
Moringa oleifera Lam Leaf Extract Stimulates NRF2 and Attenuates ARV-Induced Toxicity in Human Liver Cells (HepG2)


Abstract
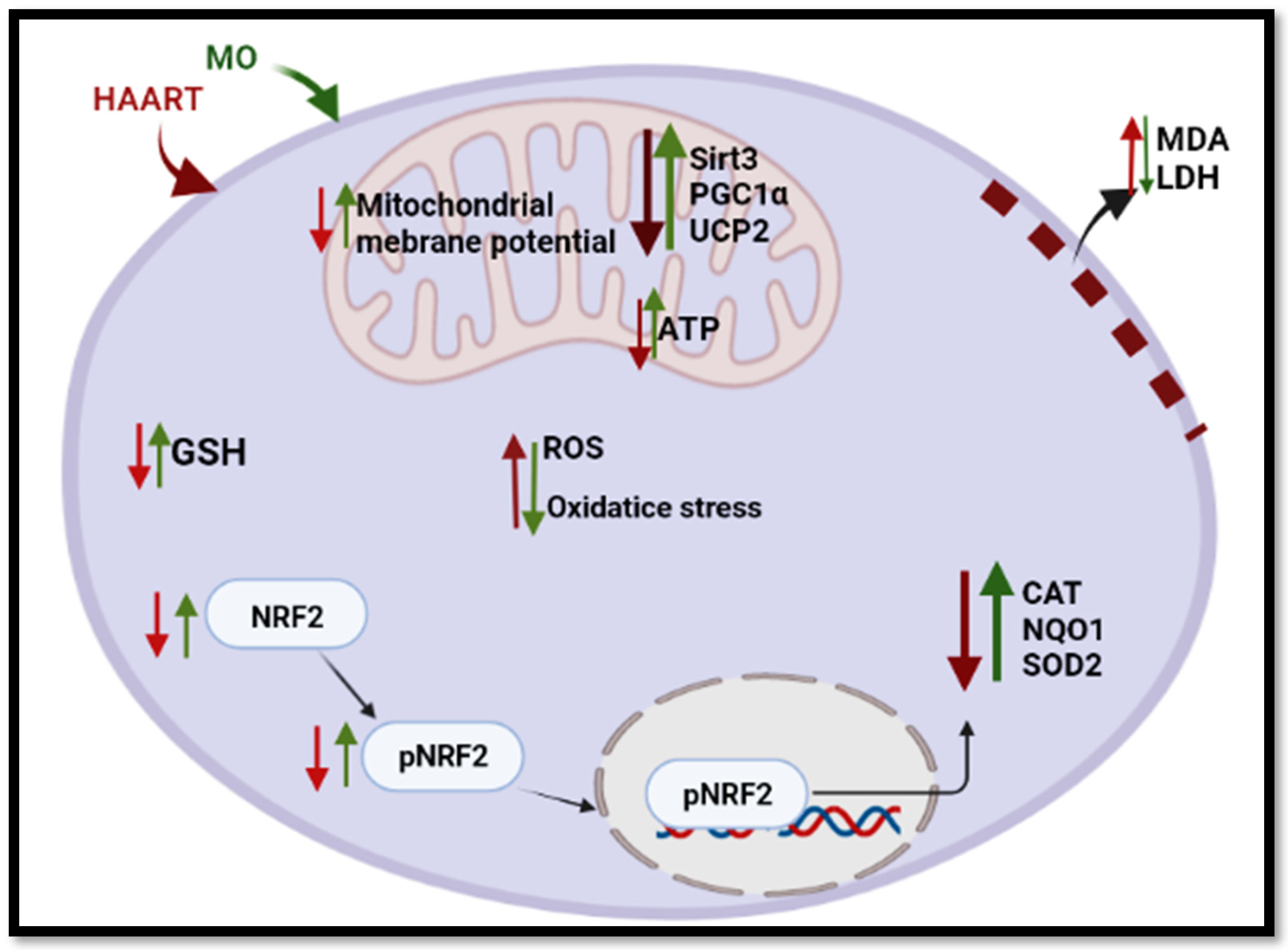
:1. Introduction
2. Results
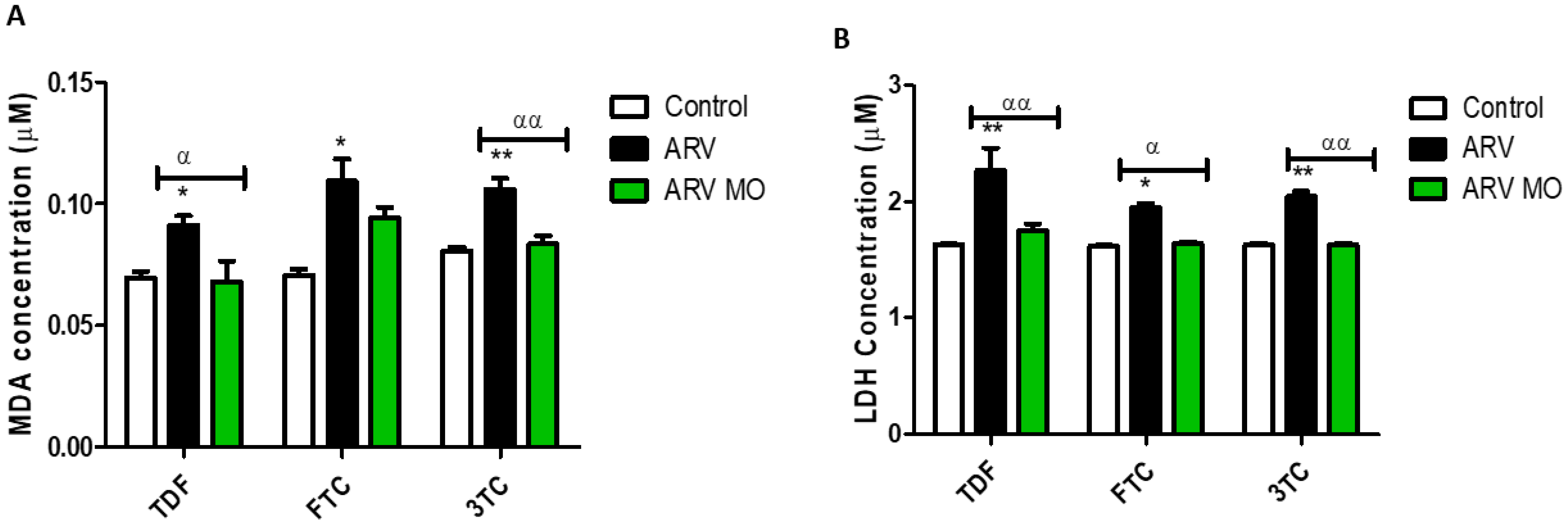
2.1. Oxidative Stress and Cellular Membrane Integrity
2.1.1. Induction of Oxidative Stress
2.1.2. Cellular Membrane Integrity
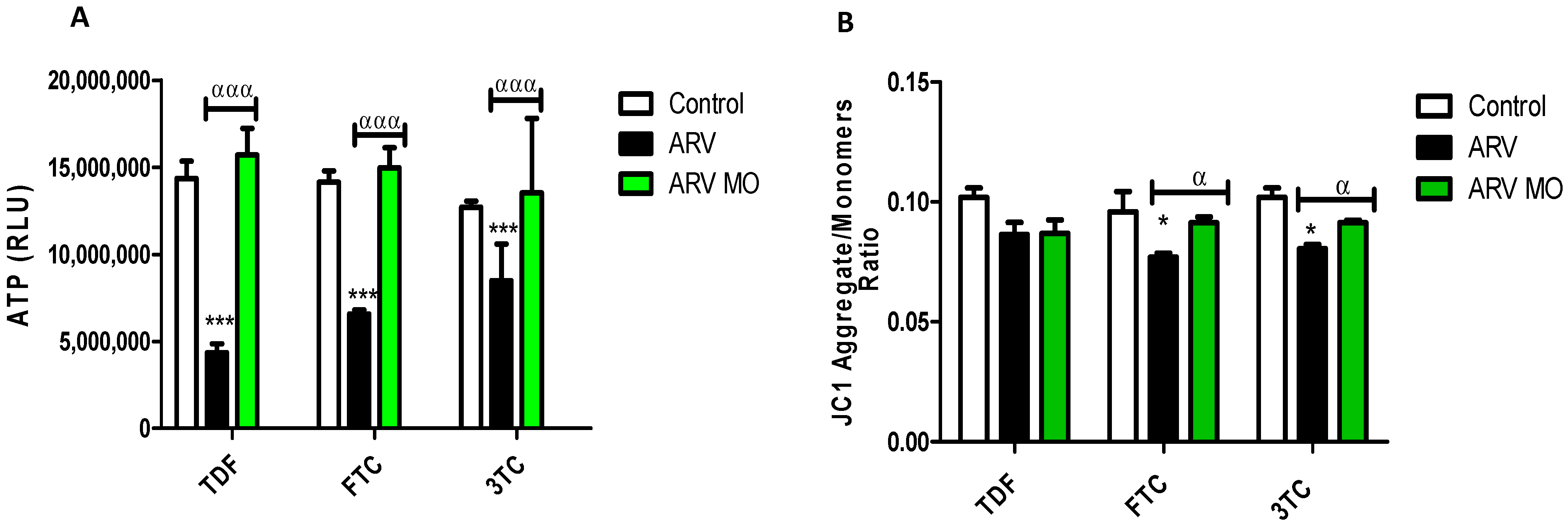
2.2. Mitochondrial Integrity
2.3. Intracellular GSH Expression
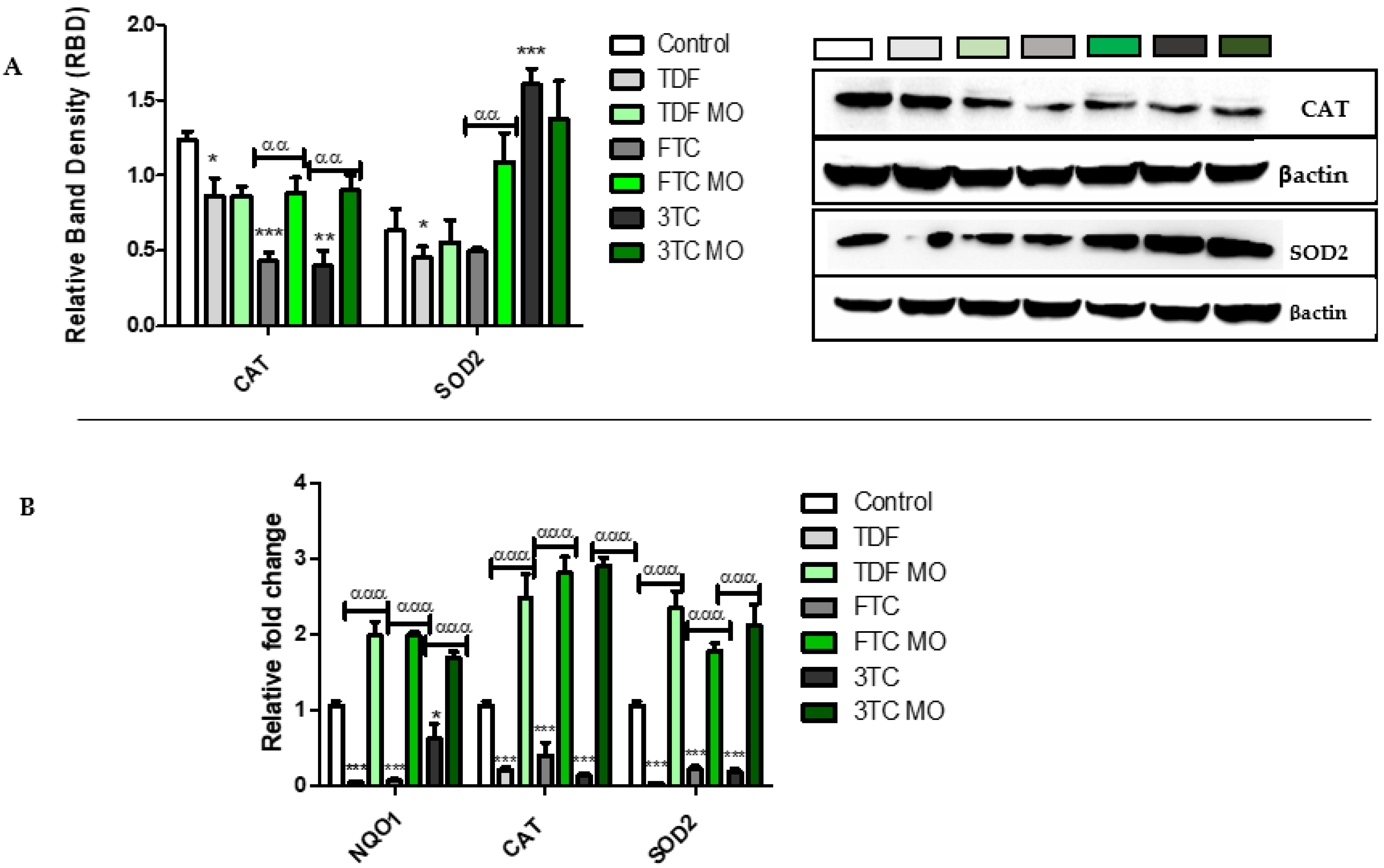
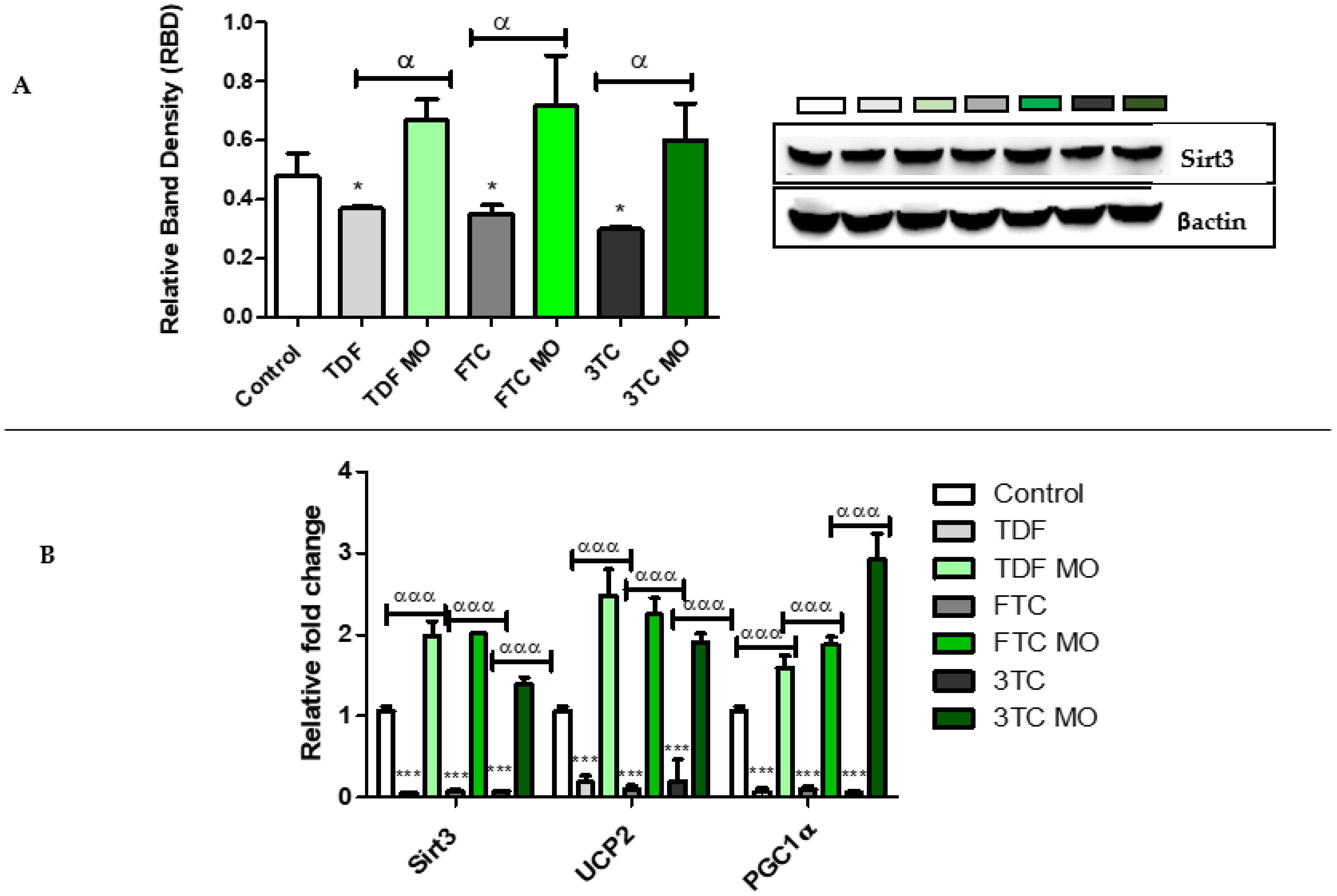
2.4. NRF2 Signalling Pathway and Related Antioxidants
2.4.1. NRF2 Expression
2.4.2. Cytoprotective Antioxidant Enzymes
2.4.3. Mitochondrial Protective Enzymes
3. Discussion
4. Materials and Methods
4.1. Moringa oleifera Lam Leaf Extracts Preparation
4.2. Drug Solutions and Treatment Preparation
4.3. Cell Culture and Treatments
4.4. Oxidative Stress Assessment
4.4.1. 2′,7′-Dichlorodihydrofluorescein-diacetate (H2DCF-DA) Assay
4.4.2. Thiobarbituric acid Reactive Substances (TBARS) Assay
4.4.3. Lactate Dehydrogenase (LDH) Assay
4.5. Mitochondrial Integrity
4.5.1. Adenosine Triphosphate (ATP) Assay
4.5.2. Mitochondrial Membrane Potential
4.6. Antioxidants Assays
4.6.1. Glutathione Assay
4.6.2. Western Blots
4.6.3. qPCR
4.7. Statistical Significance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Summary of the Global HIV Epidemic. 2020. Available online: https://www.who.int/data/gho/data/themes/hiv-aids (accessed on 14 June 2022).
- OMS. UNAIDS-Global HIV & AIDS Statistics—2020 Fact Sheet. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 12 July 2022).
- Department of Statistics South Africa. STATISTICAL RELEASE-P0302 Mid-Year Population Estimates. 2021. Available online: https://www.statssa.gov.za/publications/P0302/P03022022.pdf (accessed on 4 March 2022).
- Kim, H.; Tanser, F.; Tomita, A.; Vandormael, A.; Cuadros, D.F. Beyond HIV prevalence: Identifying people living with HIV within underserved areas in South Africa. BMJ Glob. Health 2021, 6, e004089. [Google Scholar] [CrossRef] [PubMed]
- Shafer, R.; Vuitton, D. Highly active antiretroviral therapy (Haart) for the treatment of infection with human immunodeficiency virus type 1. Biomed. Pharmacother. 1999, 53, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Hofman, P.; Nelson, A.M. The pathology induced by highly active antiretroviral therapy against human immunodeficiency virus: An update. Curr. Med. Chem. 2006, 13, 3121–3132. [Google Scholar] [CrossRef]
- Sierra, S.; Walter, H. Targets for Inhibition of HIV Replication: Entry, Enzyme Action, Release and Maturation. Intervirology 2012, 55, 84–97. [Google Scholar] [CrossRef] [Green Version]
- Delaney, M. History of HAART—The true story of how effective multi-drug therapy was developed for treatment of HIV disease. Retrovirology 2006, 3, S6. [Google Scholar] [CrossRef] [Green Version]
- Thompson, M.A.; Aberg, J.A.; Hoy, J.F. Antiretroviral treatment of adult HIV infection: 2012 recommendations of the International Antiviral Society-USA panel. JAMA 2012, 308, 387–402. [Google Scholar] [CrossRef] [PubMed]
- Llibre, J.M.; Clotet, B. Once-daily single-tablet regimens: A long and winding road to excellence in antiretroviral treatment. Aids Rev. 2012, 14, 168–178. [Google Scholar]
- Caplan, M.R.; Daar, E.S.; Corado, K.C. Next generation fixed dose combination pharmacotherapies for treating HIV. Expert Opin. Pharmacother. 2018, 19, 589–596. [Google Scholar] [CrossRef]
- Mohan, J.; Ghazi, T.; Chuturgoon, A.A. A Critical Review of the Biochemical Mechanisms and Epigenetic Modifications in HIV- and Antiretroviral-Induced Metabolic Syndrome. Int. J. Mol. Sci. 2021, 22, 12020. [Google Scholar] [CrossRef]
- Bertrand, L.; Velichkovska, M.; Toborek, M. Cerebral Vascular Toxicity of Antiretroviral Therapy. J. Neuroimmune Pharmacol. 2019, 16, 74–89. [Google Scholar] [CrossRef] [Green Version]
- Gyurászová, M.; Gurecká, R.; Bábíčková, J.; Tóthová, Ľ. Oxidative Stress in the Pathophysiology of Kidney Disease: Implications for Noninvasive Monitoring and Identification of Biomarkers. Oxidative Med. Cell. Longev. 2020, 2020, 1–11. [Google Scholar] [CrossRef] [Green Version]
- White, A.J. Mitochondrial toxicity and HIV therapy. Sex. Transm. Infect. 2001, 77, 158–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feeney, E.R.; Mallon, P.W. Impact of mitochondrial toxicity of HIV-1 antiretroviral drugs on lipodystrophy and metabolic dysregulation. Curr. Pharm. Des. 2010, 16, 3339–3351. [Google Scholar]
- Nooka, S.; Ghorpade, A. HIV-1-associated inflammation and antiretroviral therapy regulate astrocyte endoplasmic reticulum stress responses. Cell Death Discov. 2017, 3, 17061. [Google Scholar] [CrossRef] [Green Version]
- Ganta, K.K.; Chaubey, B. Mitochondrial dysfunctions in HIV infection and antiviral drug treatment. Expert Opin. Drug Metab. Toxicol. 2019, 15, 1043–1052. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.H.; Dar, K.B.; Anees, S.; Zargar, M.A.; Masood, A.; Sofi, M.A.; Ganie, S.A. Oxidative stress, mitochondrial dysfunction and neurodegenerative diseases; a mechanistic insight. Biomed. Pharmacother. 2015, 74, 101–110. [Google Scholar] [CrossRef]
- Nagiah, S.; Phulukdaree, A.; Chuturgoon, A. Mitochondrial and Oxidative Stress Response in HepG2 Cells Following Acute and Prolonged Exposure to Antiretroviral Drugs. J. Cell. Biochem. 2015, 116, 1939–1946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [Green Version]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Jozefczak, M.; Remans, T.; Vangronsveld, J.; Cuypers, A. Glutathione Is a Key Player in Metal-Induced Oxidative Stress Defenses. Int. J. Mol. Sci. 2012, 13, 3145–3175. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.; Nioi, P.; Pickett, C.B. The Nrf2-Antioxidant Response Element Signaling Pathway and Its Activation by Oxidative Stress. J. Biol. Chem. 2009, 284, 13291–13295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q. Role of Nrf2 in Oxidative Stress and Toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [Green Version]
- Jaramillo, M.C.; Zhang, D.D. The emerging role of the Nrf2–Keap1 signaling pathway in cancer. Genes Dev. 2013, 27, 2179–2191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buendia, I.; Michalska, P.; Navarro, E.; Gameiro, I.; Egea, J.; León, R. Nrf2–ARE pathway: An emerging target against oxidative stress and neuroinflammation in neurodegenerative diseases. Pharmacol. Ther. 2016, 157, 84–104. [Google Scholar] [CrossRef]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2018, 1865, 721–733. [Google Scholar] [CrossRef]
- South African National Department of Health. National Consolidated Guidelines. 2020. Available online: https://sahivsoc.org/Files/ART%20Guidelines%2015052015.pdf (accessed on 21 July 2020).
- Abraham, P.; Ramamoorthy, H.; Isaac, B. Depletion of the cellular antioxidant system contributes to tenofovir disoproxil fumarate-induced mitochondrial damage and increased oxido-nitrosative stress in the kidney. J. Biomed. Sci. 2013, 20, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, E.S.; Jain, R.; Roxby, A.C. Abacavir/Dolutegravir/Lamivudine (Triumeq)–Induced Liver Toxicity in a Human Immunodeficiency Virus–Infected Patient. Open Forum Infect. Dis. 2017, 4, ofx122. [Google Scholar] [CrossRef] [Green Version]
- Schank, M.; Zhao, J.; Moorman, J.; Yao, Z. The Impact of HIV- and ART-Induced Mitochondrial Dysfunction in Cellular Senescence and Aging. Cells 2021, 10, 174. [Google Scholar] [CrossRef]
- Sahakitpichan, P.; Mahidol, C.; Disadee, W.; Ruchirawat, S.; Kanchanapoom, T. Unusual glycosides of pyrrole alkaloid and 4′-hydroxyphenylethanamide from leaves of Moringa oleifera. Phytochemistry 2011, 72, 791–795. [Google Scholar] [CrossRef]
- Leone, A.; Spada, A.; Battezzati, A.; Schiraldi, A.; Aristil, J.; Bertoli, S. Cultivation, Genetic, Ethnopharmacology, Phytochemistry and Pharmacology of Moringa oleifera Leaves: An Overview. Int. J. Mol. Sci. 2015, 16, 12791–12835. [Google Scholar] [CrossRef]
- Vergara-Jimenez, M.; Almatrafi, M.M.; Fernandez, M.L. Bioactive Components in Moringa Oleifera Leaves Protect against Chronic Disease. Antioxidants 2017, 6, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chhikara, N.; Kaur, A.; Mann, S.; Garg, M.K.; Sofi, S.A.; Panghal, A. Bioactive compounds, associated health benefits and safety considerations of Moringa oleifera L.: An updated review. Nutr. Food Sci. 2021, 51, 255–277. [Google Scholar] [CrossRef]
- González-Romero, J.; Guerra-Hernández, E.J.; Rodríguez-Pérez, C. Chapter 19-Bioactive compounds from Moringa oleifera as promising protectors of in vivo inflammation and oxidative stress processes. In Current Advances for Development of Functional Foods Modulating Inflammation and Oxidative Stress; Hernández-Ledesma, B., Martínez-Villaluenga, C., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 379–399. [Google Scholar]
- Silva, M.; Trancoso, J.; Tormen, L.; Bombardelli, M.M.; Corazza, M.L.; Bainy, E.M. Extraction of compounds from Moringa oleifera leaves using supercritical CO2 plus ethanol as a cosolvent. J. Food Process. Eng. 2022, 45, e13979. [Google Scholar] [CrossRef]
- Sreelatha, S.; Padma, P.R. Antioxidant Activity and Total Phenolic Content of Moringa oleifera Leaves in Two Stages of Maturity. Plant Foods Hum. Nutr. 2009, 64, 303–311. [Google Scholar] [CrossRef]
- Wright, R.J.; Lee, K.S.; Hyacinth, H.I.; Hibbert, J.M.; Reid, M.E.; Wheatley, A.O.; Asemota, H.N. An Investigation of the Antioxidant Capacity in Extracts from Moringa oleifera Plants Grown in Jamaica. Plants 2017, 6, 48. [Google Scholar] [CrossRef] [Green Version]
- Vongsak, B.; Sithisarn, P.; Gritsanapan, W. Bioactive contents and free radical scavenging activity of Moringa oleifera leaf extract under different storage conditions. Ind. Crop. Prod. 2013, 49, 419–421. [Google Scholar] [CrossRef]
- Khor, K.Z.; Lim, V.; Moses, E.J.; Samad, N.A. The In Vitro and In Vivo Anticancer Properties of Moringa oleifera. Evid. Based Complement. Alternat. Med. 2018, 2018, 1071243. [Google Scholar] [CrossRef] [Green Version]
- Sodvadiya, M.; Patel, H.; Mishra, A.; Nair, S. Emerging Insights into Anticancer Chemopreventive Activities of Nutraceutical Moringa oleifera: Molecular Mechanisms, Signal Transduction and In Vivo Efficacy. Curr. Pharmacol. Rep. 2020, 6, 38–51. [Google Scholar] [CrossRef]
- Toppo, R.; Roy, B.K.; Gora, R.H.; Baxla, S.L.; Kumar, P. Hepatoprotective activity of Moringa oleifera against cadmium toxicity in rats. Veter- World 2015, 8, 537–540. [Google Scholar] [CrossRef] [Green Version]
- Nova, E.; Redondo-Useros, N.; Martínez-García, R.M.; Gómez-Martínez, S.; Díaz-Prieto, L.E.; Marcos, A. Potential of Moringa oleifera to Improve Glucose Control for the Prevention of Diabetes and Related Metabolic Alterations: A Systematic Review of Animal and Human Studies. Nutrients 2020, 12, 2050. [Google Scholar] [CrossRef] [PubMed]
- Ruoß, M.; Damm, G.; Vosough, M.; Ehret, L.; Grom-Baumgarten, C.; Petkov, M.; Naddalin, S.; Ladurner, R.; Seehofer, D.; Nussler, A.; et al. Epigenetic Modifications of the Liver Tumor Cell Line HepG2 Increase Their Drug Metabolic Capacity. Int. J. Mol. Sci. 2019, 20, 347. [Google Scholar] [CrossRef] [Green Version]
- Sosa-Gutiérrez, J.A.; Valdéz-Solana, M.A.; Forbes-Hernández, T.Y.; Avitia-Domínguez, C.I.; Garcia-Vargas, G.G.; Salas-Pacheco, J.M.; Flores-Herrera, O.; Téllez-Valencia, A.; Battino, M.; Sierra-Campos, E. Effects of Moringa oleifera Leaves Extract on High Glucose-Induced Metabolic Changes in HepG2 Cells. Biology 2018, 7, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, L.T.; Minsky, N.W.; Espinosa, L.E.M.; Aranda, R.S.; Meseguer, J.P.; Pérez, P.C. In vitro assessment of hepatoprotective agents against damage induced by acetaminophen and CCl4. BMC Complement. Altern. Med. 2017, 17, 39. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, M.A.; Ali, Z.; Chittiboyina, A.; Khan, I.A. Hepatoprotective Effect of Steroidal Glycosides from Dioscorea villosa on Hydrogen Peroxide-Induced Hepatotoxicity in HepG2 Cells. Front. Pharmacol. 2018, 9, 797. [Google Scholar] [CrossRef] [Green Version]
- Paemanee, A.; Sornjai, W.; Kittisenachai, S.; Sirinonthanawech, N.; Roytrakul, S.; Wongtrakul, J.; Smith, D.R. Nevirapine induced mitochondrial dysfunction in HepG2 cells. Sci. Rep. 2017, 7, 9194. [Google Scholar] [CrossRef]
- Haron, M.H.; Avula, B.; Gurley, B.J.; Chittiboyina, A.G.; Khan, I.A.; Khan, S.I. Possible Herb-Drug Interaction Risk of Some Nutritional and Beauty Supplements on Antiretroviral Therapy in HIV Patients. J. Diet. Suppl. 2020, 19, 62–77. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, T.; Strigun, A.; Verlohner, A.; Huener, H.-A.; Peter, E.; Herold, M.; Bordag, N.; Mellert, W.; Walk, T.; Spitzer, M.; et al. Prediction of liver toxicity and mode of action using metabolomics in vitro in HepG2 cells. Arch. Toxicol. 2017, 92, 893–906. [Google Scholar] [CrossRef] [Green Version]
- Razis, A.F.A.; Ibrahim, M.D.; Kntayya, S.B. Health Benefits of Moringa oleifera. Asian Pac. J. Cancer Prev. 2014, 15, 8571–8576. [Google Scholar] [CrossRef] [Green Version]
- Rui, L. Energy metabolism in the liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar]
- Gerets, H.H.J.; Tilmant, K.; Gerin, B.; Chanteux, H.; Depelchin, B.O.; Dhalluin, S.; Atienzar, F.A. Characterization of primary human hepatocytes, HepG2 cells, and HepaRG cells at the mRNA level and CYP activity in response to inducers and their predictivity for the detection of human hepatotoxins. Cell Biol. Toxicol. 2012, 28, 69–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xuan, J.; Chen, S.; Ning, B.; Tolleson, W.H.; Guo, L. Development of HepG2-derived cells expressing cytochrome P450s for assessing metabolism-associated drug-induced liver toxicity. Chem. Interact. 2016, 255, 63–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Wu, Q.; Li, X.; Li, D.; Mei, N.; Ning, B.; Puig, M.; Ren, Z.; Tolleson, W.H.; Guo, L. Characterization of cytochrome P450s (CYP)-overexpressing HepG2 cells for assessing drug and chemical-induced liver toxicity. J. Environ. Sci. Health Part C 2021, 39, 68–86. [Google Scholar] [CrossRef]
- Loveday, C. International perspectives on antiretroviral resistance. Nucleoside reverse transcriptase inhibitor resistance. J. Acquir. Immune Defic. Syndr. 2001, 26, S10–S24. [Google Scholar] [CrossRef]
- Gu, S.-X.; Zhu, Y.-Y.; Wang, C.; Wang, H.-F.; Liu, G.-Y.; Cao, S.; Huang, L. Recent discoveries in HIV-1 reverse transcriptase inhibitors. Curr. Opin. Pharmacol. 2020, 54, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.Q.; Lichterfeld, M. Diversity of HIV-1 reservoirs in CD4+ T-cell subpopulations. Curr. Opin. HIV AIDS 2016, 11, 383–387. [Google Scholar] [CrossRef] [Green Version]
- Kobin, A.B.; Sheth, N.U. Levels of Adherence Required for Virologic Suppression Among Newer Antiretroviral Medications. Ann. Pharmacother. 2011, 45, 372–379. [Google Scholar] [CrossRef]
- Viswanathan, S.; Detels, R.; Mehta, S.H.; Macatangay, B.J.C.; Kirk, G.D.; Jacobson, L.P. Level of Adherence and HIV RNA Suppression in the Current Era of Highly Active Antiretroviral Therapy (HAART). AIDS Behav. 2014, 19, 601–611. [Google Scholar] [CrossRef] [Green Version]
- Margolis, A.M.; Heverling, H.; Pham, P.A.; Stolbach, A. A review of the toxicity of HIV medications. J. Med. Toxicol. 2014, 10, 26–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venter, W.F.; Kaiser, K.B.; Pillay, Y.; Conradie, F.; Gomez, G.B.; Clayden, P.; Matsolo, M.; Amole, C.; Rutter, L.; Abdullah, F.; et al. Cutting the cost of South African antiretroviral therapy using newer, safer drugs. S. Afr. Med. J. 2016, 107, 28–30. [Google Scholar] [CrossRef]
- Kohler, J.J.; Hosseini, S.H.; Hoying-Brandt, A.; Green, E.; Johnson, D.M.; Russ, R.; Tran, D.; Raper, C.M.; Santoianni, R.; Lewis, W. Tenofovir renal toxicity targets mitochondria of renal proximal tubules. Lab. Investig. 2009, 89, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Ramamoorthy, H.; Abraham, P.; Isaac, B.; Selvakumar, D. Mitochondrial pathway of apoptosis and necrosis contribute to tenofovir disoproxil fumarate–induced renal damage in rats. Hum. Exp. Toxicol. 2018, 38, 288–302. [Google Scholar] [CrossRef]
- Olojede, S.O.; Lawal, S.K.; Dare, A.; Naidu, E.C.S.; Rennie, C.O.; Azu, O.O. Evaluation of tenofovir disoproxil fumarate loaded silver nanoparticle on testicular morphology in experimental type-2 diabetic rats. Artif. Cells Nanomed. Biotechnol. 2022, 50, 71–80. [Google Scholar] [CrossRef]
- Oguntibeju, O.; Aboua, G.; Omodanisi, E. Effects of Moringa oleifera on oxidative stress, apoptotic and inflammatory biomarkers in streptozotocin-induced diabetic animal model. S. Afr. J. Bot. 2020, 129, 354–365. [Google Scholar] [CrossRef]
- Yetuk, G.; Pandir, D.; Bas, H. Protective Role of Catechin and Quercetin in Sodium Benzoate-Induced Lipid Peroxidation and the Antioxidant System in Human Erythrocytes In Vitro. Sci. World J. 2014, 2014, 874824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.; Zhou, Y.; Lyons, K.E.; Pahwa, R.; Reddy, M.B. Green Tea Consumption Reduces Oxidative Stress in Parkinson’s Disease Patients. J. Behav. Brain Sci. 2015, 5, 194–202. [Google Scholar] [CrossRef] [Green Version]
- Omotoso, G.O.; Gbadamosi, I.T.; Olajide, O.J.; Dada-Habeeb, S.O.; Arogundade, T.T.; Yawson, E.O. Moringa oleifera phytochemicals protect the brain against experimental nicotine-induced neurobehavioral disturbances and cerebellar degeneration. Pathophysiology 2018, 25, 57–62. [Google Scholar] [CrossRef]
- Muhammed, R.E.; El-Desouky, M.A.; Abo-Seda, S.B.; Nahas, A.; Elhakim, H.K.; Alkhalaf, M.I. The protecting role of Moringa oleifera in cypermethrin-induced mitochondrial dysfunction and apoptotic events in rats brain. J. King Saud Univ. Sci. 2020, 32, 2717–2722. [Google Scholar] [CrossRef]
- Kwiecien, S.; Jasnos, K.; Magierowski, M.; Sliwowski, Z.; Pajdo, R.; Brzozowski, B.; Mach, T.; Wojcik, D.; Brzozowski, T. Lipid peroxidation, reactive oxygen species and antioxidative factors in the pathogenesis of gastric mucosal lesions and mechanism of protection against oxidative stress-induced gastric injury. J. Physiol. Pharmacol. 2014, 65, 613–622. [Google Scholar]
- Offor, U.; Naidu, E.C.; Ogedengbe, O.O.; Jegede, A.I.; Peter, A.I.; Azu, O.O. Nephrotoxicity and highly active antiretroviral therapy: Mitigating action of Momordica charantia. Toxicol Rep. 2018, 5, 1153–1160. [Google Scholar] [CrossRef]
- Soliman, M.M.; Aldhahrani, A.; Alkhedaide, A.; Nassan, M.A.; Althobaiti, F.; Mohamed, W.A. The ameliorative impacts of Moringa oleifera leaf extract against oxidative stress and methotrexate-induced hepato-renal dysfunction. Biomed. Pharmacother. 2020, 128, 110259. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Sun, L.; Chen, X.; Zhang, D. Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural Regen. Res. 2013, 8, 2003–2014. [Google Scholar] [CrossRef]
- Bhatti, J.S.; Bhatti, G.K.; Reddy, P.H. Mitochondrial dysfunction and oxidative stress in metabolic disorders—A step towards mitochondria based therapeutic strategies. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2017, 1863, 1066–1077. [Google Scholar] [CrossRef] [PubMed]
- Mccomsey, G.A.; Daar, E.S.; O’Riordan, M.; Collier, A.C.; Kosmiski, L.; Santana, J.L.; Fichtenbaum, C.J.; Fink, H.; Sax, P.E.; Libutti, D.E.; et al. Changes in Fat Mitochondrial DNA and Function in Subjects Randomized to Abacavir-Lamivudine or Tenofovir DF–Emtricitabine with Atazanavir-Ritonavir or Efavirenz: AIDS Clinical Trials Group Study A5224s, Substudy of A5202. J. Infect. Dis. 2012, 207, 604–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olaniyan, L.W.B.; Maduagwu, E.N.; Akintunde, O.W.; Oluwayelu, O.O.; Brai, B.I.C. Open Access Macedonian Journal of Medical Sciences. Open Access Maced. J. Med Sci. 2015, 3, 545–550. [Google Scholar] [CrossRef] [Green Version]
- Funes, H.A.; Blas-Garcia, A.; Esplugues, J.V.; Apostolova, N. Efavirenz alters mitochondrial respiratory function in cultured neuron and glial cell lines. J. Antimicrob. Chemother. 2015, 70, 2249–2254. [Google Scholar] [CrossRef] [Green Version]
- Apostolova, N.; Blas-Garcia, A.; Galindo, M.J.; Esplugues, J.V. Efavirenz: What is known about the cellular mechanisms responsible for its adverse effects. Eur. J. Pharmacol. 2017, 812, 163–173. [Google Scholar] [CrossRef]
- Cheney, L.; Barbaro, J.; Berman, J. Antiretroviral Drugs Impact Autophagy with Toxic Outcomes. Cells 2021, 10, 909. [Google Scholar] [CrossRef]
- González-Burgos, E.; Ureña-Vacas, I.; Sánchez, M.; Gómez-Serranillos, M. Nutritional Value of Moringa oleifera Lam. Leaf Powder Extracts and Their Neuroprotective Effects via Antioxidative and Mitochondrial Regulation. Nutrients 2021, 13, 2203. [Google Scholar] [CrossRef]
- Vomhof-DeKrey, E.E.; Picklo, M.J. The Nrf2-antioxidant response element pathway: A target for regulating energy metabolism. J. Nutr. Biochem. 2012, 23, 1201–1206. [Google Scholar] [CrossRef]
- Claudia Tonelli, I.I.C.C.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxid. Redox Signal. 2018, 29, 1727–1745. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Et Biophys. Acta (BBA)-Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Marcus, J.M.; Andrabi, S.A. SIRT3 Regulation Under Cellular Stress: Making Sense of the Ups and Downs. Front. Neurosci. 2018, 12, 799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, D.; Siegel, D. Functions of NQO1 in Cellular Protection and CoQ10 Metabolism and its Potential Role as a Redox Sensitive Molecular Switch. Front. Physiol. 2017, 8, 595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sibiya, T.; Ghazi, T.; Mohan, J.; Nagiah, S.; Chuturgoon, A.A. Spirulina platensis Ameliorates Oxidative Stress Associated with Antiretroviral Drugs in HepG2 Cells. Plants 2022, 11, 3143. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.V.; Kotla, S.; Le, N.-T.; Ko, K.A.; Heo, K.-S.; Wang, Y.; Fujii, Y.; Vu, H.T.; McBeath, E.; Thomas, T.N.; et al. Senescent Phenotype Induced by p90RSK-NRF2 Signaling Sensitizes Monocytes and Macrophages to Oxidative Stress in HIV-Positive Individuals. Circulation 2019, 139, 1199–1216. [Google Scholar] [CrossRef]
- Kang, C.; Ji, L.L. Role of PGC-1α signaling in skeletal muscle health and disease. Ann. N. Y. Acad. Sci. 2012, 1271, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Rius-Pérez, S.; Torres-Cuevas, I.; Millán, I.; Ortega, A.L.; Pérez, S. PGC-1α, Inflammation, and Oxidative Stress: An Integrative View in Metabolism. Oxid. Med. Cell. Longev. 2020, 2020, 1452696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Ren, X.; Zhang, Q.; Li, Z.; Ma, S.; Bao, J.; Li, Z.; Bai, X.; Zheng, L.; Zhang, Z.; et al. PGC-1α/ERRα-Sirt3 Pathway Regulates DAergic Neuronal Death by Directly Deacetylating SOD2 and ATP Synthase β. Antioxid. Redox Signal. 2016, 24, 312–328. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.; Wang, R.; Xue, Y.; Liu, X.; Zhang, H.; Chen, Y.; Fang, F.; Chang, Y. Sirtuin 3, a New Target of PGC-1α, Plays an Important Role in the Suppression of ROS and Mitochondrial Biogenesis. PLoS ONE 2010, 5, e11707. [Google Scholar] [CrossRef] [Green Version]
- Elzohary, L.; Weglicki, W.B.; Chmielinska, J.J.; Kramer, J.H.; Mak, I.T. Mg-supplementation attenuated lipogenic and oxidative/nitrosative gene expression caused by Combination Antiretroviral Therapy (cART) in HIV-1-transgenic rats. PLOS ONE 2019, 14, e0210107. [Google Scholar] [CrossRef] [Green Version]
- Cheng, D.; Gao, L.; Su, S.; Sargsyan, D.; Wu, R.; Raskin, I.; Kong, A.-N. Moringa Isothiocyanate Activates Nrf2: Potential Role in Diabetic Nephropathy. AAPS J. 2019, 21, 31. [Google Scholar] [CrossRef] [PubMed]
- Sailaja, B.S.; Aita, R.; Maledatu, S.; Ribnicky, D.; Verzi, M.P.; Raskin, I. Moringa isothiocyanate-1 regulates Nrf2 and NF-κB pathway in response to LPS-driven sepsis and inflammation. PLoS ONE 2021, 16, e0248691. [Google Scholar] [CrossRef] [PubMed]
- Duranti, G.; Maldini, M.; Crognale, D.; Sabatini, S.; Corana, F.; Horner, K.; Ceci, R. Moringa oleifera leaf extract influences oxidative metabolism in C2C12 myotubes through SIRT1-PPARα pathway. Phytomedicine Plus 2020, 1, 100014. [Google Scholar] [CrossRef]
- Duranti, G.; Maldini, M.; Crognale, D.; Horner, K.; Dimauro, I.; Sabatini, S.; Ceci, R. Moringa oleifera Leaf Extract Upregulates Nrf2/HO-1 Expression and Ameliorates Redox Status in C2C12 Skeletal Muscle Cells. Molecules 2021, 26, 5041. [Google Scholar] [CrossRef]
- Wen, Y.; Liu, Y.; Huang, Q.; Liu, R.; Liu, J.; Zhang, F.; Liu, S.; Jiang, Y. Moringa oleifera Lam. seed extract protects kidney function in rats with diabetic nephropathy by increasing GSK-3β activity and activating the Nrf2/HO-1 pathway. Phytomedicine 2022, 95, 153856. [Google Scholar] [CrossRef]
- Abdou, K.H.; Moselhy, W.A.; Mohamed, H.M.; El-Nahass, E.-S.; Khalifa, A.G. Moringa oleifera Leaves Extract Protects Titanium Dioxide Nanoparticles-Induced Nephrotoxicity via Nrf2/HO-1 Signaling and Amelioration of Oxidative Stress. Biol. Trace Elem. Res. 2019, 187, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Mundkar, M.; Bijalwan, A.; Soni, D.; Kumar, P. Neuroprotective potential of Moringa oleifera mediated by NF-kB/Nrf2/HO-1 signaling pathway: A review. J. Food Biochem. 2022, 46, e14451. [Google Scholar] [CrossRef]
- Kim, C.G.; Chang, S.N.; Park, S.M.; Hwang, B.S.; Kang, S.-A.; Kim, K.S.; Park, J.G. Moringa oleifera mitigates ethanol-induced oxidative stress, fatty degeneration and hepatic steatosis by promoting Nrf2 in mice. Phytomedicine 2022, 100, 154037. [Google Scholar] [CrossRef]
- Rani, N.Z.A.; Husain, K.; Kumolosasi, E. Moringa Genus: A Review of Phytochemistry and Pharmacology. Front. Pharmacol. 2018, 9, 108. [Google Scholar] [CrossRef] [Green Version]
- Guerrero-Beltrán, C.E.; Calderón-Oliver, M.; Pedraza-Chaverri, J.; Chirino, Y.I. Protective effect of sulforaphane against oxidative stress: Recent advances. Exp. Toxicol. Pathol. 2012, 64, 503–508. [Google Scholar] [CrossRef]
- Kubo, E.; Chhunchha, B.; Singh, P.; Sasaki, H.; Singh, D.P. Sulforaphane reactivates cellular antioxidant defense by inducing Nrf2/ARE/Prdx6 activity during aging and oxidative stress. Sci. Rep. 2017, 7, 14130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiloke, C.; Phulukdaree, A.; Gengan, R.M.; Chuturgoon, A.A. Moringa oleifera Aqueous Leaf Extract Induces Cell-Cycle Arrest and Apoptosis in Human Liver Hepatocellular Carcinoma Cells. Nutr. Cancer 2019, 71, 1165–1174. [Google Scholar] [CrossRef] [PubMed]
- Kirindage, K.G.I.S.; Shanura Fernando, I.P.; Kumari Jayasinghe, A.M.; Han, E.-J.; Madhawa Dias, M.K.H.; Kang, K.-P.; Moon, S.-I. Moringa oleifera Hot Water Extract Protects Vero Cells from Hydrogen Peroxide-Induced Oxidative Stress by Regulating Mitochondria-Mediated Apoptotic Pathway and Nrf2/HO-1 Signaling. Foods 2022, 11, 420. [Google Scholar] [CrossRef]
- Matshediso, P.G.; Cukrowska, E.; Chimuka, L. Development of pressurised hot water extraction (PHWE) for essential compounds from Moringa oleifera leaf extracts. Food Chem. 2015, 172, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yin, Q.; Yang, Y. Comprehensive Investigation of Moringa oleifera from Different Regions by Simultaneous Determination of 11 Polyphenols Using UPLC-ESI-MS/MS. Molecules 2020, 25, 676. [Google Scholar] [CrossRef] [Green Version]
- Maiyo, C.; Moodley, F.R.; Singh, M. Cytotoxicity, antioxidant and apoptosis studies of quercetin-3-O glucoside and 4-(β-D-glucopyranosyl-1→ 4-α-L-rhamnopyranosyloxy)-benzyl isothiocyanate from Moringa oleifera. Anti-Cancer Agents Med. Chem. 2016, 16, 648–656. [Google Scholar] [CrossRef]
- Muzumbukilwa, W.T.; Nlooto, M.; Owira, P.M.O. Hepatoprotective effects of Moringa oleifera Lam (Moringaceae) leaf extracts in streptozotocin-induced diabetes in rats. J. Funct. Foods 2019, 57, 75–82. [Google Scholar] [CrossRef]
- Elshoff, J.-P.; Braun, M.; Andreas, J.-O.; Middle, M.; Cawello, W. Steady-State Plasma Concentration Profile of Transdermal Rotigotine: An Integrated Analysis of Three, Open-Label, Randomized, Phase I Multiple Dose Studies. Clin. Ther. 2012, 34, 966–978. [Google Scholar] [CrossRef]
- Thabethe, K.R.; Adefolaju, G.A.; Hosie, M.J. The effects of HAART on the expression of MUC1 and P65 in a cervical cancer cell line, HCS-2. Biomed. Pharmacother. 2015, 71, 227–232. [Google Scholar] [CrossRef]
- Gerber, I.B.; Dubery, I.A. Fluorescence microplate assay for the detection of oxidative burst products in tobacco cell suspensions using 2′, 7′-dichlorofluorescein. Meth. Cell Sci. 2004, 25, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Abdul, N.S.; Nagiah, S.; Chuturgoon, A.A. Fusaric acid induces mitochondrial stress in human hepatocellular carcinoma (HepG2) cells. Toxicon 2016, 119, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Weyermann, J.; Lochmann, D.; Zimmer, A. A practical note on the use of cytotoxicity assays. Int. J. Pharm. 2005, 288, 369–376. [Google Scholar] [CrossRef]
- Cree, I.; Andreotti, P. Measurement of cytotoxicity by ATP-based luminescence assay in primary cell cultures and cell lines. Toxicol. Vitr. 1997, 11, 553–556. [Google Scholar] [CrossRef]
- Arumugam, T.; Pillay, Y.; Ghazi, T.; Nagiah, S.; Abdul, N.S.; Chuturgoon, A.A. Fumonisin B1-induced oxidative stress triggers Nrf2-mediated antioxidant response in human hepatocellular carcinoma (HepG2) cells. Mycotoxin Res. 2018, 35, 99–109. [Google Scholar] [CrossRef]
- Ndlovu, S.; Nagiah, S.; Abdul, N.S.; Ghazi, T.; Chuturgoon, A.A. Deoxynivalenol downregulates NRF2-induced cytoprotective response in human hepatocellular carcinoma (HepG2) cells. Toxicon 2021, 193, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, T.; Yang, P.C. Western blot: Technique, theory, and trouble shooting. N. Am. J. Med. Sci. 2012, 4, 429–434. [Google Scholar]
- Patra, U.; Mukhopadhyay, U.; Mukherjee, A.; Sarkar, R.; Chawla-Sarkar, M. Progressive Rotavirus Infection Downregulates Redox-Sensitive Transcription Factor Nrf2 and Nrf2-Driven Transcription Units. Oxid Med Cell Longev. 2020, 2020, 7289120. [Google Scholar] [CrossRef]
- Čipak Gašparović, A.; Milković, L.; Dandachi, N.; Stanzer, S.; Pezdirc, I.; Vrančić, J.; Šitić, S.; Suppan, C.; Balic, M. Chronic Oxidative Stress Promotes Molecular Changes Associated with Epithelial Mesenchymal Transition, NRF2, and Breast Cancer Stem Cell Phenotype. Antioxidants 2019, 8, 633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul, N.S.; Nagiah, S.; Chuturgoon, A.A. Fusaric acid induces NRF2 as a cytoprotective response to prevent NLRP3 activation in the liver derived HepG2 cell line. Toxicol. Vitr. 2018, 55, 151–159. [Google Scholar] [CrossRef]
- Mohan, J.; Abdul, N.S.; Nagiah, S.; Ghazi, T.; Chuturgoon, A.A. Fumonisin B2 Induces Mitochondrial Stress and Mitophagy in Human Embryonic Kidney (Hek293) Cells—A Preliminary Study. Toxins 2022, 14, 171. [Google Scholar] [CrossRef] [PubMed]
- Chuturgoon, A.A.; Phulukdaree, A.; Moodley, D. Fumonisin B1 modulates expression of human cytochrome P450 1b1 in human hepatoma (Hepg2) cells by repressing Mir-27b. Toxicol. Lett. 2014, 227, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Catalog Number | Reference |
|---|---|---|
| pNRF2 | Abcam, (ab76026) Rabbit mAb | [123] |
| NRF2 | Cell signaling, Rabbit mAb #12721 | [124,125] |
| SOD2 | Cell signaling, Rabbit mAb #13194 | [125] |
| CAT | Cell signaling, Rabbit mAb #14097 | [121] |
| Sirt3 | Cell signaling, Rabbit mAb #5490 | [126] |
| Gene | Primer Sequences | Annealing Temperature (°C) |
|---|---|---|
| NRF2 | Sense 5′CACATCCAGTCAGAAACCAGTGG3′ Antisense 5′GGAATGTCTGCGCCAAAAGCTG3′ | 60 |
| SOD2 | Sense 5′CTGGACAAACCTCAGCCCTAAC3′ Antisense 5′AACCTGAGCCTTGGACACCAAC3′ | 57 |
| Sirt3 | Sense 5′CCCTGGAAACTACAAGCCCAAC3′ Antisense 5′GCAGAGGCAAAGGTTCCATGAG3′ | 58 |
| PGC1α | Sense 5′CCAAAGGATGCGCTCTCGTTCA3′ Antisense 5′CGGTGTCTGTAGTGGCTTGACT3′ | 62 |
| NQO1 | Sense 5′CCTGCCATTCTGAAAGGCTGGT3′ Antisense 5′GTGGTGATGGAAAGCACTGCCT3′ | 58 |
| CAT | Sense 5′GTGCGGAGATTCAACACTGCCA3′ Antisense 5′CGGCAATGTTCTCACACAGACG3′ | 60 |
| UCP2 | Sense 5′TGGTCGGAGATACCAAAGCACC3′ Antisense 5′GCTCAGCACAGTTGACAATGGC3′ | 59 |
| GAPDH | Sense 5′CACCATTGGCAATGAGCGGTTC3′ Antisense 5′AGGTCTTTGCGGATGTCCACGT3′ | Variable |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ndlovu, S.S.; Chuturgoon, A.A.; Ghazi, T. Moringa oleifera Lam Leaf Extract Stimulates NRF2 and Attenuates ARV-Induced Toxicity in Human Liver Cells (HepG2). Plants 2023, 12, 1541. https://doi.org/10.3390/plants12071541
Ndlovu SS, Chuturgoon AA, Ghazi T. Moringa oleifera Lam Leaf Extract Stimulates NRF2 and Attenuates ARV-Induced Toxicity in Human Liver Cells (HepG2). Plants. 2023; 12(7):1541. https://doi.org/10.3390/plants12071541
Chicago/Turabian StyleNdlovu, Siqiniseko S., Anil A. Chuturgoon, and Terisha Ghazi. 2023. "Moringa oleifera Lam Leaf Extract Stimulates NRF2 and Attenuates ARV-Induced Toxicity in Human Liver Cells (HepG2)" Plants 12, no. 7: 1541. https://doi.org/10.3390/plants12071541
APA StyleNdlovu, S. S., Chuturgoon, A. A., & Ghazi, T. (2023). Moringa oleifera Lam Leaf Extract Stimulates NRF2 and Attenuates ARV-Induced Toxicity in Human Liver Cells (HepG2). Plants, 12(7), 1541. https://doi.org/10.3390/plants12071541