materials and Methods
4.1. Data Collection
4.2. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| As | arsenic |
| Cd | cadmium |
| Cu | copper |
| Ni | nickel |
| Pb | lead |
| Zn | zinc |
| NO | nitric oxide |
| SNP | sodium nitroprusside |
| SOD | superoxide dismutase |
| CAT | catalase |
| APX | ascorbate peroxidase |
| POD | peroxidase |
| AsA | ascorbate |
| GSH | glutathione |
| H2O2 | hydrogen peroxide |
| MDA | malondialdehyde |
References
- Kowalska, J.B.; Mazurek, R.; Gasiorek, M.; Zaleski, T. Pollution indices as useful tools for the comprehensive evaluation of the degree of soil contamination-A review. Environ. Geochem. Health 2018, 40, 2395–2420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, S.R.; Kandeel, M.M.; Ghareeb, D.; Ghoneim, T.M.; Talha, N.I.; Alaoui-Sosse, B.; Aleya, L.; Abdel-Daim, M.M. Wheat biological responses to stress caused by cadmium, nickel and lead. Sci. Total Environ. 2020, 706, 136013. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Dumat, C.; Khalid, S.; Schreck, E.; Xiong, T.; Niazi, N.K. Foliar heavy metal uptake, toxicity and detoxification in plants: A comparison of foliar and root metal uptake. J. Hazard. Mater. 2017, 325, 36–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghori, N.H.; Ghor, I.T.; Hayat, M.Q.; Imadi, S.R.; Gul, A.; Altay, V.; Ozturk, M. Heavy metal stress and responses in plants. Int. J. Environ. Sci. Technol. 2019, 16, 1807–1828. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2000, 7, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, M.; Ma, L.Q.; Singh, N.; Singh, S. Antioxidant responses of hyper-accumulator and sensitive fern species to arsenic. J. Exp. Bot. 2005, 56, 1335–1342. [Google Scholar] [CrossRef] [Green Version]
- We, J.J.; Fei, X.Y.; Fang, H.Y. Protective effect of nitric oxide against arsenic-induced oxidative damage in tall fescue leaves. Afr. J. Biotechnol. 2010, 9, 1619–1627. [Google Scholar]
- Grun, S.; Lindermayr, C.; Sell, S.; Durner, J. Nitric oxide and gene regulation in plants. J. Exp. Bot. 2006, 57, 507–516. [Google Scholar] [CrossRef] [Green Version]
- Besson-Bard, A.; Pugin, A.; Wendehenne, D. New insights into nitric oxide signaling in plants. Annu. Rev. Plant Biol. 2008, 59, 21–39. [Google Scholar] [CrossRef]
- Wilson, I.D.; Neill, S.J.; Hancock, J.T. Nitric oxide synthesis and signalling in plants. Plant Cell Environ. 2008, 31, 622–631. [Google Scholar] [CrossRef]
- Khan, J.; Malangisha, G.K.; Ali, A.; Mahmoud, A.; Yang, J.H.; Zhang, M.F.; Hu, Z.Y. Nitric oxide alleviates lead toxicity by inhibiting lead translocation and regulating root growth in watermelon seedlings. Hortic. Environ. Biotechnol. 2021, 62, 701–714. [Google Scholar] [CrossRef]
- Nabi, R.B.S.; Tayade, R.; Hussain, A.; Kulkarni, K.P.; Imran, Q.M.; Mun, B.G.; Yun, B.W. Nitric oxide regulates plant responses to drought, salinity, and heavy metal stress. Environ. Exp. Bot. 2019, 161, 120–133. [Google Scholar] [CrossRef]
- Gong, B.; Nie, W.; Yan, Y.; Gao, Z.; Shi, Q. Unravelling cadmium toxicity and nitric oxide induced tolerance in Cucumis sativus: Insight into regulatory mechanisms using proteomics. J. Hazard. Mater. 2017, 336, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Li, S.W.; Li, Y.; Leng, Y.; Zeng, X.Y.; Ma, Y.H. Nitric oxide donor improves adventitious rooting in mung bean hypocotyl cuttings exposed to cadmium and osmotic stresses. Environ. Exp. Bot. 2019, 164, 114–123. [Google Scholar] [CrossRef]
- Praveen, A.; Pandey, A.; Gupta, M. Nitric oxide alters nitrogen metabolism and PIN gene expressions by playing protective role in arsenic challenged Brassica juncea L. Ecotoxicol. Environ. Saf. 2019, 176, 95–107. [Google Scholar] [CrossRef]
- Hu, K.D.; Hu, L.Y.; Li, Y.H.; Zhang, F.Q.; Zhang, H. Protective roles of nitric oxide on germination and antioxidant metabolism in wheat seeds under copper stress. Plant Growth Regul. 2007, 53, 173–183. [Google Scholar] [CrossRef]
- Liu, S.; Yang, R.; Pan, Y.; Ren, B.; Chen, Q.; Li, X.; Xiong, X.; Tao, J.; Cheng, Q.; Ma, M. Beneficial behavior of nitric oxide in copper-treated medicinal plants. J. Hazard. Mater. 2016, 314, 140–154. [Google Scholar] [CrossRef]
- Amooaghaie, R.; Zangene-Madar, F.; Enteshari, S. Role of two-sided crosstalk between NO and H2S on improvement of mineral homeostasis and antioxidative defense in Sesamum indicum under lead stress. Ecotoxicol. Environ. Saf. 2017, 139, 210–218. [Google Scholar] [CrossRef]
- Rizwan, M.; Mostofa, M.G.; Ahmad, M.Z.; Imtiaz, M.; Mehmood, S.; Adeel, M.; Dai, Z.; Li, Z.; Aziz, O.; Zhang, Y.; et al. Nitric oxide induces rice tolerance to excessive nickel by regulating nickel uptake, reactive oxygen species detoxification and defense-related gene expression. Chemosphere 2018, 191, 23–35. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Mishra, R.K.; Singh, S.; Singh, S.; Vishwakarma, K.; Sharma, S.; Singh, V.P.; Singh, P.K.; Prasad, S.M.; Dubey, N.K.; et al. Nitric oxide ameliorates zinc oxide nanoparticles phytotoxicity in wheat seedlings: Implication of the sscorbate-glutathione cycle. Front. Plant Sci. 2017, 8, 1. [Google Scholar] [CrossRef]
- Li, H.Y.; Shao, H. Exogenous Nitric oxide reduces cadmium toxicity of maize roots. Adv. Mater. Res. 2011, 378–379, 409–413. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Fujita, M. Exogenous sodium nitroprusside alleviates arsenic-induced oxidative stress in wheat (Triticum aestivum L.) seedlings by enhancing antioxidant defense and glyoxalase system. Ecotoxicology 2013, 22, 584–596. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Soares, C.; Sousa, B.; Martins, M.; Kumar, V.; Shahzad, B.; Sidhu, G.P.S.; Bali, A.S.; Asgher, M.; Bhardwaj, R.; et al. Nitric oxide-mediated regulation of oxidative stress in plants under metal stress: A review on molecular and biochemical aspects. Physiol. Plant. 2019, 168, 318–344. [Google Scholar] [CrossRef] [Green Version]
- Laspina, N.V.; Groppa, M.D.; Tomaro, M.L.; Benavides, M.P. Nitric oxide protects sunflower leaves against Cd-induced oxidative stress. Plant Sci. 2005, 169, 323–330. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernandez-Ruiz, J. Melatonin: A new plant hormone and/or a plant master regulator? Trends Plant Sci. 2019, 24, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.T.; Kao, C.H. Cadmium toxicity is reduced by nitric oxide in rice leaves. Plant Growth Regul. 2004, 42, 227–238. [Google Scholar] [CrossRef]
- Wang, Q.; Liang, X.; Dong, Y.; Xu, L.; Zhang, X.; Hou, J.; Fan, Z. Effects of exogenous nitric oxide on cadmium toxicity, element contents and antioxidative system in perennial ryegrass. Plant Growth Regul. 2012, 69, 11–20. [Google Scholar] [CrossRef]
- Dong, Y.J.; Chen, W.F.; Liu, F.Z.; Wan, Y.S. Physiological responses of peanut seedlings to exposure to low or high cadmium concentration and the alleviating effect of exogenous nitric oxide to high cadmium concentration stress. Plant Biosyst. 2019, 154, 405–412. [Google Scholar] [CrossRef]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.; An, L.; Lu, H.; Zhu, C. Exogenous nitric oxide enhances cadmium tolerance of rice by increasing pectin and hemicellulose contents in root cell wall. Planta 2009, 230, 755–765. [Google Scholar] [CrossRef]
- Xu, J.; Wang, W.; Yin, H.; Liu, X.; Sun, H.; Mi, Q. Exogenous nitric oxide improves antioxidative capacity and reduces auxin degradation in roots of Medicago truncatula seedlings under cadmium stress. Plant Soil 2009, 326, 321–330. [Google Scholar] [CrossRef]
- Wang, H.; Li, H.; Cai, B.; Huang, Z.X.; Sun, H. The effect of nitric oxide on metal release from metallothionein-3: Gradual unfolding of the protein. J. Biol. Inorg. Chem. 2008, 13, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Graziano, M.; Lamattina, L. Nitric oxide and iron in plants: An emerging and converging story. Trends Plant Sci. 2005, 10, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.P.; Mehta, S.K.; Liu, Z.P.; Yang, Z.M. Copper-induced proline synthesis is associated with nitric oxide generation in Chlamydomonas reinhardtii. Plant Cell Physiol. 2008, 49, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, U. Enhancing phytoextraction: The effect of chemical soil manipulation on mobility, plant accumulation, and leaching of heavy metals. J. Environ. Qual. 2003, 32, 1939–1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Israr, M.; Sahi, S.V.; Jain, J. Cadmium accumulation and antioxidative responses in the Sesbania drummondii callus. Arch. Environ. Contam. Toxicol. 2006, 50, 121–127. [Google Scholar] [CrossRef]
- Diwan, H.; Ahmad, A.; Iqbal, M. Genotypic variation in the phytoremediation potential of Indian mustard for chromium. Environ. Manag. 2008, 41, 734–741. [Google Scholar] [CrossRef]
- Khan, I.; Ahmad, A.; Iqbal, M. Modulation of antioxidant defence system for arsenic detoxification in Indian mustard. Ecotoxicol. Environ. Saf. 2009, 72, 626–634. [Google Scholar] [CrossRef]
- Pandey, V.; Dixit, V.; Shyam, R. Antioxidative responses in relation to growth of mustard (Brassica juncea cv. Pusa Jaikisan) plants exposed to hexavalent chromium. Chemosphere 2005, 61, 40–47. [Google Scholar] [CrossRef]
- Metwally, A.; Safronova, V.I.; Belimov, A.A.; Dietz, K.J. Genotypic variation of the response to cadmium toxicity in Pisum sativum L. J. Exp. Bot. 2005, 56, 167–178. [Google Scholar] [CrossRef]
- Talukdar, D. Arsenic-induced oxidative stress in the common bean legume, Phaseolus vulgaris L. seedlings and its amelioration by exogenous nitric oxide. Physiol. Mol. Biol. Plants 2013, 19, 69–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.L.; Fan, Z.Y.; Dong, Y.J.; Kong, J.; Bai, X.Y. Effects of exogenous salicylic acid and nitric oxide on physiological characteristics of two peanut cultivars under cadmium stress. Biol. Plant. 2014, 59, 171–182. [Google Scholar] [CrossRef]
- Singh, A.P.; Dixit, G.; Kumar, A.; Mishra, S.; Singh, P.K.; Dwivedi, S.; Trivedi, P.K.; Chakrabarty, D.; Mallick, S.; Pandey, V.; et al. Nitric oxide alleviated arsenic toxicity by modulation of antioxidants and thiol metabolism in rice (Oryza sativa L.). Front. Plant Sci. 2016, 6, 1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadeghipour, O. Pretreatment with nitric oxide reduces lead toxicity in cowpea (Vigna unguiculata [L.] walp.). Arch. Biol. Sci. 2016, 68, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Chen, W.; Xu, L.; Kong, J.; Liu, S.; He, Z. Nitric oxide can induce tolerance to oxidative stress of peanut seedlings under cadmium toxicity. Plant Growth Regul. 2015, 79, 19–28. [Google Scholar] [CrossRef]
- Zhao, H.; Jin, Q.; Wang, Y.; Chu, L.; Li, X.; Xu, Y. Effects of nitric oxide on alleviating cadmium stress in Typha angustifolia. Plant Growth Regul. 2015, 78, 243–251. [Google Scholar] [CrossRef]
- Mur, L.A.; Mandon, J.; Persijn, S.; Cristescu, S.M.; Moshkov, I.E.; Novikova, G.V.; Hall, M.A.; Harren, F.J.; Hebelstrup, K.H.; Gupta, K.J. Nitric oxide in plants: An assessment of the current state of knowledge. AOB Plants 2013, 5, pls052. [Google Scholar] [CrossRef]
- Brett, M.T. Meta-analysis in Ecology. Bull. Ecol. Soc. Am. 1997, 78, 92–94. [Google Scholar]
- Khairy, A.I.H.; Oh, M.J.; Lee, S.M.; Kim, D.S.; Roh, K.S. Nitric oxide overcomes Cd and Cu toxicity in in vitro-grown tobacco plants through increasing contents and activities of rubisco and rubisco activase. Biochimie 2016, 2, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, P.; Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alam, P. Exogenous application of nitric oxide modulates osmolyte metabolism, antioxidants, enzymes of ascorbate-glutathione cycle and promotes growth under cadmium stress in tomato. Protoplasma 2018, 255, 79–93. [Google Scholar] [CrossRef]
- Kohli, S.K.; Handa, N.; Bali, S.; Khanna, K.; Arora, S.; Sharma, A.; Bhardwaj, R. Current scenario of Pb toxicity in pants: Unraveling plethora of pysiological responses. Rev. Environ. Contam. Toxicol. 2019, 249, 153–197. [Google Scholar]
- Soares, C.; Carvalho, M.E.A.; Azevedo, R.A.; Fidalgo, F. Plants facing oxidative challenges—A little help from the antioxidant networks. Environ. Exp. Bot. 2019, 161, 4–25. [Google Scholar] [CrossRef]
- He, J.; Ren, Y.; Chen, X.; Chen, H. Protective roles of nitric oxide on seed germination and seedling growth of rice (Oryza sativa L.) under cadmium stress. Ecotoxicol. Environ. Saf. 2014, 108, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Phang, I.C.; Leung, D.W.; Taylor, H.H.; Burritt, D.J. The protective effect of sodium nitroprusside (SNP) treatment on Arabidopsis thaliana seedlings exposed to toxic level of Pb is not linked to avoidance of Pb uptake. Ecotoxicol. Environ. Saf. 2011, 74, 1310–1315. [Google Scholar] [CrossRef]
- Freeman, J.L.; Persans, M.W.; Nieman, K.; Albrecht, C.; Peer, W.; Pickering, I.J.; Salt, D.E. Increased glutathione biosynthesis plays a role in nickel tolerance in thlaspi nickel hyperaccumulators. Plant Cell 2004, 16, 2176–2191. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [Green Version]
- Shukla, P.; Singh, A.K. Nitric oxide mitigates arsenic-induced oxidative stress and genotoxicity in Vicia faba L. Environ. Sci. Pollut. Res. 2015, 22, 13881–13891. [Google Scholar] [CrossRef]
- Liu, S.; Yang, R.; Pan, Y.; Ma, M.; Pan, J.; Zhao, Y.; Cheng, Q.; Wu, M.; Wang, M.; Zhang, L. Nitric oxide contributes to minerals absorption, proton pumps and hormone equilibrium under cadmium excess in Trifolium repens L. plants. Ecotoxicol. Environ. Saf. 2015, 119, 35–46. [Google Scholar] [CrossRef]
- Courtois, C.; Besson, A.; Dahan, J.; Bourque, S.; Dobrowolska, G.; Pugin, A.; Wendehenne, J.D. Nitric oxide signalling in plants: Interplays with Ca2+ and protein kinases. Exp. Bot. 2008, 59, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Besson-Bard, A.; Gravot, A.; Richaud, P.; Auroy, P.; Duc, C.; Gaymard, F.; Taconnat, L.; Renou, J.P.; Pugin, A.; Wendehenne, D. Nitric oxide contributes to cadmium toxicity in Arabidopsis by promoting cadmium accumulation in roots and by up-regulating genes related to iron uptake. Plant Physiol. 2009, 149, 1302–1315. [Google Scholar] [CrossRef] [Green Version]
- Szczerba, M.W.; Britto, D.T.; Kronzucker, H.J. K+ transport in plants: Physiology and molecular biology. J. Plant Physiol. 2009, 166, 447–466. [Google Scholar] [CrossRef] [PubMed]
- Praveen, A.; Gupta, M. Nitric oxide confronts arsenic stimulated oxidative stress and root architecture through distinct gene expression of auxin transporters, nutrient related genes and modulates biochemical responses in Oryza sativa L. Environ. Pollut. 2018, 240, 950–962. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Akram, N.A.; Sürücü, A.; Ashraf, M. Alleviating effect of nitric oxide on oxidative stress and antioxidant defence system in pepper (Capsicum annuum L.) plants exposed to cadmium and lead toxicity applied separately or in combination. Sci. Hortic. 2019, 255, 52–60. [Google Scholar] [CrossRef]
- Dong, Y.; Chen, W.; Bai, X.; Liu, F.; Wan, Y. Effects of exogenous nitric oxide and 24-Epibrassinolide on the physiological characteristics of peanut seedlings under cadmium stress. Pedosphere 2019, 29, 45–59. [Google Scholar] [CrossRef]
- Bai, X.Y.; Dong, Y.J.; Wang, Q.H.; Xu, L.L.; Kong, J.; Liu, S. Effects of lead and nitric oxide on photosynthesis, antioxidative ability, and mineral element content of perennial ryegrass. Biol. Plant. 2014, 59, 163–170. [Google Scholar] [CrossRef]
- Nasiri-Savadkoohi, S. Protective Rrole of exogenous nitric oxide against zinc toxicity in Plantago major L. Appl. Ecol. Environ. Res. 2017, 15, 511–524. [Google Scholar] [CrossRef]
- Singh, P.K.; Indoliya, Y.; Chauhan, A.S.; Singh, S.P.; Singh, A.P.; Dwivedi, S.; Tripathi, R.D.; Chakrabarty, D. Nitric oxide mediated transcriptional modulation enhances plant adaptive responses to arsenic stress. Sci. Rep. 2017, 7, 3592. [Google Scholar] [CrossRef]
- Dong, Y.; Xu, L.; Wang, Q.; Fan, Z.; Kong, J.; Bai, X. Effects of exogenous nitric oxide on photosynthesis, antioxidative ability, and mineral element contents of perennial ryegrass under copper stress. J. Plant Interact. 2013, 9, 402–411. [Google Scholar] [CrossRef] [Green Version]
- Burda, B.U.; O’Connor, E.A.; Webber, E.M.; Redmond, N.; Perdue, L.A. Estimating data from figures with a Web-based program: Considerations for a systematic review. Res. Synth. Methods 2017, 8, 258–262. [Google Scholar] [CrossRef]
- Hedges, L.V. The meta-analysis of response ratios in experimental ecology. Curtis Ecol. 1999, 80, 1150–1156. [Google Scholar] [CrossRef]








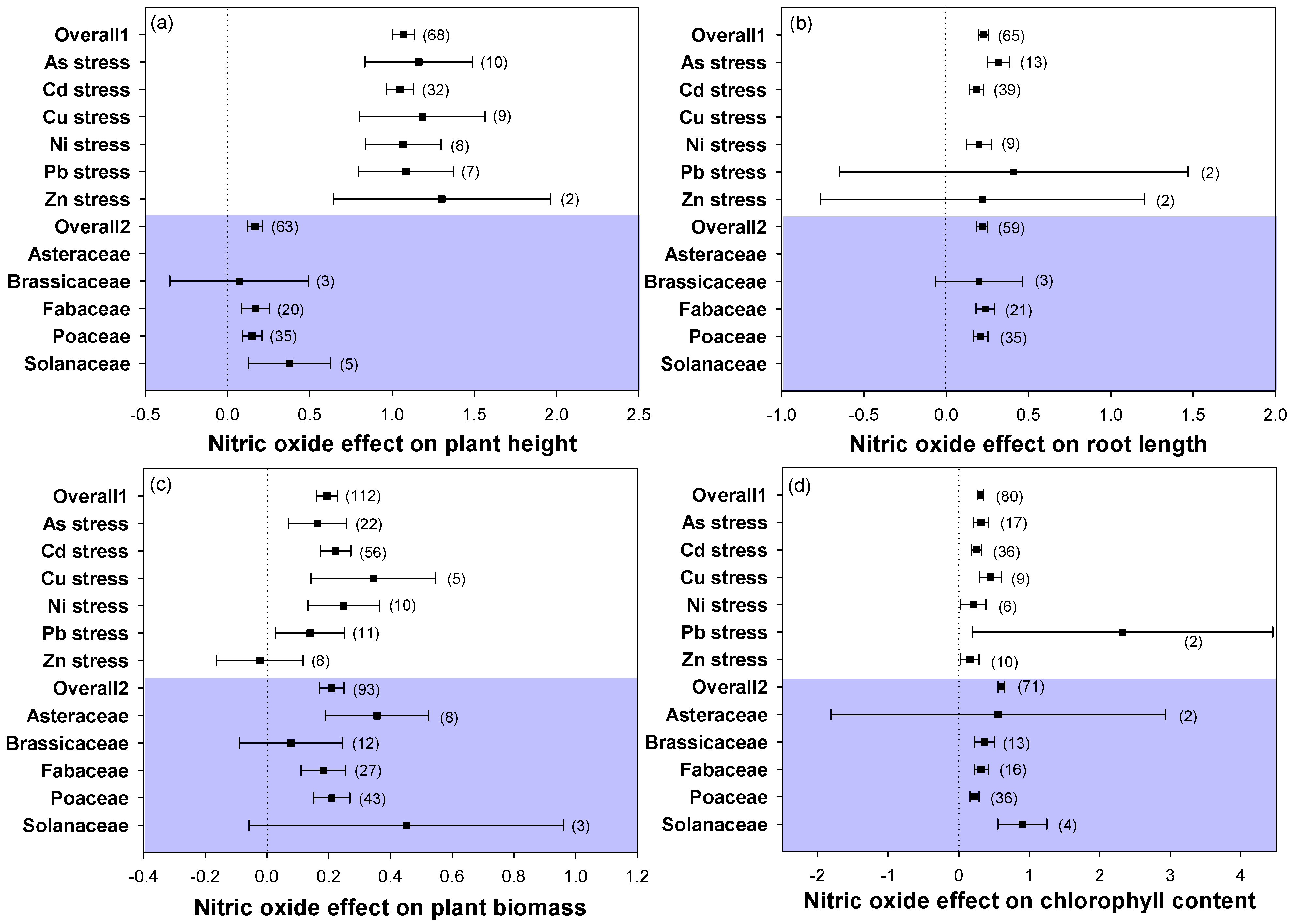{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species |
|---|---|
| Asteraceae | Carthamus tinctorius (safflower), Helianthus annuus (sunflower), Lactuca sativa (lettuce), Matricaria chamomilla (chamomile) |
| Brassicaceae | Arabidopsis thaliana, Brassica juncea (mustard), Brassica napus (canola), Nasturtium officinale (watercress), Isatis cappadocica |
| Fabaceae | Arachis hypogaea (peanut), Lupinus termis (lupine), Medicago sativa (alfalfa), Medicago truncatula (alfalfa), Phaseolus vulgaris (bean), Pisum sativum (pea), Trifolium repens (white clover), Vicia faba (faba bean), Vigna radiata (mungbean), Vigna unguiculata (cowpea) |
| Poaceae | Eleusine coracana (finger millet), Festuca arundinacea (tall fescue), Hordeum vulgare (hulless barley), Lolium perenne (ryegrass), Oryza sativa (rice), Triticum aestivum (wheat), Zea mays (maize) |
| Solanaceae | Capsicum annuum (pepper), Lycopersicon esculentum (tomato), Nicotiana tabacum (tobacco), Solanum lycopersicum (tomato) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Gong, D.; Ke, Q.; Yin, L.; Wang, S.; Gao, T. Meta-Analysis of the Effect of Nitric Oxide Application on Heavy Metal Stress Tolerance in Plants. Plants 2023, 12, 1494. https://doi.org/10.3390/plants12071494
Liu X, Gong D, Ke Q, Yin L, Wang S, Gao T. Meta-Analysis of the Effect of Nitric Oxide Application on Heavy Metal Stress Tolerance in Plants. Plants. 2023; 12(7):1494. https://doi.org/10.3390/plants12071494
Chicago/Turabian StyleLiu, Xiaoxiao, Di Gong, Qingbo Ke, Lina Yin, Shiwen Wang, and Tianpeng Gao. 2023. "Meta-Analysis of the Effect of Nitric Oxide Application on Heavy Metal Stress Tolerance in Plants" Plants 12, no. 7: 1494. https://doi.org/10.3390/plants12071494
APA StyleLiu, X., Gong, D., Ke, Q., Yin, L., Wang, S., & Gao, T. (2023). Meta-Analysis of the Effect of Nitric Oxide Application on Heavy Metal Stress Tolerance in Plants. Plants, 12(7), 1494. https://doi.org/10.3390/plants12071494