
Phenotypic Evaluation of Soybean Genotypes for Their Reaction to a Mississippi Isolate of Phakopsora pachyrhizi Causing Soybean Rust

Abstract
:1. Introduction
2. Results
2.1. Reaction/Lesion Type
2.2. Rust Severity
2.3. Rust Sporulation
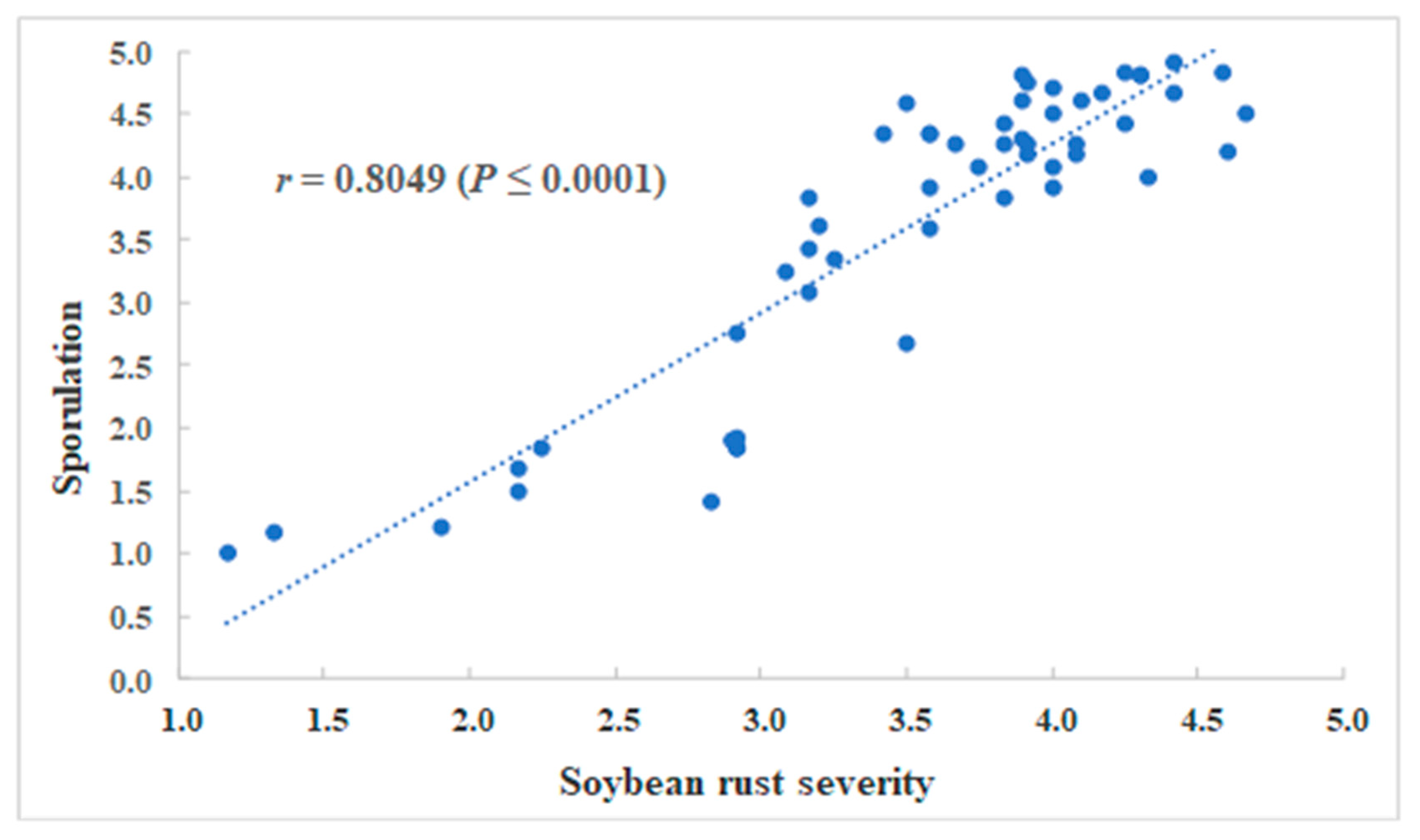
2.4. Correlation Analysis between Soybean Rust Severity and Sporulation
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Breeding Line Development
4.2. Pathogen Isolate, Purification, and Maintenance
4.3. Inoculum Preparation and Plant Inoculation
4.4. Assessment of Lesion Types, Rust Severity, and Sporulation
4.5. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hartman, G.L.; Sikora, E.J.; Rupe, J.C. Rust. In Compendium of Soybean Diseases and Pets, 5th ed.; Hartman, G.L., Rupe, J.C., Sikora, E.J., Domier, L.L., Davis, J.A., Steffey, K.L., Eds.; APS Press: Saint Paul, MN, USA, 2015; pp. 56–69. [Google Scholar]
- Godoy, C.V.; Seixas, C.D.S.; Soares, R.M.; Marcelino-Guimaraes, F.C.; Meyer, M.C.; Costamilan, L.M. Asian soybean rust in Brazil: Past, present, and future. Pesq. Agropec. Bras. 2016, 51, 407–421. [Google Scholar] [CrossRef]
- Kumudini, S.; Godoy, C.V.; Board, J.E.; Omielan, J.; Tollenar, M. Mechanisms involved in soybean rust-inducedyield reduction. Crop Sci. 2008, 48, 2334–2342. [Google Scholar] [CrossRef]
- Ishiwata, Y.I.; Furuya, J. Evaluating the contribution of soybean rust–resistant cultivars to soybean production and the soybean market in Brazil: A supply and demand model analysis. Sustainability 2020, 12, 1422. [Google Scholar] [CrossRef]
- Bromfield, K.R. Soybean Rust. Mongr; No. 11; American Phytopathlogical Society Press: Saint Paul, MN, USA, 1984. [Google Scholar]
- Hennings, P. Some new Japanese Uredinale. IV. Hedwigia Suppl. 1903, 42, 107–108. [Google Scholar]
- Sydow, H.; Sydow, P. A contribution to knowledge of the parasitic fungi on the island of Formosa. Ann. Mycol. 1914, 12, 105. [Google Scholar]
- Goellner, K.; Loehrer, M.; Langenbach, C.; Conrath, U.; Koch, E.; Schaffrath, U. Phakopsora pachyrhizi, the causal agent of Asian soybean rust. Mol. Plant Pathol. 2010, 11, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Killgore, E.; Heu, R.; Gardner, D.E. First report of soybean rust in Hawaii. Plant Dis. 1994, 78, 1216. [Google Scholar] [CrossRef]
- Schneider, R.W.; Hollier, C.A.; Whitam, H.K.; Palm, M.E.; McKemy, J.M.; Hernández, J.R.; Levy, L.; DeVries-Paterson, R. First report of soybean rust caused by Phakopsora pachyrhizi in the continental United States. Plant Dis. 2005, 89, 774. [Google Scholar] [CrossRef]
- Pham, T.A.; Miles, M.R.; Frederick, R.D.; Hill, C.B.; Hartman, G.L. Differential responses of resistant soybean entries to isolates of Phakopsora pachyrhizi. Plant Dis. 2009, 93, 224–228. [Google Scholar] [CrossRef]
- Miles, M.R.; Bonde, M.R.; Nester, S.E.; Berner, D.K.; Frederick, R.D.; Hartman, G.L. Characterizing resistance to Phakopsora pachyrhizi in soybean. Plant Dis. 2011, 95, 577–581. [Google Scholar] [CrossRef]
- Bonde, M.R.; Nester, S.E.; Austin, C.N.; Stone, C.L.; Frederick, R.D.; Hartman, G.L.; Miles, M.R. Evaluation of virulence of Phakopsora pachyrhizi and P. meibomiae isolates. Plant Dis. 2006, 90, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Bromfield, K.R.; Hartwig, E.E. Resistance to soybean rust and mode of inheritance. Crop Sci. 1980, 20, 254–255. [Google Scholar]
- Bromfield, K.R.; Melching, J.S.; Kingsolver, C.H. Virulence and aggressiveness of Phakopsora pachyrhizi isolates causing soybean rust. Phytopathology 1980, 70, 17–21. [Google Scholar] [CrossRef]
- Hartwig, E.E.; Bromfield, K.R. Relationships among three genes conferring specific resistance to rust in soybeans. Crop Sci. 1983, 23, 237–239. [Google Scholar] [CrossRef]
- Hartwig, E.E. Identification of a fourth major gene conferring resistance to soybean rust. Crop Sci. 1986, 26, 1135–1136. [Google Scholar] [CrossRef]
- Garcia, A.; Calvo, E.S.; de Souza Kiihl, R.A.; Harada, A.; Hiromoto, D.M.; Vieira, L.G. Molecular mapping of soybean rust (Phakopsora pachyrhizi) resistance genes: Discovery of a novel locus and alleles. Theor. Appl. Genet. 2008, 117, 545–553. [Google Scholar] [CrossRef]
- Li, S.; Smith, J.; Ray, J.; Frederick, D. Identification of a new soybean rust resistance gene in PI 567102B. Theor. Appl. Genet. 2012, 125, 133–142. [Google Scholar] [CrossRef]
- Liu, M.; Li, S.; Swaminathan, S.; Sahu, B.; Leandro, L.; Cardinal, A.; Bhattacharyya, M.; Song, Q.; Walker, D.; Cianzio, S. Identification of a soybean rust resistance gene in PI 567104B. Theor. Appl. Genet. 2016, 129, 863–877. [Google Scholar] [CrossRef]
- Childs, S.P.; King, Z.R.; Walker, D.R.; Harris, D.K.; Pedley, K.F.; Buck, J.W.; Boerma, H.R.; Li, Z. Discovery of a seventh Rpp soybean rust resistance locus in soybean accession PI 605823. Theor. Appl. Genet. 2018, 131, 27–41. [Google Scholar] [CrossRef]
- Kendrick, M.D.; Harris, D.K.; Ha, B.; Hyten, D.L.; Cregan, P.B.; Frederick, R.D.; Boerma, H.R.; Pedley, K.F. Identification of a second Asian soybean rust resistance gene in Hyuuga soybean. Phytopathology 2011, 101, 535–543. [Google Scholar] [CrossRef]
- Hartman, G.L.; Miles, M.R.; Frederick, R.D. Breeding for resistance to soybean rust. Plant Dis. 2005, 89, 664–666. [Google Scholar] [CrossRef] [PubMed]
- Li, S. Identification of new sources of resistance to soybean rust. In Plant Pathology in the 21st Century: Recent Development in Disease Management; Gullino, M.L., Gisi, U., Chet, I., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 1, pp. 357–373. [Google Scholar]
- Paul, C.; Motter, H.Z.; Walker, D.R. Reactions of soybean germplasm accessions to six Phakopsora pachyrhizi. isolates from the United States. Plant Dis. 2020, 104, 1087–1095. [Google Scholar] [CrossRef] [PubMed]
- Grieshop, C.M.; Fahey, G.C., Jr. Comparison of quality characteristics of soybeans from Brazil, China, and the United States. J. Agric. Food Chem. 2001, 49, 2669–2673. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.F. Seed composition. In Soybean: Improvement, Production, and Uses, 3rd ed.; Boerma, H.R., Specht, J.E., Eds.; American Society of Agronomy: Madison, WI, USA, 2004; pp. 621–677. [Google Scholar]
- Li, S.; Moore, W.F.; Spinks, B.L.; Wells, B.C.; Sciumbato, G.L.; Robinson, S.J.; Libous-Bailey, L. Occurrence of Asian soybean rust caused by Phakopsora pachyrhizi in Mississippi. Plant Health Prog. 2007, 8, 45. [Google Scholar] [CrossRef]
- Miles, M.R.; Frederick, R.D.; Hartman, G.L. Evaluation of soybean germplasm for resistance to Phakopsora pachyrhizi. Plant Health Progress. 2006, 7, 33. [Google Scholar] [CrossRef]
- Li, S. Reaction of soybean rust-resistant lines identified in Paraguay to Mississippi isolates of Phakopsora pachyrhizi. Crop Sci. 2009, 49, 887–894. [Google Scholar] [CrossRef]
- Miles, M.R.; Pastor-Corrales, M.A.; Hartman, G.; Frederick, R.D. Differential Response of common bean cultivars to Phakopsora pachyrhizi. Plant Dis. 2007, 91, 698–704. [Google Scholar] [CrossRef]
- Sinclair, J.B.; Hartman, G.L. Proceedings of the Soybean Rust Workshop. In Proceedings of the Soybean Rust Workshop Urbana-Champaign, Urbana, IL, USA, 9–11 August 1995. [Google Scholar]
- Sinclair, J.B.; Hartman, G.L. Soybean rust. In Compendium of Soybean Diseases; Hartman, G.L., Sinclair, J.B., Rupe, J.C., Eds.; American Phytopathlogical Society Press: Saint Paul, MN, USA, 1999; pp. 25–26. [Google Scholar]
- Yorinori, J.T.; Pavia, W.M.; Frederick, R.D.; Costamilan, L.M.; Bertagnolli, P.F.; Hartman, G.L.; Godoy, C.V.; Nunes, J., Jr. Epidemics of soybean rust (Phakopsora pachyrhizi) in Brazil and Paraguay from 2001 to 2003. Plant Dis. 2005, 89, 675–677. [Google Scholar] [CrossRef]
- Slaminko, T.L.; Miles, M.R.; Marois, J.J.; Wright, D.L.; Hartman, G.L. Hosts of Phakopsora pachyrhizi identified in field evaluations in Florida. Plant Health Prog. 2008, 9, 6. [Google Scholar] [CrossRef]
- Pan, Z.; Yang, X.B.; Pivonia, S.; Xue, L.; Pasken, R.; Roads, J. Long-term prediction of soybean rust entry into the continental United States. Plant Dis. 2006, 90, 840–846. [Google Scholar] [CrossRef]
- Twizeyimana, M.; Ojiambo, P.S.; Haudenshield, J.S.; Caetano-Anollés, G.; Pedley, K.F.; Bandyopadhayay, R.; Hartman, G.L. Genetic structure and diversity of Phakopsora pachyrhizi isolates from soyabean. Plant Pathol. 2011, 60, 719–729. [Google Scholar] [CrossRef]
- Darben, L.M.; Yokoyama, A.; Castanho, F.M.; Lopes-Caitar, V.S.; De Carvalho, M.C.; De Carvalho, M.C.C.G.; Gonela, A.; Marcelino-Guimarães, F.C. Characterization of genetic diversity and pathogenicity of Phakopsora pachyrhizi mono-uredinial isolates collected in Brazil. Eur. J. Plant Pathol. 2020, 156, 355–372. [Google Scholar] [CrossRef]
- Twizeyimana, M.; Hartman, G.L. Pathogenic variation of Phakopsora pachyrhizi isolates on soybean in the United States from 2006 to 2009. Plant Dis. 2012, 96, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Akamatsu, H.; Yamanaka, N.; Yamaoka, Y.; Soares, R.M.; Morel, W.; Ivanovich, A.J.G.; Bogado, A.N.; Kato, M.; Yorinori, J.T.; Suenaga, K. Pathogenic diversity of soybean rust in Argentina, Brazil, and Paraguay. J. Gen Plant Pathol. 2013, 79, 28–40. [Google Scholar] [CrossRef]
- García-Rodríguez, J.C.; Vicente-Hernández, Z.; Grajales-Solís, M.; Yamanaka, N. Virulence Diversity of Phakopsora pachyrhizi in Mexico. Phy. Front. 2022, 2, 52–59. [Google Scholar] [CrossRef]
- Isard, S.A.; Barnes, C.W.; Hambleton, S.; Ariatti, A.; Russo, J.M.; Tenuta, A.; Gay, D.A.; Szabo, L.J. Predicting soybean rust incursions into the North American continental interior using crop monitoring, spore trapping, and aerobiological modeling. Plant Dis. 2011, 95, 1346–1357. [Google Scholar] [CrossRef]
- Christiano, R.C.S.; Scherm, H. Quantitative aspects of the spread of Asian soybean rust in the southeastern United States, 2005 to 2006. Phytopathology 2007, 97, 1428–1433. [Google Scholar] [CrossRef]
- Walker, D.R.; Harris, D.K.; King, Z.R.; Li, Z.; Boerma, H.R.; Buckley, J.B.; Weaver, D.B.; Sikora, E.J.; Shipe, E.R.; Mueller, J.D.; et al. Evaluation of soybean germplasm accessions for resistance to Phakopsora pachyrhizi populations in the southeastern United States, 2009–2012. Crop Sci. 2014, 54, 1673–1689. [Google Scholar] [CrossRef]
- Li, S.; Young, L.D. Evaluation of selected genotypes of soybean for resistance to Phakopsora pachyrhizi. Plant Health Prog. 2009, 10, 15. [Google Scholar] [CrossRef]
- Miles, M.R.; Morel, W.; Ray, J.D.; Smith, J.R.; Frederick, R.D.; Hartman, G.L. Adult plant evaluation of soybean accessions for resistance to Phakopsora pachyrhizi in the field and greenhouse in Paraguay. Plant Dis. 2008, 92, 96–105. [Google Scholar] [CrossRef]
- Yamanaka, N.; Yamaoka, Y.; Kato, M.; Lemos, N.G.; Passionotto, A.L.L.; dos Santos, J.V.M.; Benitez, E.R.; Abdelnoor, R.V.; Soares, R.M.; Suenaga, K. Development of classification criteria for resistance to soybean rust and differences in virulence among Japanese and Brazilian rust populations. Trop. Plant Pathol. 2010, 35, 153–162. [Google Scholar] [CrossRef]
- Ray, J.D.; Morel, W.; Smith, J.R.; Frederick, R.D.; Miles, M.R. Genetics and mapping of adult plant rust resistance in soybean PI 587886 and PI 587880A. Theor. Appl. Genet. 2009, 119, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Stone, C.L.; Smith, J.R.; Ray, J.D.; Gillen, A.M.; Frederick, R.D. Phenotypic reactions of 53 soybean genotypes to infection with each of 16 isolates of Phakopsora pachyrhizi. J. Crop Improv. 2021, 36, 25–39. [Google Scholar] [CrossRef]
- Fehr, W.R.; Caviness, C.E. Stages of soybean development. Iowa Agricultural Experiment Station, Special Report 80; Iowa State University: Ames, IA, USA, 1977. [Google Scholar]



{kind=link}
{kind=link}
| Entry Code | Line Name | Genotype | MG a | Pedigree or Accession Name |
|---|---|---|---|---|
| SB01 | CM422 | Rpp4 | V | 5601T × L87-0482 (PI 547879) |
| SB02 | 4013-2-1-1-4-1 | Rpp2 &/or ? b | III | L86-1752 (PI 547878) × GC 00138-29 |
| SB03 | 4014-242-341 | Rpp2 &/or ? b | III | L86-1752 (PI 547878) × GC 84058-18-4 |
| SB04 | 6112-13-6211 | Rpp1d c | III | LG01-5087-101 × PI 587886 |
| SB05 | 6112-55-3-1-21 | Rpp1d c | IV | LG01-5087-101 × PI 587886 |
| SB06 | 6112-13-3-1-11 | Rpp1d c | III | LG01-5087-101 × PI 587886 |
| SB07 | RN6-313-362 | Rpp1b | IV | NE2701 × PI 594538A |
| SB08 | 4013-1-3-5-3-2 | Unknown | III | L86-1752 (PI 547878) × GC 00138-29 |
| SB09 | 4018-4-1-1-212 | Unknown | III | L87-0482 (PI 547879) × GC 84051-9-1 |
| SB10 | 4019-2-2-3-3-1 | Unknown | IV | L87-0482 (PI 547879) × GC 00138-29 |
| SB11 | 6112-13-6-42 | Rpp1d c | IV | LG01-5087-101 × PI 587886 |
| SB12 | 6112-13-33-1-1 | Rpp1d c | IV | LG01-5087-101 × PI 587886 |
| SB13 | 6112-136-411 | Rpp1d c | IV | LG01-5087-101 × PI 587886 |
| SB14 | RN6-3-134211 | Rpp1b | IV | NE2701 × PI 594538A |
| SB15 | 6108-1043321 | Rpp1c c | IV | JTN-5503 × PI 587880A |
| SB16 | 6116-5-4-1-1 | Unknown | IV | DS-880 × PI 605779E |
| SB17 | 7092-x-1 | rpp3 | IV | LG01-5087-9 × PI 567099A |
| SB18 | 88-203-1-2-1 | Unknown | V | DS-880 × PI 567145C |
| SB19 | 7088-198-212 | Unknown | V | DS-880 × PI 567145C |
| SB20 | 7088-198-521 | Unknown | V | DS-880 × PI 567145C |
| SB21 | 6108-1662111 | Rpp1c c | V | JTN-5503 × PI 587880A |
| SB22 | 6115-22-511 | Unknown | V | DS-880 × PI 594723 |
| SB23 | 6113-216-121 | Rpp1e c | VI | DS-880 × PI 587905 |
| SB24 | RN06-32-1(7-b | Unknown | V | Dillon × PI 605891A |
| SB25 | 6106-132-1-2 | Unknown | IV | DS-880 × PI 567102B |
| SB26 | DS5-67 | Rpp3 | V | Williams 82 × Ankur (PI 462312) |
| SB27 | G01-PR16 | Rpp3 + Rpp5 | V | Dillon × Hyuuga |
| SB28 | 4022-1-4-124 | Rpp4 | V | 5601T × L87-0482 (PI 547879) |
| SB29 | 4022-1-4-121 | Rpp4 | V | 5601T × L87-0482 (PI 547879) |
| SB30 | Williams 82 | Susceptible | III | Williams (7) × Kingwa |
| SB31 | PI 594538A | Rpp1b | VIII | ‘Min hou bai sha wan dou’ |
| SB32 | PI 200492 | Rpp1a | VII | Komata |
| SB33 | PI 587886 | Rpp1d c | VI | Bai dou |
| SB34 | PI 587880A | Rpp1c c | VI | Huang dou |
| SB36 | PI 587905 | Rpp1e c | VII | Xiao huang dou |
| SB37 | PI 567099A | rpp3 | VIII | MARIF 2740 |
| SB38 | PI 462312 | Rpp3 | VIII | Ankur |
| SB39 | GC00138-29 | Unknown | VI | Unknown |
| SB40 | GC84058-18-4 | Unknown | V | Unknown |
| SB41 | GC84051-9-1 | Unknown | IV | Unknown |
| SB42 | PI 567102B | Rpp6 | VII | MARIF 2767 |
| SB43 | PI 594723 | Unknown | VII | He xian hei dou |
| SB44 | PI 605779E | Unknown | VIII | Sample 42 |
| SB45 | PI 230970 | Rpp2 | VII | No. 3 |
| SB46 | PI 459025B | Rpp4 | VIII | Bing nan |
| SB47 | PI 594760B | Unknown | VIII | ‘Gou jiao huang dou’ |
| SB48 | PI 567145C | Unknown | VIII | MARIF 2816 |
| SB49 | Stafford | Susceptible | IV | V66-318 × V68-2331 |
| SB50 | 5002T | Susceptible | V | Holladay × Manokin |
| SB51 | PI 605833 | Unknown | IX | Sample 102 |
| SB52 | Osage | Susceptible | V | Hartz 5545 × KS 4895 |
| SB54 | NC-Roy | Susceptible | VI | Holladay × Brim |
| SB55 | PI 200456 | rpp5 | VIII | Awashima Zairai |
| SB56 | PI 200487 | Rpp5 | VIII | Kinoshita |
| Entry Code | Line Name | Genotype | MG a | Lesion Type b | Severity c | Sporulation d | ||
|---|---|---|---|---|---|---|---|---|
| SB42 | PI 567102B e | Rpp6 | VII | NIM | 1.2 | p f | 1.0 | r f |
| SB01 | CM422 | Rpp4 | V | RB | 3.2 | lmn | 3.4 | klm |
| SB02 | 4013-2-1-1-4-1 | Rpp2 &/or ? g | III | RB | 2.9 | n | 1.9 | o |
| SB03 | 4014-242-341 | Rpp2 &/or ? g | III | RB | 3.6 | hijkl | 3.6 | jkl |
| SB17 | 7092-x-1 | rpp3 | IV | RB | 2.9 | n | 1.8 | op |
| SB24 | RN06-32-1(7-b) | Unknown | V | RB | 2.2 | o | 1.7 | op |
| SB26 | DS5-67 | Rpp3 | V | RB | 2.9 | n | 1.8 | op |
| SB27 | G01-PR16 | Rpp3 + Rpp5 | V | RB | 2.8 | n | 1.4 | pqr |
| SB28 | 4022-1-4-124 | Rpp4 | V | RB | 3.5 | ijklm | 2.7 | n |
| SB29 | 4022-1-4-121 | Rpp4 | V | RB | 3.2 | klmn | 3.6 | jkl |
| SB32 | PI 200492 e | Rpp1a | VII | RB | 1.3 | p | 1.2 | qr |
| SB38 | PI 462312 e | Rpp3 | VIII | RB | 2.9 | n | 1.9 | o |
| SB39 | GC00138-29 | Unknown | VI | RB | 2.2 | o | 1.5 | opq |
| SB40 | GC84058-18-4 | Unknown | V | RB | 3.2 | lmn | 3.1 | mn |
| SB41 | GC84051-9-1 | Unknown | IV | RB | 2.3 | o | 1.8 | op |
| SB45 | PI 230970 e | Rpp2 | VII | RB | 2.9 | n | 2.8 | n |
| SB46 | PI 459025B e | Rpp4 | VIII | RB | 3.3 | klmn | 3.3 | lm |
| SB56 | PI 200487 e | Rpp5 | VIII | RB | 1.9 | o | 1.2 | qr |
| SB04 | 6112-13-6211 | Rpp1d h | III | TAN | 4.0 | cdefgh | 4.1 | fghi |
| SB05 | 6112-55-3-1-21 | Rpp1d h | IV | TAN | 4.0 | cdefgh | 3.9 | hij |
| SB06 | 6112-13-3-1-11 | Rpp1d h | III | TAN | 3.6 | hijkl | 4.3 | cdefgh |
| SB07 | RN6-313-362 | Rpp1b | IV | TAN | 4.1 | cdefg | 4.3 | defghi |
| SB08 | 4013-1-3-5-3-2 | Unknown | III | TAN | 3.6 | hijkl | 3.9 | hij |
| SB09 | 4018-4-1-1-212 | Unknown | III | TAN | 3.8 | efghij | 4.3 | defghi |
| SB10 | 4019-2-2-3-3-1 | Unknown | IV | TAN | 3.9 | defghij | 4.6 | abcde |
| SB11 | 6112-13-6-42 | Rpp1d h | IV | TAN | 3.4 | jklm | 4.3 | cdefgh |
| SB12 | 6112-13-33-1-1 | Rpp1d h | IV | TAN | 3.6 | hijkl | 4.3 | cdefgh |
| SB13 | 6112-136-411 | Rpp1d h | IV | TAN | 4.1 | bcdefg | 4.6 | abcde |
| SB14 | RN6-3-134211 | Rpp1b | IV | TAN | 4.3 | abcde | 4.4 | cdefg |
| SB15 | 6108-1043321 | Rpp1c h | IV | TAN | 4.4 | abc | 4.9 | a |
| SB16 | 6116-5-4-1-1 | Unknown | IV | TAN | 4.1 | cdefg | 4.2 | efghi |
| SB18 | 88-203-1-2-1 | Unknown | V | TAN | 4.3 | abcde | 4.8 | ab |
| SB19 | 7088-198-212 | Unknown | V | TAN | 3.9 | defghi | 4.3 | defghi |
| SB20 | 7088-198-521 | Unknown | V | TAN | 3.8 | efghij | 4.4 | cdefg |
| SB21 | 6108-1662111 | Rpp1c h | V | TAN | 4.0 | cdefgh | 4.7 | abcd |
| SB22 | 6115-22-511 | Unknown | V | TAN | 3.5 | ijklm | 4.6 | abcde |
| SB23 | 6113-216-121 | Rpp1e h | VI | TAN | 3.9 | defghij | 4.8 | ab |
| SB25 | 6106-132-1-2 | Unknown | IV | TAN | 3.9 | defghi | 4.2 | efghi |
| SB30 | Williams 82 i | Susceptible | III | TAN | 4.6 | ab | 4.8 | ab |
| SB31 | PI 594538A | Rpp1b | VIII | TAN | 4.3 | abcde | 4.8 | ab |
| SB33 | PI 587886 | Rpp1d h | VI | TAN | 4.3 | abcd | 4.0 | ghij |
| SB34 | PI 587880A | Rpp1c h | VI | TAN | 3.8 | fghij | 4.1 | fghi |
| SB36 | PI 587905 | Rpp1e h | VII | TAN | 4.7 | a | 4.5 | abcdef |
| SB37 | PI 567099A | rpp3 | VIII | TAN | 3.1 | mn | 3.3 | lm |
| SB43 | PI 594723 | Unknown | VII | TAN | 3.2 | lmn | 3.8 | ijk |
| SB44 | PI 605779E | Unknown | VIII | TAN | 3.8 | efghij | 3.8 | ijk |
| SB47 | PI 594760B | Unknown | VIII | TAN | 4.2 | bcdef | 4.7 | abcd |
| SB48 | PI 567145C | Unknown | VIII | TAN | 4.4 | abc | 4.7 | abcd |
| SB49 | Stafford i | Susceptible | IV | TAN | 3.9 | defghij | 4.3 | cdefgh |
| SB50 | 5002T i | Susceptible | V | TAN | 4.3 | abcde | 4.8 | ab |
| SB51 | PI 605833 | Unknown | IX | TAN | 3.9 | defghi | 4.8 | abc |
| SB52 | Osage i | Susceptible | V | TAN | 3.7 | ghijk | 4.3 | defghi |
| SB54 | NC-Roy i | Susceptible | VI | TAN | 4.6 | ab | 4.2 | efghi |
| SB55 | PI 200456 | rpp5 | VIII | TAN | 4.0 | cdefgh | 4.5 | abcdef |
| Mean | 3.5 | 3.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Smith, J.R. Phenotypic Evaluation of Soybean Genotypes for Their Reaction to a Mississippi Isolate of Phakopsora pachyrhizi Causing Soybean Rust. Plants 2023, 12, 1797. https://doi.org/10.3390/plants12091797
Li S, Smith JR. Phenotypic Evaluation of Soybean Genotypes for Their Reaction to a Mississippi Isolate of Phakopsora pachyrhizi Causing Soybean Rust. Plants. 2023; 12(9):1797. https://doi.org/10.3390/plants12091797
Chicago/Turabian StyleLi, Shuxian, and James R. Smith. 2023. "Phenotypic Evaluation of Soybean Genotypes for Their Reaction to a Mississippi Isolate of Phakopsora pachyrhizi Causing Soybean Rust" Plants 12, no. 9: 1797. https://doi.org/10.3390/plants12091797
APA StyleLi, S., & Smith, J. R. (2023). Phenotypic Evaluation of Soybean Genotypes for Their Reaction to a Mississippi Isolate of Phakopsora pachyrhizi Causing Soybean Rust. Plants, 12(9), 1797. https://doi.org/10.3390/plants12091797