
Regulation of NADP-Malic Enzyme Activity in Maize (Zea mays L.) under Salinity with Reference to Light and Darkness
 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Seedling Development
2.2. Salt Treatment
2.3. Induction of Enzyme Activity by Light (L)/Darkness (D) and Bicarbonate Treatments
2.4. Extraction and Purification of NADP-ME Enzyme
2.5. Assay Buffer and Reaction for NADP-ME Activity In Vitro
2.6. Separation and Staining of Polymorphic Isoenzyme Bands of NADP-ME
2.7. Estimation of Chlorophyll Content and Determination of Hill Activity
2.8. Determination of Reducing Sugar and Total Sugar Content
2.9. Quantification of Proline Content
2.10. Determination of Na+ and K+
2.11. Determination of Malic Acid and Pyruvic Acid Content through High-Performance Liquid Chromatography (HPLC)
2.12. Statistical Analysis
3. Results
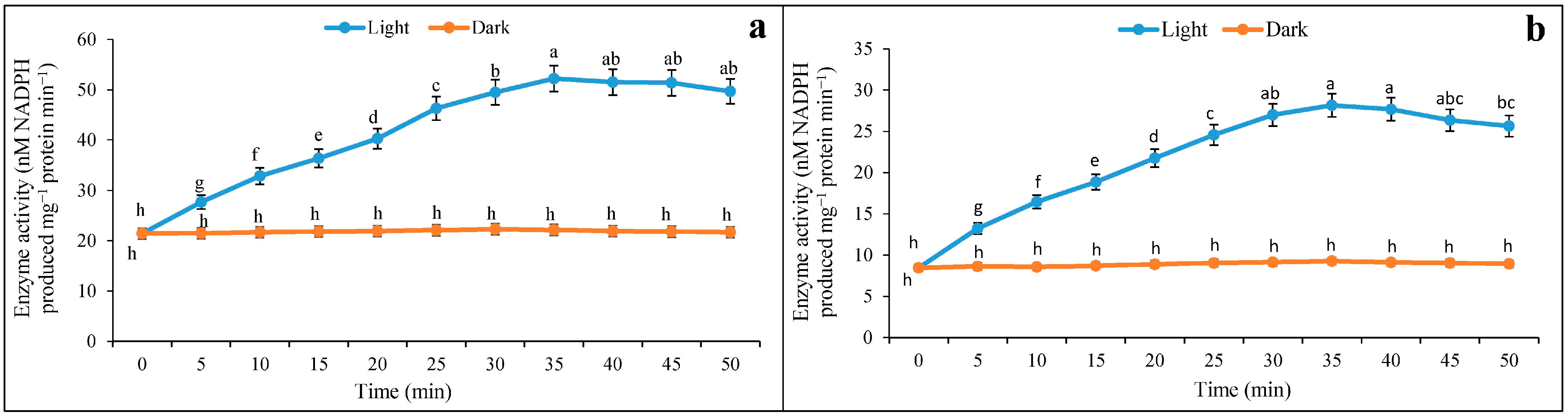
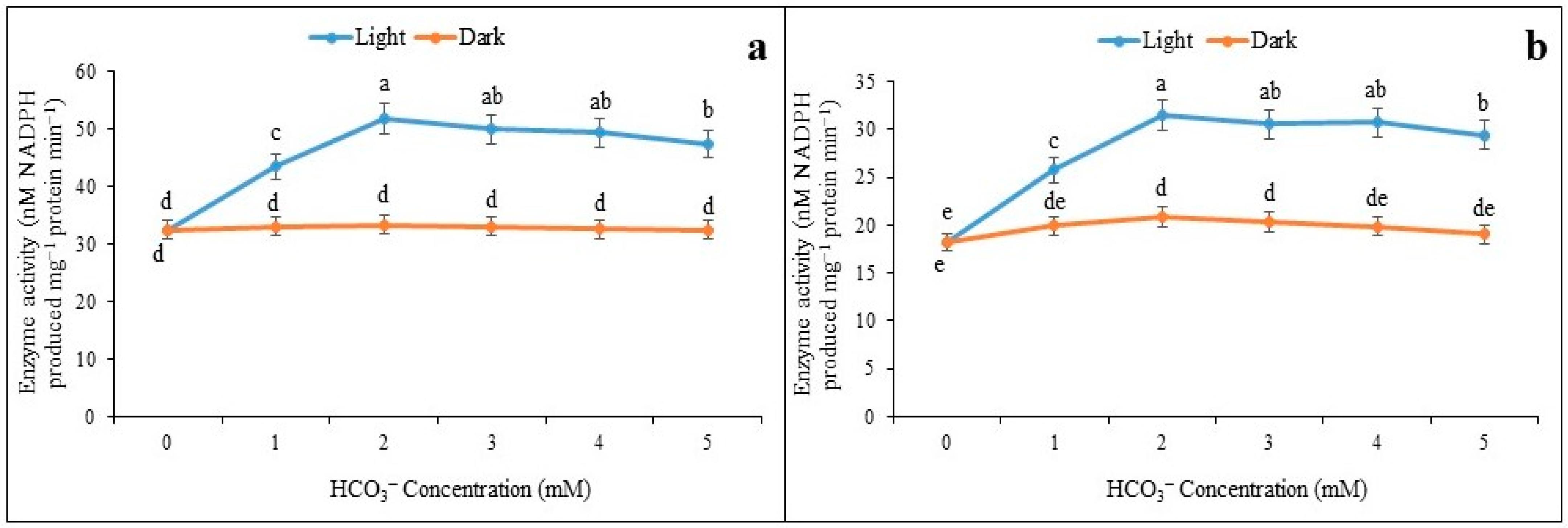
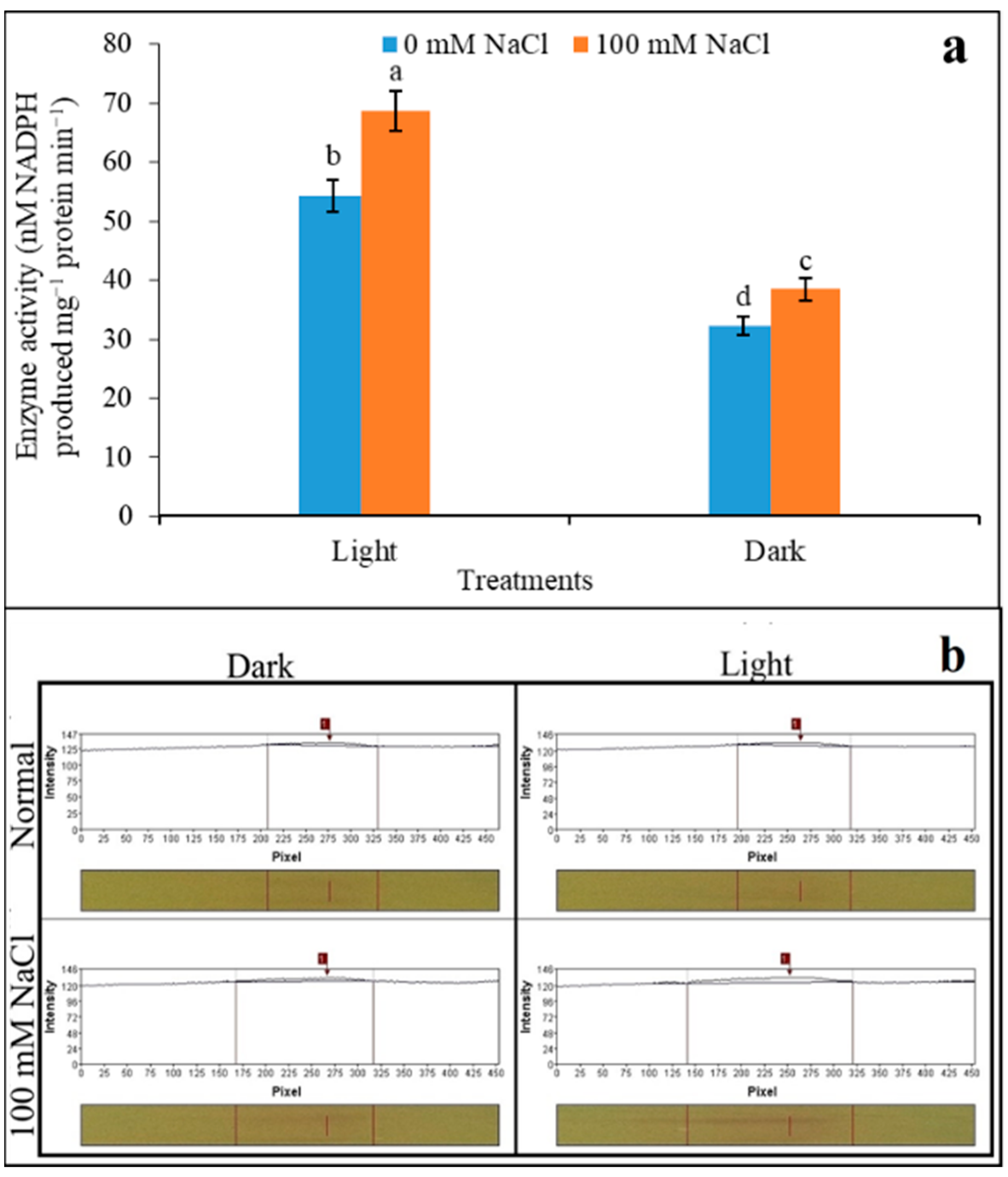
3.1. Inductions of NADP-ME under Light and Darkness
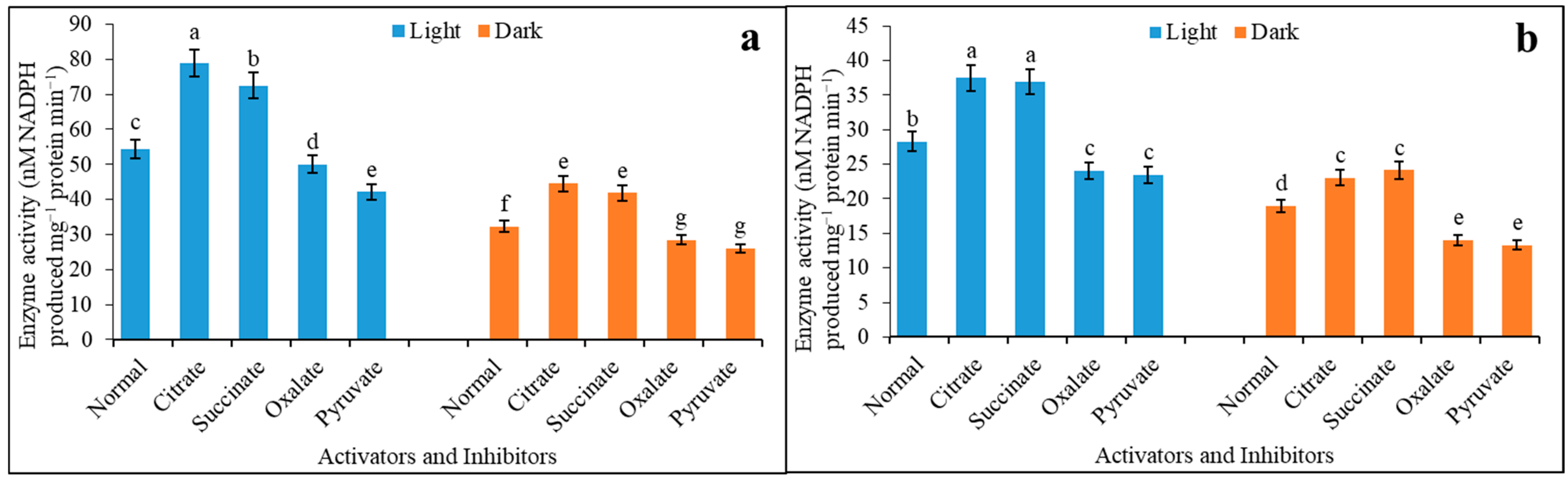
3.2. Activators and Inhibitors Affect the Regulation of Enzyme Sensitivity under Light and Darkness
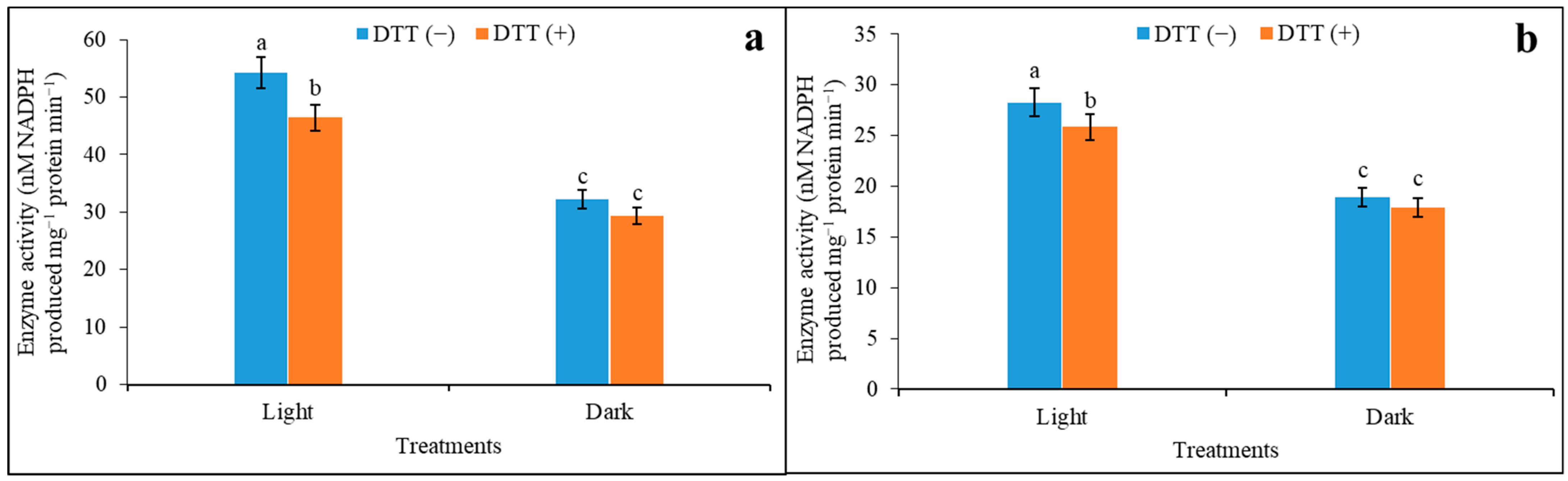
3.3. Effects of Reducing Agent on Activation of Regulatory Properties of NADP-ME under Saturated and Limiting Malate Concentration
3.4. Variability of Polymorphism of NADP-ME under Salinity
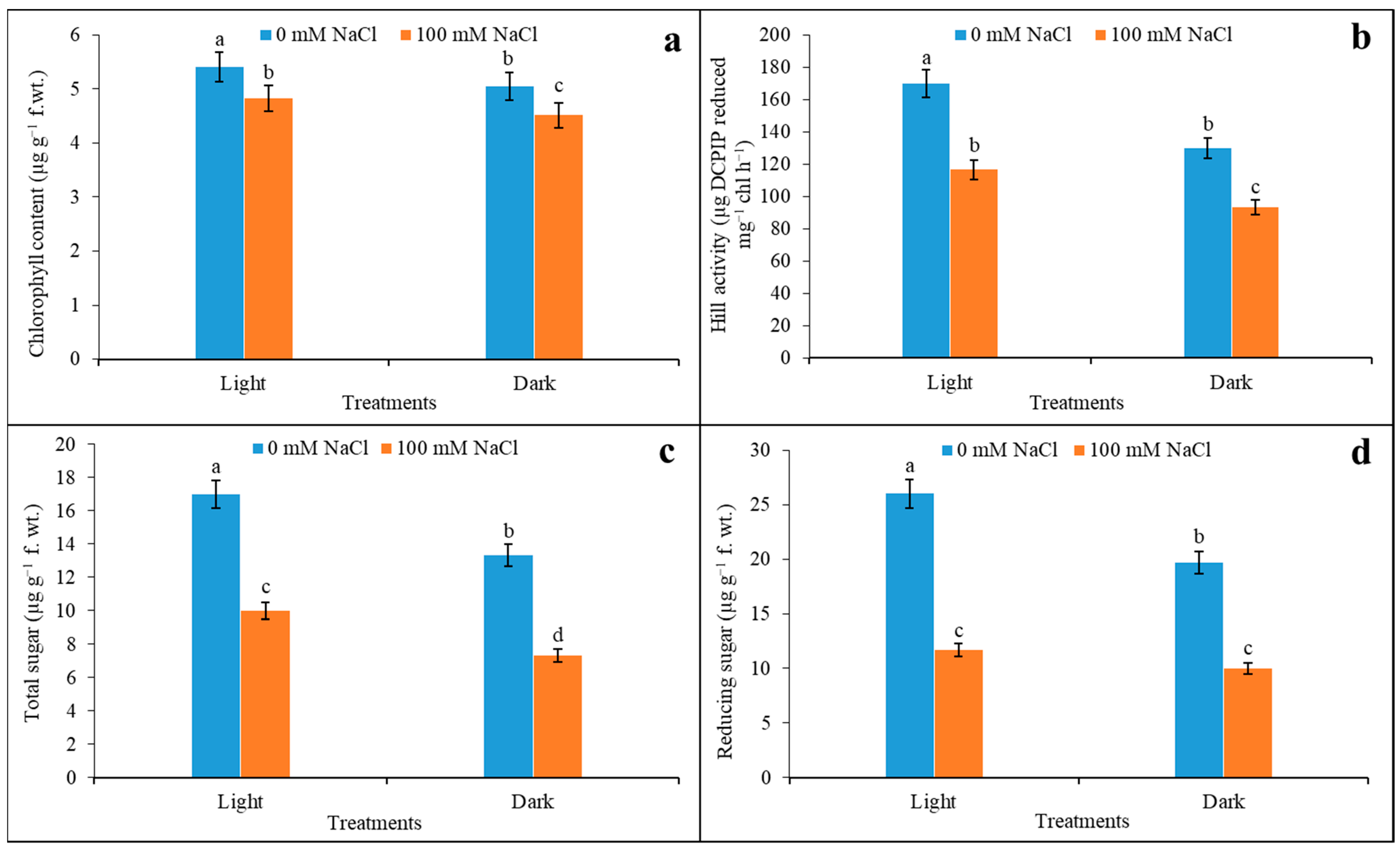
3.5. Chlorophyll Content, Hill Activity, and Sugar Content
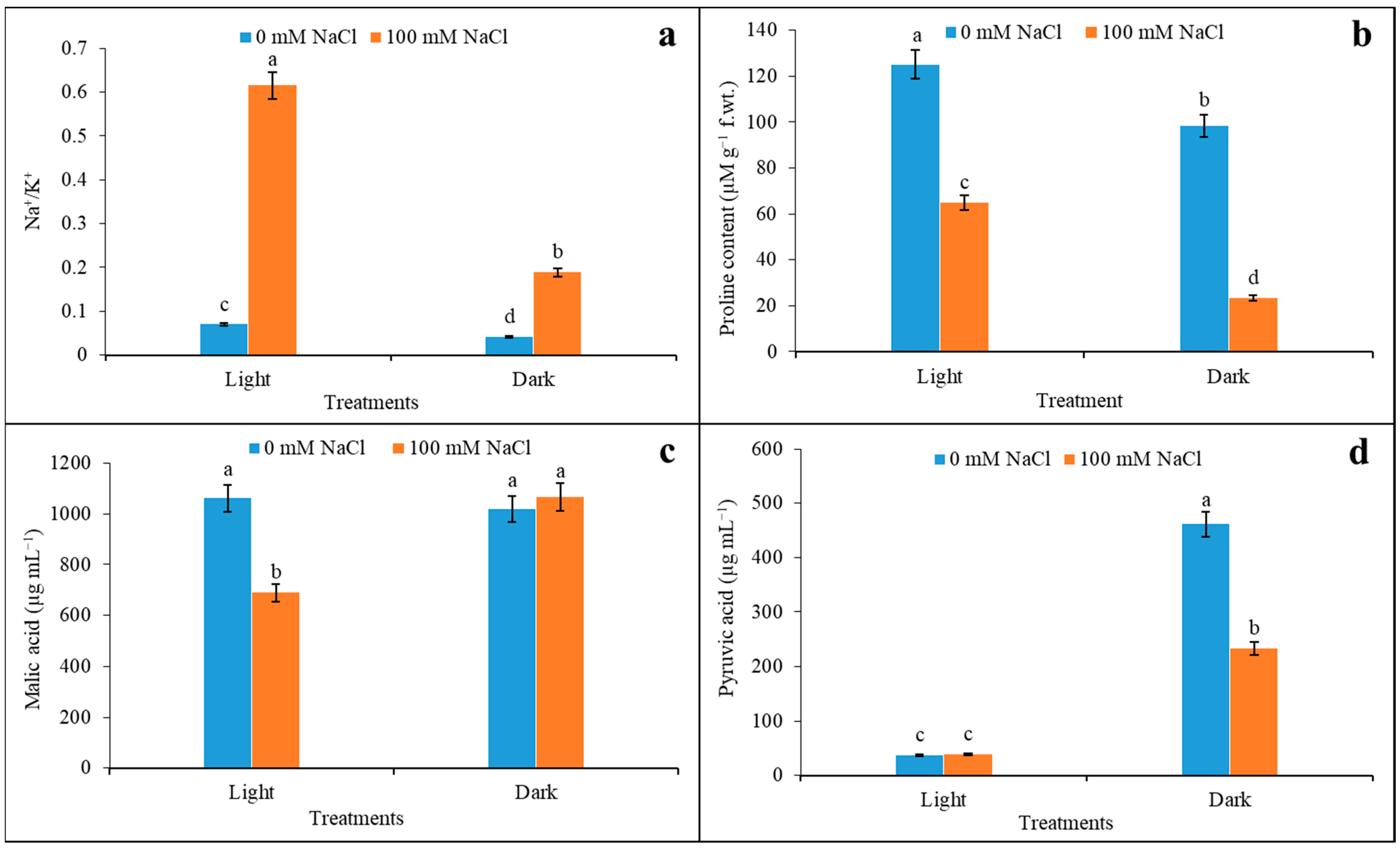
3.6. Na+/K+, Proline Content, Malic Acid, and Pyruvic Acid Content under Salinity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alonso-Cantabrana, H.; Cousins, A.B.; Danila, F.; Ryan, T.; Sharwood, R.E.; von Caemmerer, S.; Furbank, R.T. Diffusion of CO2 across the mesophyll-bundle sheath cell interface in a C4 plant with genetically reduced PEP carboxylase activity. Plant Physiol. 2018, 178, 72–81. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Liu, J.; Wang, Z.; Nai, J.; Lü, M.; Zhou, X.; Cheng, Y. Characterization of the NADP-malic enzymes in the woody plant Populus trichocarpa. Mol. Biol. Rep. 2013, 40, 1385–1396. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, H.; Nagasaki, J.; Yoshikawa, N.; Yamamoto, A.; Takito, S.; Kawasaki, M.; Sugiyama, T.; Miyake, H.; Weber, A.P.; Taniguchi, M. The chloroplastic 2—oxoglutarate/malate transporter has dual function as the malate valve and in carbon/nitrogen metabolism. Plant J. 2011, 65, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Rakhmankulova, Z.F. Photorespiration: Its role in the productive process and evolution of C4 plants. Russ. J. Plant Physiol. 2018, 65, 303–318. [Google Scholar] [CrossRef]
- Jiao, D.; Huang, X.; Li, X.; Chi, W.; Kuang, T.; Zhang, Q.; Ku, M.S.; Cho, D. Photosynthetic characteristics and tolerance to photo-oxidation of transgenic rice expressing C4 photosynthesis enzymes. Photosynth. Res. 2002, 72, 85–93. [Google Scholar] [CrossRef]
- Moreno-Villena, J.J.; Zhou, H.; Gilman, I.S.; Tausta, S.L.; Cheung, C.M.; Edwards, E.J. Spatial resolution of an integrated C4 +CAM photosynthetic metabolism. Sci. Adv. 2022, 8, eabn2349. [Google Scholar] [CrossRef]
- Sarkar, B.; Hasanuzzaman, M.; Adak, M.K. Insights into the Role of Iron Supplementation in Conferring Bicarbonate-Mediated Alkaline Stress Tolerance in Maize. J. Soil Sci. Plant Nutr. 2022, 22, 2719–2734. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, B.; Ding, H.; Zhang, J.; Li, S. The role of NADP-malic enzyme in plants under stress. Plant Sci. 2019, 281, 206–212. [Google Scholar] [CrossRef]
- Giordano, M.; Petropoulos, S.A.; Rouphael, Y. Response and defence mechanisms of vegetable crops against drought, heat and salinity stress. Agriculture 2021, 11, 463. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Sarkar, B.; De, A.K.; Saha, I.; Ghosh, A.; Debnath, S.C.; Adak, M.K. Amelioration with titanium dioxide nanoparticle for regulation of oxidative stress in maize (Zea mays L.). J. Microbiol. Biotechnol. Food Sci. 2019, 9, 320–329. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Murmu, J.; Chinthapalli, B.; Raghavendra, A.S. Light activation of NADP malic enzyme in leaves of maize: Marginal increase in activity, but marked change in regulatory properties of enzyme. J. Plant Physiol. 2003, 160, 51–56. [Google Scholar] [CrossRef]
- Chang, G.G.; Wang, J.K.; Huang, T.M.; Lee, H.J.; Chou, W.Y.; Meng, C.L. Purification and characterization of the cytosolic NADP+-dependent malic enzyme from human breast cancer cell line. Eur. J. Biochem. 1991, 202, 681–688. [Google Scholar] [CrossRef]
- Liang, Y.; Urano, D.; Liao, K.L.; Hedrick, T.L.; Gao, Y.; Jones, A.M. A nondestructive method to estimate the chlorophyll content of Arabidopsis seedlings. Plant Methods 2017, 13, 1–10. [Google Scholar] [CrossRef]
- Sawhney, S.K.; Naik, M.S. Role of light in the synthesis of nitrate reductase and nitrite reductase in rice seedlings. Biochem. J. 1972, 130, 475–485. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Hedge, J.E.; Hofreiter, B.T. Determination of reducing sugars and carbohydrates: Anthrone colorimetric method. In Methods in Carbohydrate Chemistry; Whistler, L.R., Ed.; Academic Press: Cambridge, UK, 1962; pp. 389–390. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Rashed, M.N. Trace elements in some wild plants from the shores of the high dam lake and the adjacent desert, as determined by atomic absorption spectroscopy. J. Arid. Environ. 1995, 29, 185–197. [Google Scholar] [CrossRef]
- Lee, H.S. HPLC method for separation and determination of non-volatile organic acids in orange juice. J. Agric. Food Chem. 1993, 41, 1991–1993. [Google Scholar] [CrossRef]
- Michelet, L.; Zaffagnini, M.; Morisse, S.; Sparla, F.; Pérez-Pérez, M.E.; Francia, F.; Danon, A.; Marchand, C.H.; Fermani, S.; Trost, P.; et al. Redox regulation of the Calvin–Benson cycle: Something old, something new. Front. Plant Sci. 2013, 4, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahra, N.; Al Hinai, M.S.; Hafeez, M.B.; Rehman, A.; Wahid, A.; Siddique, K.H.; Farooq, M. Regulation of photosynthesis under salt stress and associated tolerance mechanisms. Plant Physiol. Biochem. 2022, 178, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yu, H.; Zhou, J.M.; Smith, S.M.; Li, J. Malate circulation: Linking chloroplast metabolism to mitochondrial ROS. Trends Plant Sci. 2020, 25, 446–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweetman, C.; Deluc, L.G.; Cramer, G.R.; Ford, C.M.; Soole, K.L. Regulation of malate metabolism in grape berry and other developing fruits. Phytochemistry 2009, 70, 1329–1344. [Google Scholar] [CrossRef]
- Wilson, S.; Johnson, M.P.; Ruban, A.V. Proton motive force in plant photosynthesis dominated by ΔpH in both low and high light. Plant Physiol. 2021, 187, 263–275. [Google Scholar] [CrossRef]
- Ewe, D.; Tachibana, M.; Kikutani, S.; Gruber, A.; Río Bártulos, C.; Konert, G.; Kaplan, A.; Matsuda, Y.; Kroth, P.G. The intracellular distribution of inorganic carbon fixing enzymes does not support the presence of a C4 pathway in the diatom Phaeodactylum tricornutum. Photosynth. Res. 2018, 137, 263–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, M. The roles of organic acids in C4 photosynthesis. Front. Plant Sci. 2016, 7, 647. [Google Scholar] [CrossRef] [Green Version]
- Igamberdiev, A.U.; Bykova, N.V. Role of organic acids in the integration of cellular redox metabolism and mediation of redox signalling in photosynthetic tissues of higher plants. Free Radic. Biol. Med. 2018, 122, 74–85. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Eprintsev, A.T. Organic acids: The pools of fixed carbon involved in redox regulation and energy balance in higher plants. Front. Plant Sci. 2016, 7, 1042. [Google Scholar] [CrossRef] [Green Version]
- Abrahám, E.; Rigó, G.; Székely, G.; Nagy, R.; Koncz, C.; Szabados, L. Light-dependent induction of proline biosynthesis by abscisic acid and salt stress is inhibited by brassinosteroid in Arabidopsis. Plant Mol. Biol. 2003, 51, 363–372. [Google Scholar] [CrossRef]
- Alvarez, C.E.; Detarsio, E.; Moreno, S.; Andreo, C.S.; Drincovich, M.F. Functional characterization of residues involved in redox modulation of maize photosynthetic NADP-malic enzyme activity. Plant Cell Physiol. 2012, 53, 1144–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, A.; Roychoudhury, A. Plant responses to light stress: Oxidative damages, photoprotection, and role of phytohormones. In Plant Hormones under Challenging Environmental Factors; Ahammed, G.J., Yu, J.Q., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 181–213. [Google Scholar]
- Hýsková, V.D.; Miedzińska, L.; Dobrá, J.; Vankova, R.; Ryšlavá, H. Phosphoenolpyruvate carboxylase, NADP-malic enzyme, and pyruvate, phosphate dikinase are involved in the acclimation of Nicotiana tabacum L. to drought stress. J. Plant Physiol. 2014, 171, 19–25. [Google Scholar] [CrossRef]
- Bose, J.; Munns, R.; Shabala, S.; Gilliham, M.; Pogson, B.; Tyerman, S.D. Chloroplast function and ion regulation in plants growing on saline soils: Lessons from halophytes. J. Exp. Bot. 2017, 68, 3129–3143. [Google Scholar] [CrossRef] [PubMed]
- Romanowska, E.; Buczyńska, A.; Wasilewska, W.; Krupnik, T.; Drożak, A.; Rogowski, P.; Parys, E.; Zienkiewicz, M. Differences in photosynthetic responses of NADP-ME type C4 species to high light. Planta 2017, 245, 641–657. [Google Scholar] [CrossRef] [Green Version]
- Babayev, H.; Mehvaliyeva, U.; Aliyeva, M.; Feyziyev, Y.; Guliyev, N. The study of NAD-malic enzyme in Amaranthus cruentus L. under drought. Plant Physiol. Biochem. 2014, 81, 84–89. [Google Scholar] [CrossRef]
- Doubnerová, V.; Ryšlavá, H. What can enzymes of C4 photosynthesis do for C3 plants under stress? Plant Sci. 2011, 180, 575–583. [Google Scholar] [CrossRef]
- Nunes-Nesi, A.; Fernie, A.R.; Stitt, M. Metabolic and signaling aspects underpinning the regulation of plant carbon nitrogen interactions. Mol. Plant 2010, 3, 973–996. [Google Scholar] [CrossRef]
- Perkins, A.; Nelson, K.J.; Williams, J.R.; Parsonage, D.; Poole, L.B.; Karplus, P.A. The sensitive balance between the fully folded and locally unfolded conformations of a model peroxiredoxin. Biochemistry 2013, 52, 8708–8721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitrac, H.; Mallampalli, V.K.; Dowhan, W. Importance of phosphorylation/dephosphorylation cycles on lipid-dependent modulation of membrane protein topology by posttranslational phosphorylation. J. Biol. Chem. 2019, 294, 18853–18862. [Google Scholar] [CrossRef] [PubMed]
- Farias de Aragao, M.E.; Jolivet, Y.; Guia Silva Lima, M.; Fernandes de Melo, D.; Dizengremel, P. NaCl-induced changes of NAD(P)-malic enzyme activities in Eucalyptus citriodora leaves. Trees 1997, 12, 66–72. [Google Scholar] [CrossRef]
- Omoto, E.; Taniguchi, M.; Miyake, H. Adaptation responses in C4 photosynthesis of maize under salinity. J. Plant Physiol. 2012, 169, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Spampinato, C.P.; Ferreyra, M.L.; Andreo, C.S. Conformational changes of maize and wheat NADP-malic enzyme studied by quenching of protein native fluorescence. Int. J. Biol. Macromol. 2007, 41, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Bulychev, A.A.; Komarova, A.V. Photoregulation of photosystem II activity mediated by cytoplasmic streaming in Chara and its relation to pH bands. Biochim. Biophys. Acta Bioenerg. 2017, 1858, 386–395. [Google Scholar] [CrossRef]
- Zou, X.X.; Xing, S.S.; Su, X.; Zhu, J.; Huang, H.Q.; Bao, S.X. The effects of temperature, salinity and irradiance upon the growth of Sargassum polycystum C. Agardh (Phaeophyceae). J. Appl. Phycol. 2018, 30, 1207–1215. [Google Scholar] [CrossRef]
- Zhou, S.; Thannhauser, T.W. Plant Cell and Organellar Proteomics and Salinity Tolerance in Plants. In Managing Salt Tolerance in Plants: Molecular and Genomic Perspectives; Wani, S.H., Hossain, M.A., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 115–131. [Google Scholar]
- Arnao, M.B.; Hernández-Ruiz, J.; Cano, A.; Reiter, R.J. Melatonin and carbohydrate metabolism in plant cells. Plants 2021, 10, 1917. [Google Scholar] [CrossRef] [PubMed]
- Tanveer, M.; Ahmed, H.A. ROS signalling in modulating salinity stress tolerance in plants. In Salt and Drought Stress Tolerance in Plants; Hasanuzzaman, M., Tanveer, M., Eds.; Springer: Cham, Switzerland, 2020; pp. 299–314. [Google Scholar]
- Khare, T.; Kumar, V.; Kishor, P.B. Na+ and Cl− ions show additive effects under NaCl stress on induction of oxidative stress and the responsive antioxidative defense in rice. Protoplasma 2015, 252, 1149–1165. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Arrivault, S.; Coe, R.A.; Karki, S.; Covshoff, S.; Bagunu, E.; Lunn, J.E.; Stitt, M.; Furbank, R.T.; Hibberd, J.M.; et al. A partial C4 photosynthetic biochemical pathway in rice. Front. Plant Sci. 2020, 11, 564463. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Km (mM/L) | Vmax (nM min−1 mg−1 Protein) | ||
|---|---|---|---|---|
| Light | Dark | Light | Dark | |
| 0 mM NaCl | 0.09 ± 0.006 a | 0.11 ± 0.006 ab | 61.89 ± 1.33 a | 43.89 ± 1.80 b |
| 100 mM NaCl | 0.12 ± 0.01 b | 0.15 ± 0.01 c | 64.46 ± 2.10 a | 47.1 ± 1.70 bc |
| Treatments | Saturated (4.0 mM) Malate Concentration | Limiting (0.01 mM) Malate Concentration | ||||
|---|---|---|---|---|---|---|
| Light | Dark | L/D | Light | Dark | L/D | |
| DTT (−) | 54.32 ± 1.12 a | 32.24 ± 1.36 c | 1.68 | 28.28 ± 0.64 a | 18.92 ± 0.90 c | 1.49 |
| DTT (+) | 46.42 ± 1.52 b | 29.32 ± 0.77 c | 1.58 | 25.83 ± 0.78 b | 17.89 ± 1.01 c | 1.44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarkar, B.; Das, A.; Pal, S.; Kundu, A.; Hasanuzzaman, M.; Fujita, M.; Adak, M.K. Regulation of NADP-Malic Enzyme Activity in Maize (Zea mays L.) under Salinity with Reference to Light and Darkness. Plants 2023, 12, 1836. https://doi.org/10.3390/plants12091836
Sarkar B, Das A, Pal S, Kundu A, Hasanuzzaman M, Fujita M, Adak MK. Regulation of NADP-Malic Enzyme Activity in Maize (Zea mays L.) under Salinity with Reference to Light and Darkness. Plants. 2023; 12(9):1836. https://doi.org/10.3390/plants12091836
Chicago/Turabian StyleSarkar, Bipul, Abir Das, Sayan Pal, Ankita Kundu, Mirza Hasanuzzaman, Masayuki Fujita, and Malay Kumar Adak. 2023. "Regulation of NADP-Malic Enzyme Activity in Maize (Zea mays L.) under Salinity with Reference to Light and Darkness" Plants 12, no. 9: 1836. https://doi.org/10.3390/plants12091836
APA StyleSarkar, B., Das, A., Pal, S., Kundu, A., Hasanuzzaman, M., Fujita, M., & Adak, M. K. (2023). Regulation of NADP-Malic Enzyme Activity in Maize (Zea mays L.) under Salinity with Reference to Light and Darkness. Plants, 12(9), 1836. https://doi.org/10.3390/plants12091836