
Ethylene Inhibition Reduces De Novo Shoot Organogenesis and Subsequent Plant Development from Leaf Explants of Solanum betaceum Cav.



Abstract
:1. Introduction
2. Results
2.1. Effect of Ethylene Modulation on De Novo Shoot Organogenesis
2.2. Effect of Ethylene Modulation on the Expression of Ethylene-Related Genes and Auxin Efflux Carrier
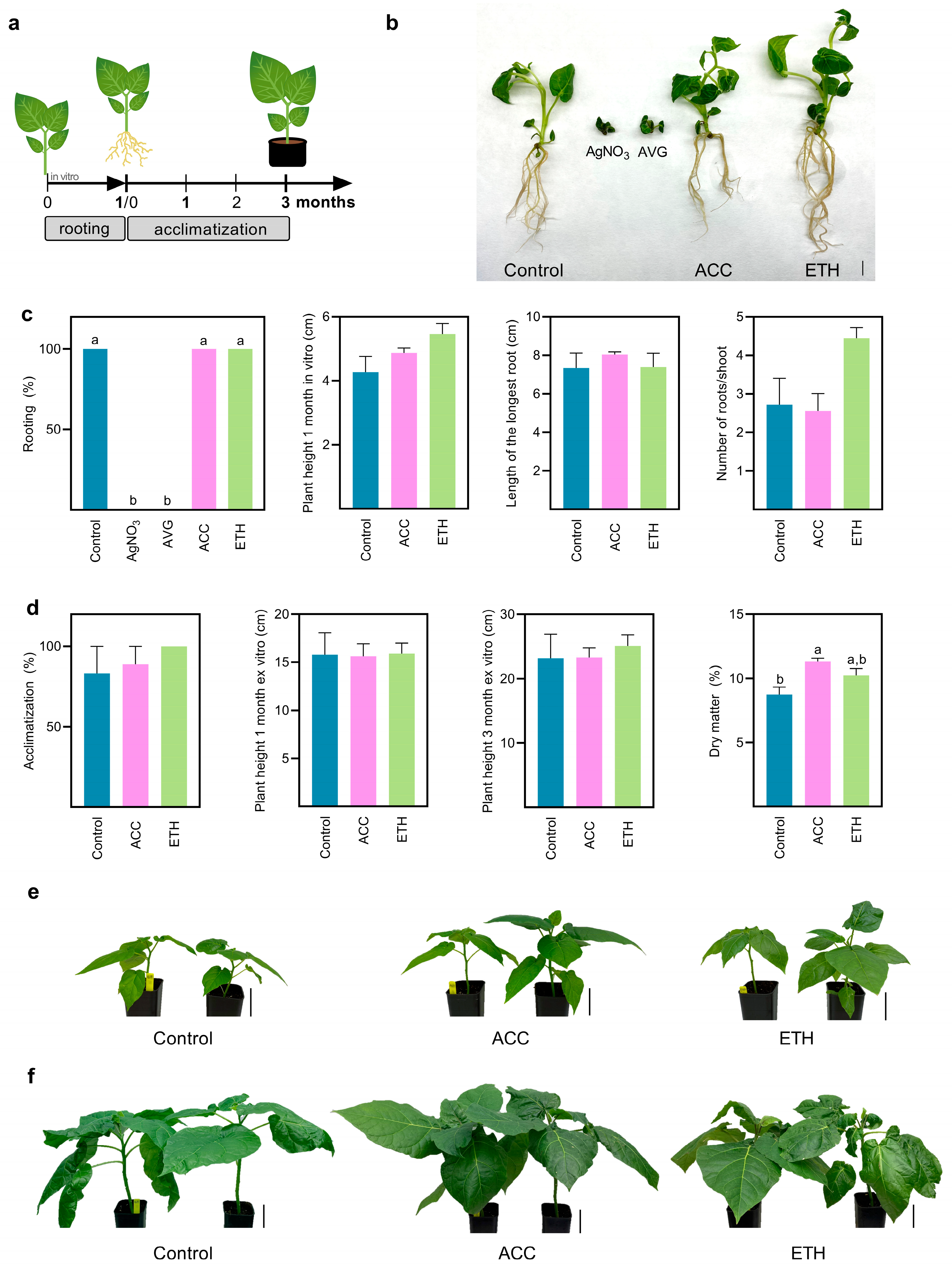
2.3. In Vitro Rotting, Plant Development and Acclimatization
3. Discussion
3.1. Ethylene Positively Modulates Shoot Regeneration from Callus Induction to Shoot Development
3.2. Ethylene Modulation Differentially Regulates Gene Expression Related to Ethylene Biosynthesis and ERF061 Depending on Regeneration Stage
3.3. Ethylene Precursors Upregulate PIN1 Expression in Cytokinin-Induced Callus
3.4. Inhibition of Ethylene Biosynthesis and Perception during Shoot Regeneration Negatively Impacts Subsequent Plant Development
4. Materials and Methods
4.1. Plant Material
4.2. Shoot Regeneration and Culture Conditions
4.3. Total RNA Isolation and Quantitative PCR Analysis
4.4. Rooting and Acclimatization
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dubois, M.; Van den Broeck, L.; Inze, D. The pivotal role of ethylene in plant growth. Trends Plant. Sci. 2018, 23, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Ruzicka, K.; Ljung, K.; Vanneste, S.; Podhorska, R.; Beeckman, T.; Friml, J.; Benkova, E. Ethylene regulates root growth through effects on auxin biosynthesis and transport-dependent auxin distribution. Plant Cell 2007, 19, 2197–2212. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.R.; Negi, S.; Sukumar, P.; Muday, G.K. Ethylene inhibits lateral root development, increases IAA transport and expression of PIN3 and PIN7 auxin efflux carriers. Development 2011, 138, 3485–3495. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Xu, P.; Li, B.; Li, P.; Wen, X.; An, F.; Gong, Y.; Xin, Y.; Zhu, Z.; Wang, Y.; et al. Ethylene promotes root hair growth through coordinated EIN3/EIL1 and RHD6/RSL1 activity in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, 13834–13839. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Tao, R.; Feng, Y.; Xiao, Z.; Zhang, D.; Peng, Y.; Wen, X.; Wang, Y.; Guo, H. EIN3 and RSL4 interfere with an MYB-bHLH-WD40 complex to mediate ethylene-induced ectopic root hair formation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2021, 118, e2110004118. [Google Scholar] [CrossRef]
- Chen, H.; Bullock, D.A., Jr.; Alonso, J.M.; Stepanova, A.N. To fight or to grow: The balancing role of ethylene in plant abiotic stress responses. Plants 2022, 11, 33. [Google Scholar] [CrossRef]
- Jasrotia, S.; Jastoria, R. Role of ethylene in combating biotic stress. In Ethylene in Plant Biology, 1st ed.; Volume, 1, Singh, S., Husain, T., Singh, V.P., Tripathi, D.K., Prasad, S.M., Dubey, N.K., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2022; pp. 388–397. [Google Scholar]
- Fatma, M.; Asgher, M.; Iqbal, N.; Rasheed, F.; Sehar, Z.; Sofo, A.; Khan, N.A. Ethylene signaling under stressful environments: Analyzing collaborative knowledge. Plants 2022, 11, 2211. [Google Scholar] [CrossRef]
- Duclercq, J.; Sangwan-Norreel, B.; Catterou, M.; Sangwan, R.S. De novo shoot organogenesis: From art to science. Trends Plant Sci. 2011, 16, 597–606. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Ogawa, Y.; Iwase, A.; Sugimoto, K. Plant regeneration: Cellular origins and molecular mechanisms. Development 2016, 143, 1442–1451. [Google Scholar] [CrossRef]
- Shin, J.; Bae, S.; Seo, P.J. De novo shoot organogenesis during plant regeneration. J. Exp. Bot. 2020, 71, 63–72. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Favero, D.S.; Sakamoto, Y.; Iwase, A.; Coleman, D.; Rymen, B.; Sugimoto, K. Molecular mechanisms of plant regeneration. Annu. Rev. Plant. Biol. 2019, 70, 3.1–3.30. [Google Scholar] [CrossRef] [PubMed]
- Ikeuchi, M.; Iwase, A.; Rymen, B.; Lambolez, A.; Kojima, M.; Takebayashi, Y.; Heyman, J.; Watanabe, S.; Seo, M.; De Veylder, L.; et al. Wounding triggers callus formation via dynamic hormonal and transcriptional changes. Plant Physiol. 2017, 175, 1158–1174. [Google Scholar] [CrossRef] [PubMed]
- Boller, T.; Kende, H. Regulation of wound ethylene synthesis in plants. Nature 1980, 286, 259–260. [Google Scholar] [CrossRef]
- Chae, H.S.; Faure, F.; Kieber, J.J. The eto1, eto2, and eto3 mutations and cytokinin treatment increase ethylene biosynthesis in Arabidopsis by increasing the stability of ACS protein. Plant Cell 2003, 15, 545–559. [Google Scholar] [CrossRef]
- Hansen, M.; Chae, H.S.; Kieber, J.J. Regulation of ACS protein stability by cytokinin and brassinosteroid. Plant J. 2009, 57, 606–614. [Google Scholar] [CrossRef]
- Tsuchisaka, A.; Theologis, A. Unique and overlapping expression patterns among the Arabidopsis 1-amino-cyclopropane-1-carboxylate synthase gene family members. Plant. Physiol. 2004, 136, 2982–3000. [Google Scholar] [CrossRef]
- Sang, Y.L.; Cheng, Z.J.; Zhang, X.S. Plant stem cells and de novo organogenesis. New Phytol. 2018, 218, 1334–1339. [Google Scholar] [CrossRef]
- Xu, L.; Huang, H. Genetic and epigenetic controls of plant regeneration. Curr. Top. Dev. Biol. 2014, 108, 1–33. [Google Scholar] [CrossRef]
- Neves, M.; Correia, S.; Cavaleiro, C.; Canhoto, J. Modulation of organogenesis and somatic embryogenesis by ethylene: An overview. Plants 2021, 10, 1208. [Google Scholar] [CrossRef]
- Amor, M.B.; Guis, M.; Latché, A.; Bouzayen, M.; Pech, J.C.; Roustan, J.P. Expression of an antisense 1-aminocyclopropane-1-carboxylate oxidase gene stimulates shoot regeneration in Cucumis melo. Plant Cell Rep. 1998, 17, 586–589. [Google Scholar] [CrossRef]
- Roustan, J.-P.; Latche, A.; Fallot, J. Enhancement of shoot regeneration from cotyledons of Cucumis melo by AgNO3, an inhibitor of ethylene action. J. Plant Physiol. 1992, 140, 485–488. [Google Scholar] [CrossRef]
- Paa, E.-C.; Chi, G.-L. De novo shoot morphogenesis and plant growth of mustard (Brassica juncea) in vitro in relation to ethylene. Physiol. Plant. 1993, 88, 467–474. [Google Scholar] [CrossRef]
- Paladi, R.K.; Rai, A.N.; Penna, S. Silver nitrate modulates organogenesis in Brassica juncea (L.) through differential antioxidant defense and hormonal gene expression. Sci. Hortic. 2017, 226, 261–267. [Google Scholar] [CrossRef]
- Trujillo-Moya, C.; Gisbert, C. The influence of ethylene and ethylene modulators on shoot organogenesis in tomato. Plant Cell Tissue Organ. Cult. 2012, 111, 41–48. [Google Scholar] [CrossRef]
- Chatfield, S.P.; Raizada, M.N. Ethylene and shoot regeneration: hookless1 modulates de novo shoot organogenesis in Arabidopsis thaliana. Plant Cell Rep. 2008, 27, 655–666. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Zheng, Y.; Perry, S.E. AGAMOUS-Like15 promotes somatic embryogenesis in Arabidopsis and soybean in part by the control of ethylene biosynthesis and response. Plant Physiol. 2013, 161, 2113–2127. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Nolan, T.M.; Jiang, H.; Yin, Y. AP2/ERF transcription factor regulatory networks in hormone and abiotic stress responses in Arabidopsis. Front Plant Sci. 2019, 10, 228. [Google Scholar] [CrossRef]
- Muller, M.; Munne-Bosch, S. Ethylene response factors: A key regulatory hub in hormone and stress signaling. Plant Physiol. 2015, 169, 32–41. [Google Scholar] [CrossRef]
- Mantiri, F.R.; Kurdyukov, S.; Lohar, D.P.; Sharopova, N.; Saeed, N.A.; Wang, X.D.; Vandenbosch, K.A.; Rose, R.J. The transcription factor MtSERF1 of the ERF subfamily identified by transcriptional profiling is required for somatic embryogenesis induced by auxin plus cytokinin in Medicago truncatula. Plant Physiol. 2008, 146, 1622–1636. [Google Scholar] [CrossRef]
- Murillo-Gómez, P.A.; Hoyos, R.S.; Chavarriaga, P. Organogenesis in-vitro using three tissues types of tree tomato [Solanum betaceum (Cav.)]. Agron. Colomb. 2017, 35, 5–11. [Google Scholar] [CrossRef]
- Obando, M.; Jordan, M. Regenerative responses of Cyphomandra betacea (Cav.) sendt. (tamarillo) cultivated in vitro. Acta Hortic. 2001, 560, 429–432. [Google Scholar] [CrossRef]
- Puga, A.P. Desenvolvimento de Biorreatores para a Propagação de Solanum Betaceum Cav. Master’s Thesis, University of Coimbra, Coimbra, Portugal, 2019. [Google Scholar]
- Santos, A.L. Estudos de Organogénese em Cyphomandra betacea (Cav.) Sendt. Master’s Thesis, University of Porto, Porto, Portugal, 2012. [Google Scholar]
- Canhoto, J.; Lopes, M.; Cruz, G. Protocol of somatic embryogenesis: Tamarillo (Cyphomandra betacea (Cav.) Sendt). In Protocols for Somatic Embryogenesis in Woody Plants; Jain, S., Gupta, P., Eds.; Springer: Cham, Switzerland, 2005; Volume 77, pp. 379–389. [Google Scholar]
- Correia, S.; Canhoto, J.M. Somatic embryogenesis of tamarillo (Solanum betaceum Cav.). In Step Wise Protocols for Somatic Embryogenesis of Important Woody Plants; Jain, S., Gupta, P., Eds.; Springer: Cham, Switzerland, 2018; Volume 85, pp. 171–179. [Google Scholar]
- Correia, S.; Lopes, M.L.; Canhoto, J.M. Somatic embryogenesis induction system for cloning an adult Cyphomandra betacea (Cav.) Sendt. (tamarillo). Trees 2011, 25, 1009–1020. [Google Scholar] [CrossRef]
- Corredoira, E.; Merkle, S.A.; Martínez, M.T.; Toribio, M.; Canhoto, J.M.; Correia, S.I.; Ballester, A.; Vieitez, A.M. Non-zygotic embryogenesis in hardwood species. Crit Rev. Plant Sci. 2019, 38, 29–97. [Google Scholar] [CrossRef]
- Vincent, C.; Munné-Bosch, S. Ethylene in the regulation of major biotechnological processes. In The Plant Hormone Ethylene—Stress Acclimation and Agricultural Applications, 1st ed.; Khan, N.A., Ferrante, A., Munné-Bosch, S., Eds.; Academic Press: Cambridge, MA, USA, 2023; Volume 1, pp. 89–105. [Google Scholar]
- Dimasi-Theriou, K.; Economou, A.S.; Sfakiotakis, E.M. Promotion of petunia (Petunia hybrida L.) regeneration in vitro by ethylene. Plant Cell Tissue Organ. Cult. 1993, 32, 219–225. [Google Scholar] [CrossRef]
- Long, Y.; Yang, Y.; Pan, G.; Shen, Y. New isights into tissue culture plant-regeneration mechanisms. Front Plant. Sci. 2022, 13, 926752. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Seo, P.J. Varying auxin levels induce distinct pluripotent states in callus cells. Front Plant Sci. 2018, 9, 1653. [Google Scholar] [CrossRef]
- Zhai, N.; Xu, L. Pluripotency acquisition in the middle cell layer of callus is required for organ regeneration. Nat. Plants 2021, 7, 1453–1460. [Google Scholar] [CrossRef]
- O’Donnell, P.J.; Calvert, C.; Atzorn, R.; Wasternack, C.; Leyser, H.M.O.; Bowles, D.J. Ethylene as a signal mediating the wound response of tomato plants. Science 1996, 274, 1914–1917. [Google Scholar] [CrossRef]
- Suttle, J.C. Cytokinin-induced ethylene biosynthesis in nonsenescing cotton leaves. Plant Physiol. 1986, 82, 930–935. [Google Scholar] [CrossRef]
- Hyodo, H.; Tanaka, K.; Suzuki, T. Wound-induced ethylene synthesis and its involvement in enzyme induction in mesocarp tissue of Cucurbita maxima. Postharvest Biol. Technol. 1991, 1, 127–136. [Google Scholar] [CrossRef]
- Ke, D.; Saltveit, M.E.J. Wound-induced ethylene production, phenolic metabolism and susceptibility to russet spotting in iceberg lettuce. Physiol. Plant. 1989, 76, 412–418. [Google Scholar] [CrossRef]
- Watanabe, T.; Seo, S.; Sakai, S. Wound-induced expression of a gene for 1-aminocyclopropane-1-carboxylate synthase and ethylene production are regulated by both reactive oxygen species and jasmonic acid in Cucurbita maxima. Plant Physiol. Biochem. 2001, 39, 121–127. [Google Scholar] [CrossRef]
- Li, W.; Nishiyama, R.; Watanabe, Y.; Van Ha, C.; Kojima, M.; An, P.; Tian, L.; Tian, C.; Sakakibara, H.; Tran, L.-S.P. Effects of overproduced ethylene on the contents of other phytohormones and expression of their key biosynthetic genes. Plant Physiol. Biochem. 2018, 128, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Salman-Minkov, A.; Levi, A.; Wolf, S.; Trebitsh, T. ACC synthase genes are polymorphic in watermelon (Citrullus spp.) and differentially expressed in flowers and in response to auxin and gibberellin. Plant Cell Physiol. 2008, 49, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Swarup, R.; Perry, P.; Hagenbeek, D.; Van Der Straeten, D.; Beemster, G.T.; Sandberg, G.; Bhalerao, R.; Ljung, K.; Bennett, M.J. Ethylene upregulates auxin biosynthesis in Arabidopsis seedlings to enhance inhibition of root cell elongation. Plant Cell 2007, 19, 2186–2196. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Gupta, S.; Lindquist, I.E.; Cameron, C.T.; Mudge, J.; Rashotte, A.M. Transcriptome analysis of cytokinin response in tomato leaves. PLoS ONE 2013, 8, e55090. [Google Scholar] [CrossRef]
- Shin, S.Y.; Choi, Y.; Kim, S.-G.; Park, S.-J.; Park, J.-S.; Moon, K.-B.; Kim, H.-S.; Jeon, J.H.; Cho, H.S.; Lee, H.-J. Submergence promotes auxin-induced callus formation through ethylene-mediated post-transcriptional control of auxin receptors. Mol. Plant 2022, 15, 1947–1961. [Google Scholar] [CrossRef]
- Lardon, R.; Wijnker, E.; Keurentjes, J.; Geelen, D. The genetic framework of shoot regeneration in Arabidopsis comprises master regulators and conditional fine-tuning factors. Commun. Biol. 2020, 3, 549. [Google Scholar] [CrossRef]
- Iwase, A.; Kondo, Y.; Laohavisit, A.; Takebayashi, A.; Ikeuchi, M.; Matsuoka, K.; Asahina, M.; Mitsuda, N.; Shirasu, K.; Fukuda, H.; et al. WIND transcription factors orchestrate wound-induced callus formation, vascular reconnection and defense response in Arabidopsis. New Phytol. 2021, 232, 734–752. [Google Scholar] [CrossRef]
- Iwase, A.; Mita, K.; Nonaka, S.; Ikeuchi, M.; Koizuka, C.; Ohnuma, M.; Ezura, H.; Imamura, J.; Sugimoto, K. WIND1-based acquisition of regeneration competency in Arabidopsis and rapeseed. J. Plant Res. 2015, 128, 389–397. [Google Scholar] [CrossRef]
- Amrhein, N.; Wenker, D. Novel inhibitors of ethylene production in higher plants. Plant Cell Physiol. 1979, 20, 1635–1642. [Google Scholar] [CrossRef]
- Yu, Y.B.; Yang, S.F. Auxin-induced ethylene production and its inhibition by aminoethoxyvinyiglycine and cobalt ion. Plant Physiol. 1979, 64, 1074–1077. [Google Scholar] [CrossRef] [PubMed]
- Binder, B.M.; Rodriguez, F.I.; Bleecker, A.B.; Patterson, S.E. The effects of Group 11 transition metals, including gold, on ethylene binding to the ETR1 receptor and growth of Arabidopsis thaliana. FEBS Lett. 2007, 581, 5105–5109. [Google Scholar] [CrossRef]
- Wang, C.; Li, W.; Chen, F.; Cheng, Y.; Huang, X.; Zou, B.; Wang, Y.; Xu, W.; Qu, S. Genome-wide identification and characterization of members of the ACS gene family in Cucurbita maxima and their transcriptional responses to the specific treatments. Int. J. Mol. Sci. 2022, 23, 8476. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Feng, Y.; Cheng, Y.; Guan, J. Effects of preharvest aminoethoxyvinylglycine (AVG) treatment on fruit ripening, core browning and related gene expression in ‘Huangguan’ pear (Pyrus bretschneideri Rehd.). Horticulturae 2023, 9, 179. [Google Scholar] [CrossRef]
- Yuan, H.; Yue, P.; Bu, H.; Han, D.; Wang, A. Genome-wide analysis of ACO and ACS genes in pear (Pyrus ussuriensis). Vitr. Cell. Dev. Biol. Plant 2019, 56, 193–199. [Google Scholar] [CrossRef]
- Wang, Z.; Yadav, V.; Yan, X.; Cheng, D.; Wei, C.; Zhang, X. Systematic genome-wide analysis of the ethylene-responsive ACS gene family: Contributions to sex form differentiation and development in melon and watermelon. Gene 2021, 805, 145910. [Google Scholar] [CrossRef]
- Jones, M.L. Ethylene biosynthetic genes are differentially regulated by ethylene and ACC in carnation styles. Plant Growth Regul. 2002, 40, 129–138. [Google Scholar] [CrossRef]
- Coppola, M.; Diretto, G.; Digilio, M.C.; Woo, S.L.; Giuliano, G.; Molisso, D.; Pennacchio, F.; Lorito, M.; Rao, R. Transcriptome and metabolome reprogramming in tomato plants by Trichoderma harzianum strain T22 primes and enhances defense responses against aphids. Front Physiol. 2019, 10, 745. [Google Scholar] [CrossRef]
- Chahel, A.A.; Yousaf, Z.; Zeng, S.; Li, Y.; Ying, W. Growth and physiological alterations related to root-specific gene function of LrERF061-OE in Glycyrrhiza uralensis Fisch. hairy root clones under cadmium stress. Plant Cell Tissue Organ Cult. 2019, 140, 115–127. [Google Scholar] [CrossRef]
- Choi, H.R.; Baek, M.W.; Jeong, C.S.; Tilahun, S. Comparative transcriptome analysis of softening and ripening-related genes in kiwifruit cultivars treated with ethylene. Curr. Issues Mol. Biol. 2022, 44, 2593–2613. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Chen, L.; Yang, Y.; Guo, X.; Chen, G.; Xiong, X.; Dong, D.; Li, G. Transcriptome analysis reveals that exogenous ethylene activates immune and defense responses in a high late blight resistant potato genotype. Sci. Rep. 2020, 10, 21294. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Chen, Y.; Zhang, J.; Wu, F.; Wang, X.; Pan, T.; Wei, Q.; Hao, Y.; Chen, X.; Jiang, C.; et al. Identification, classification, and characterization of AP2/ERF superfamily genes in Masson pine (Pinus massoniana Lamb.). Sci. Rep. 2021, 11, 5441. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Wang, Z.; Hu, G.; Yao, X. Genome-wide identification and characterization of AP2/ERF gene superfamily during flower development in Actinidia eriantha. BMC Genom. 2022, 23, 650. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.P.; Heisler, M.G.; Reddy, G.V.; Ohno, C.; Das, P.; Meyerowitz, E.M. Pattern formation during de novo assembly of the Arabidopsis shoot meristem. Development 2007, 134, 3539–3548. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Arigita, L.; Majada, J.; Sánchez Tamés, R. Ethylene involvement in in vitro organogenesis and plant growth of Populus tremula L. Plant Growth Regul. 1997, 22, 1–6. [Google Scholar] [CrossRef]
- Žiauka, J.; Kuusienė, S.; Šilininkas, M. Fast growing aspens in the development of a plant micropropagation system based on plant-produced ethylene action. Biomass Bioenergy 2013, 53, 20–28. [Google Scholar] [CrossRef]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Trivellini, A.; Francini, A.; Khan, M.I.R. Ethylene role in plant growth, development and senescence: Interaction with other phytohormones. Front. Plant Sci. 2017, 8, 475. [Google Scholar] [CrossRef]
- Lyu, J.; Wu, Y.; Jin, X.; Tang, Z.; Liao, W.; Dawuda, M.M.; Hu, L.; Xie, J.; Yu, J.; Calderon-Urrea, A. Proteomic analysis reveals key proteins involved in ethylene-induced adventitious root development in cucumber (Cucumis sativus L.). PeerJ 2021, 9, e10887. [Google Scholar] [CrossRef]
- Negi, S.; Sukumar, P.; Liu, X.; Cohen, J.D.; Muday, G.K. Genetic dissection of the role of ethylene in regulating auxin-dependent lateral and adventitious root formation in tomato. Plant J. 2010, 61, 3–15. [Google Scholar] [CrossRef]
- Qi, X.; Li, Q.; Ma, X.; Qian, C.; Wang, H.; Ren, N.; Shen, C.; Huang, S.; Xu, X.; Xu, Q.; et al. Waterlogging-induced adventitious root formation in cucumber is regulated by ethylene and auxin through reactive oxygen species signalling. Plant Cell Environ. 2019, 42, 1458–1470. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Cordeiro, D.; Rito, M.; Borges, F.; Canhoto, J.; Correia, S. Selection and validation of reference genes for qPCR analysis of miRNAs and their targets during somatic embryogenesis in tamarillo (Solanum betaceum Cav.). Plant Cell Tissue Organ. Cult. 2020, 143, 109–120. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, E45. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Regeneration Percentage (%) | No of Buds and Shoots per Explant | Observations |
|---|---|---|---|
| Control | 73.61 ± 6.05 a | 12.21 ± 1.19 b | Presence of well-developed and elongated shoots with fully opened leaves. |
| AgNO3 | 65.28 ± 6.05 a | 6.36 ± 1.18 c | Shoots not completely developed nor elongated. Some leaves were fully open but presented abnormal shape. |
| AVG | 48.61 ± 1.39 a | 6.40 ± 1.16 c | Shoots neither developed nor elongated. Some leaves were fully opened. |
| ACC | 66.67 ± 6.37 a | 19.09 ± 1.03 a | Presence of well-developed and elongated shoots with fully opened leaves. |
| ETH | 58.33 ± 6.37 a | 18.00 ± 1.29 a | Presence of well-developed and elongated shoots with fully opened leaves. |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| ACO1 | GCTAACTCTTGGAGCTGGCA | GCCACTACTCTGTGTGCAGT |
| ACS | TCCACAGTGAATCCCATTTTGAT | GGCTTAGCTTTGTTCTTTGTTGT |
| ACT | CCATGTTCCCGGGTATTGCT | GTGCTGAGGGAAGCCAAGAT |
| ERF061 | TCTTCGCGATCCAAGCAAGT | ACCACCACCAACCAAAGAAGA |
| FeSOD | TCACCATCGACGTTTGGGAG | GACTGCTTCCCATGACACCA |
| PIN1 | ACCAAGGATCATAGCATGTGGA | CTTGTGGTAGAGCTGCCTGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neves, M.; Correia, S.; Canhoto, J. Ethylene Inhibition Reduces De Novo Shoot Organogenesis and Subsequent Plant Development from Leaf Explants of Solanum betaceum Cav. Plants 2023, 12, 1854. https://doi.org/10.3390/plants12091854
Neves M, Correia S, Canhoto J. Ethylene Inhibition Reduces De Novo Shoot Organogenesis and Subsequent Plant Development from Leaf Explants of Solanum betaceum Cav. Plants. 2023; 12(9):1854. https://doi.org/10.3390/plants12091854
Chicago/Turabian StyleNeves, Mariana, Sandra Correia, and Jorge Canhoto. 2023. "Ethylene Inhibition Reduces De Novo Shoot Organogenesis and Subsequent Plant Development from Leaf Explants of Solanum betaceum Cav." Plants 12, no. 9: 1854. https://doi.org/10.3390/plants12091854
APA StyleNeves, M., Correia, S., & Canhoto, J. (2023). Ethylene Inhibition Reduces De Novo Shoot Organogenesis and Subsequent Plant Development from Leaf Explants of Solanum betaceum Cav. Plants, 12(9), 1854. https://doi.org/10.3390/plants12091854