

Nitrogen Addition and Heterotroph Exclusion Affected Plant Species Diversity–Biomass Relationship by Affecting Plant Functional Traits

 and
and
Abstract
:1. Introduction
2. Results
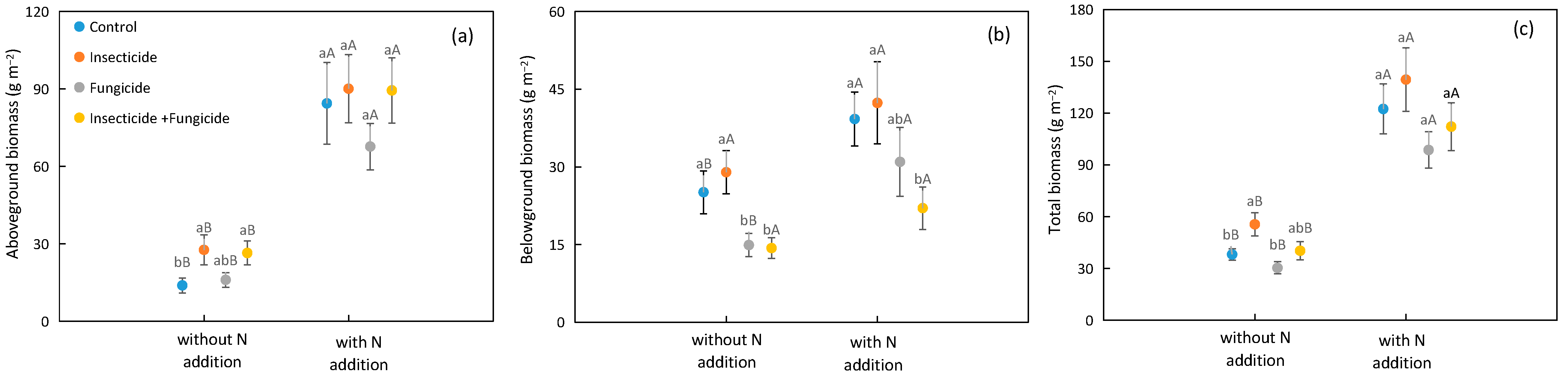
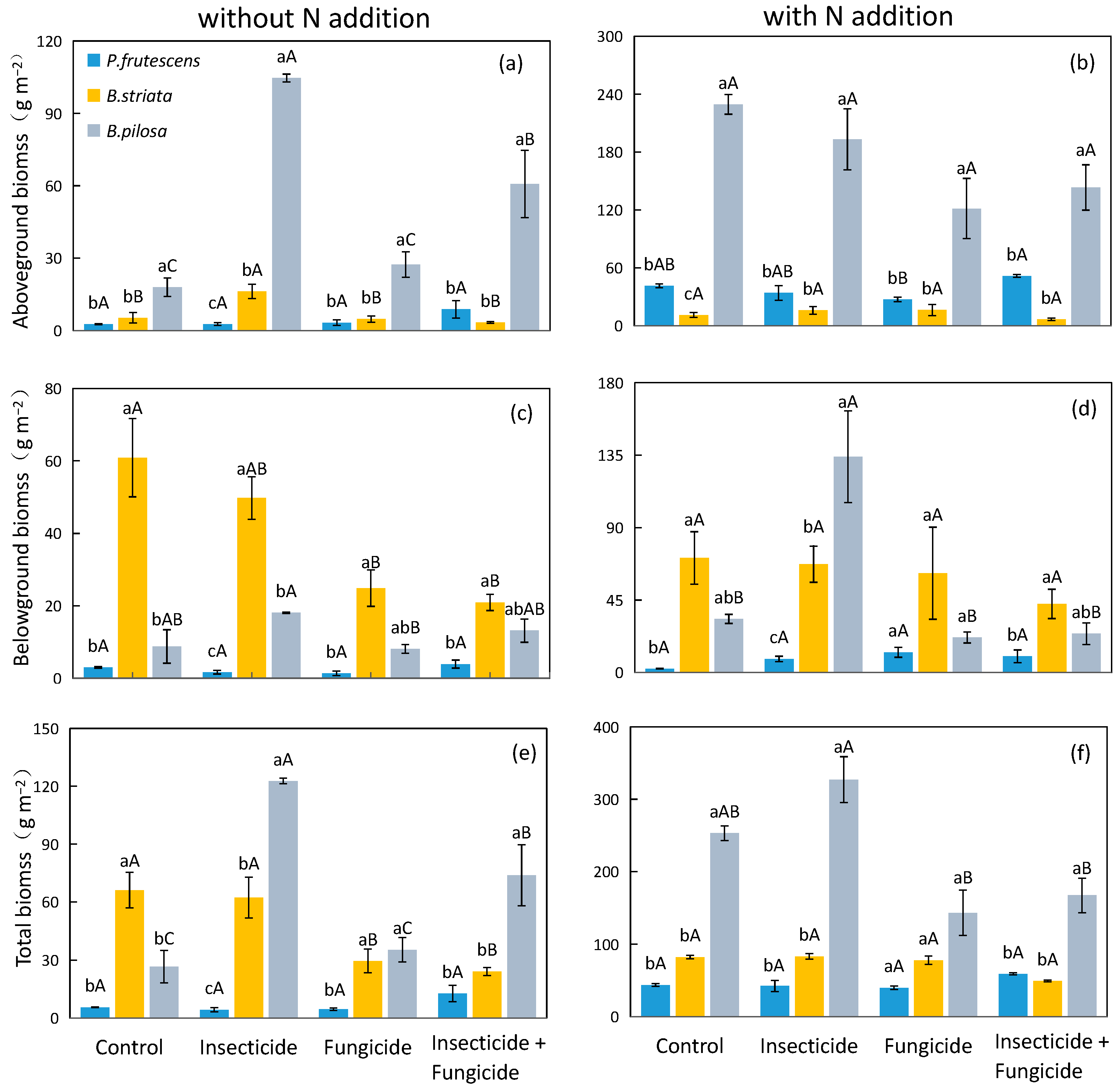
2.1. Plant Biomass Responds to N Addition and Heterotroph Removal
2.2. The Relationship between Plant Species Diversity and Plant Biomass
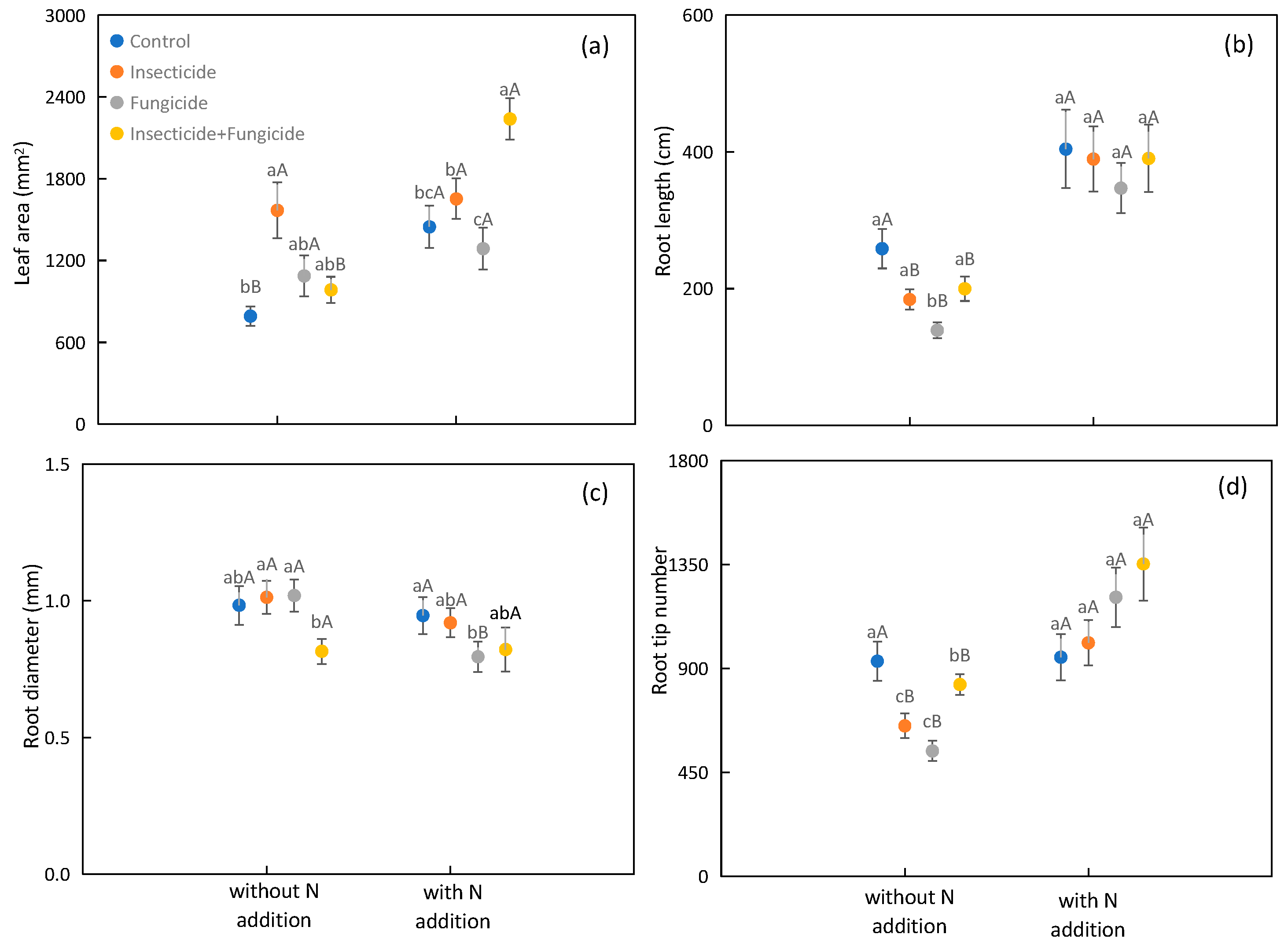
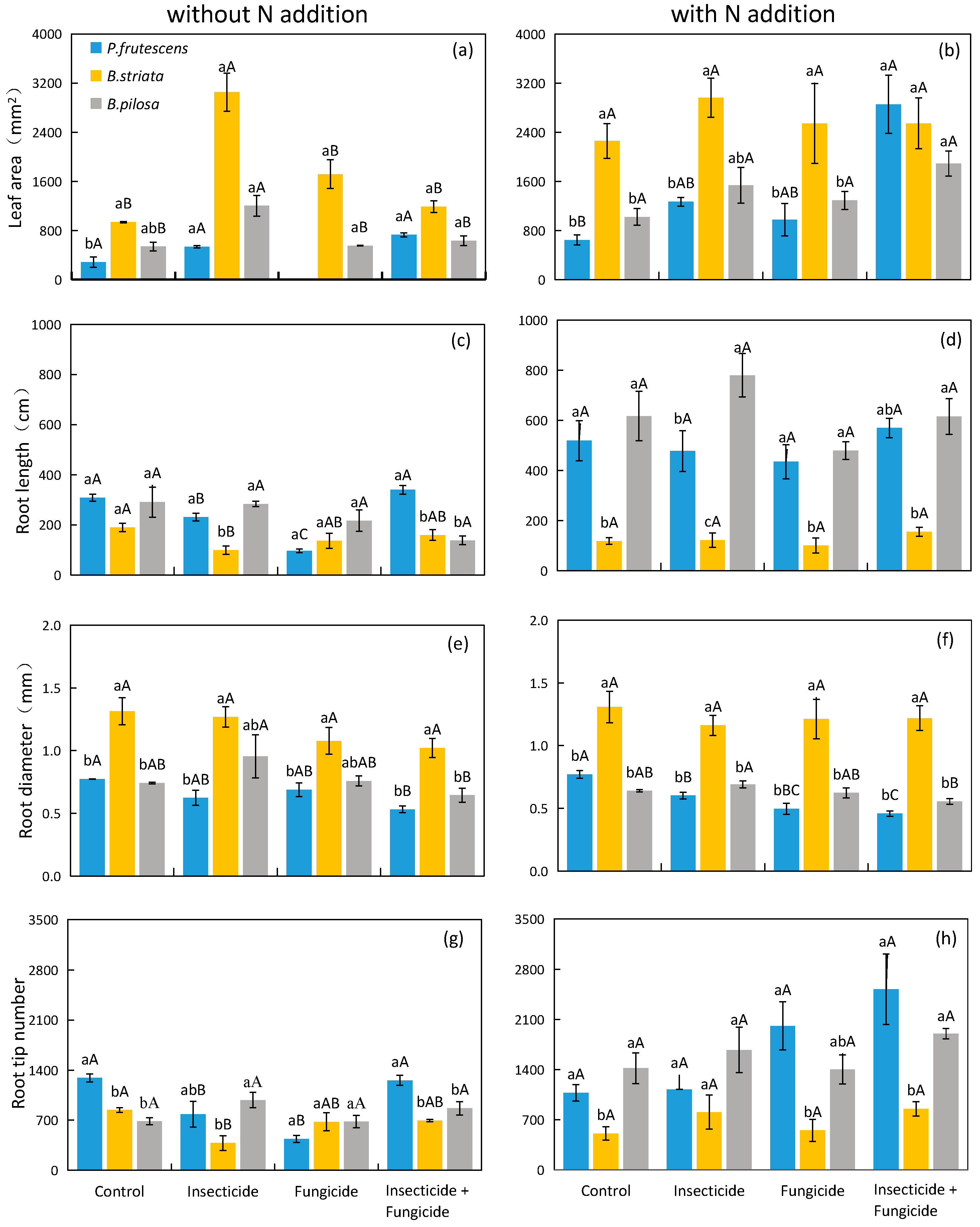
2.3. Functional Traits of Plant Leaves and Roots Respond to N Addition and Heterotroph Removal
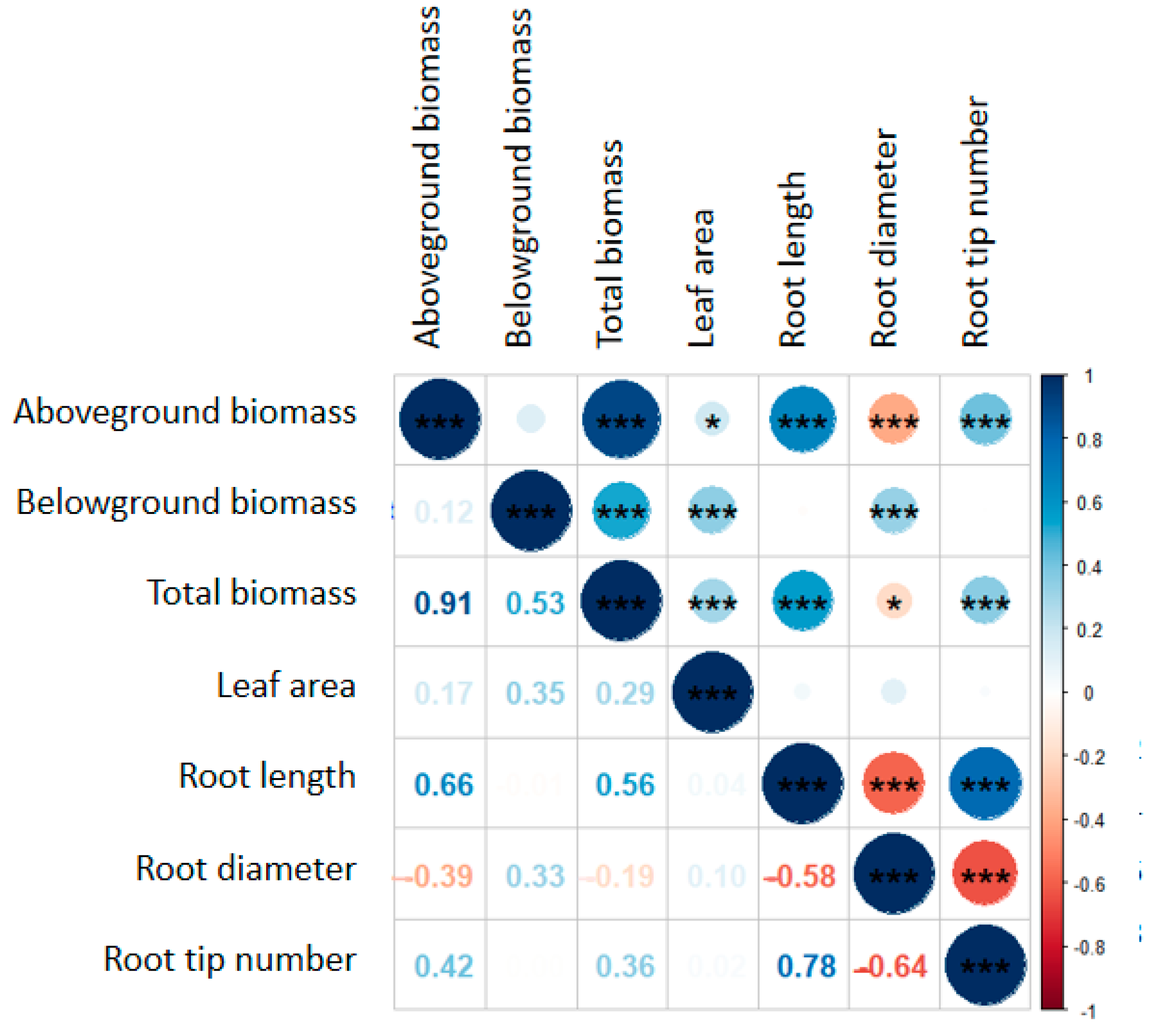
2.4. The Relationship between Plant Species Diversity and Functional Traits of Plant Leaves and Roots
3. Discussion
3.1. The Effect of Heterotroph Removal on Plant Biomass
3.2. The Effect of Heterotroph Removal on Plant Species Richness–Biomass Relationship
3.3. The Effect of Heterotroph Removal on the Effect of Plant Species Identity on Plant Biomass
4. Materials and Methods
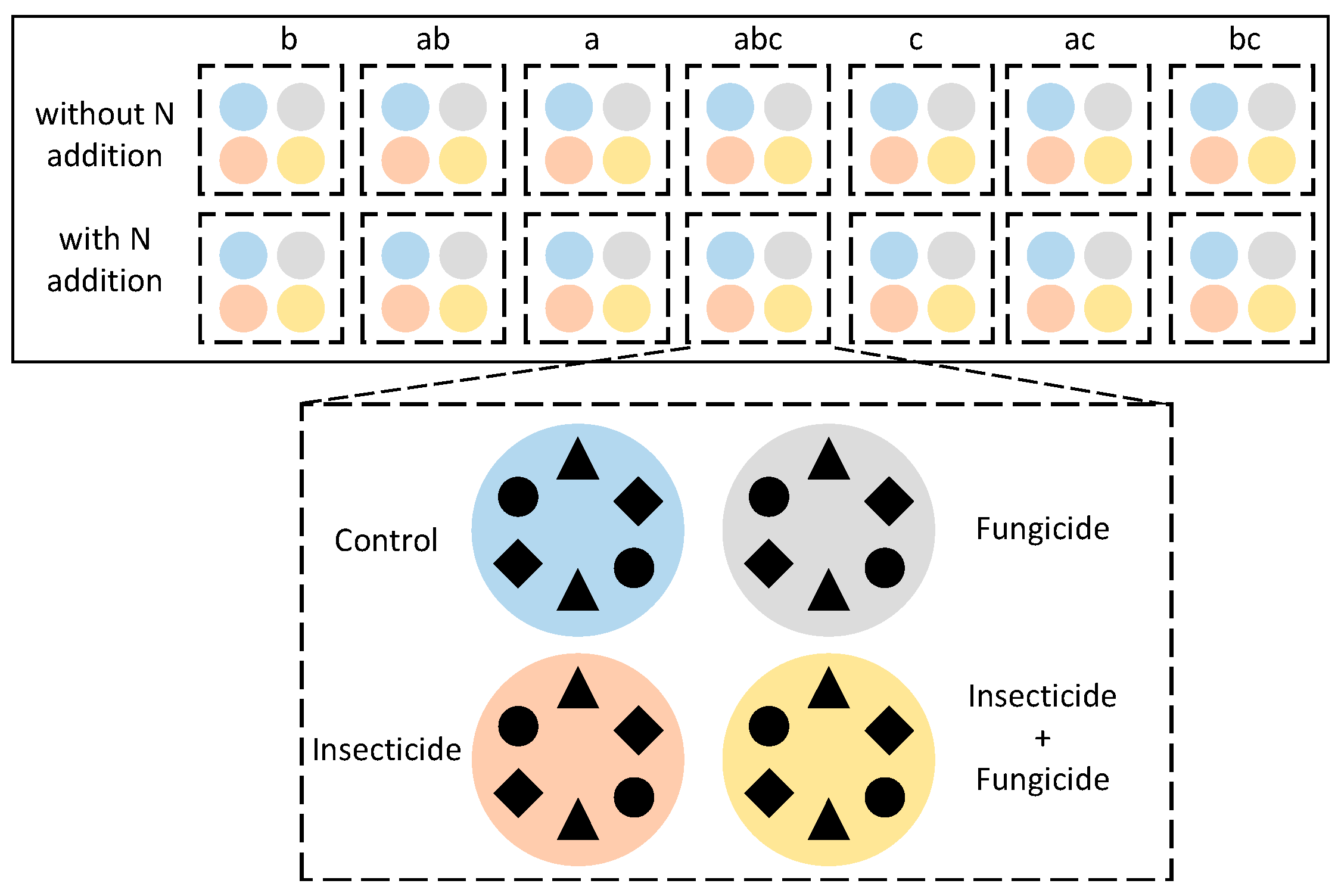
4.1. Experimental Design
4.2. Sample Collection and Calculation
- Plant functional traits of leaves and roots:
- 2.
- Relative yield:
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tilman, D.; Isbell, F.; Cowles, M.J. Biodiversity and ecosystem functioning. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 471–493. [Google Scholar] [CrossRef]
- Tilman, D.; Knops, J.; Wedin, D.; Reich, P.; Ritchie, M.; Siemann, E. The influence of functional diversity and compositions on ecosystem processes. Science 1997, 277, 1300–1302. [Google Scholar] [CrossRef]
- Tilman, D.; Reich, P.B.; Knops, J.; Wedin, D.; Mielke, T.; Lehman, C. Diversity and productivity in a long-term grassland experiment. Science 2001, 294, 843–845. [Google Scholar] [CrossRef]
- van der Plas, F. Biodiversity and ecosystem functioning in naturally assembled communities. Biol. Rev. 2019, 94, 1220–1245. [Google Scholar] [CrossRef]
- He, J.S.; Bazzaz, F.A.; Schmid, B. Interactive effects of diversity nutrients and elevated CO2 on experimental plant communities. Oikos 2002, 97, 337–348. [Google Scholar] [CrossRef]
- Isbell, F.; Reich, P.B.; Tilman, D.; Hobbie, S.E.; Polasky, S.; Binder, S. Nutrient enrichment biodiversity loss and consequent declines in ecosystem productivity. Proc. Natl. Acad. Sci. USA 2013, 110, 11911–11916. [Google Scholar] [CrossRef]
- Roscher, C.; Schmid, B.; Kolle, O.; Schulze, E.D. Complementarity among four highly productive grassland species depends on resource availability. Oecologia 2016, 181, 571–582. [Google Scholar] [CrossRef]
- Seabloom, E.W.; Kinkel, L.; Borer, E.T.; Hautier, Y.; Montgomery, R.A.; Tilman, D. Food webs obscure the strength of plant diversity effects on primary productivity. Ecol. Lett. 2017, 20, 505–512. [Google Scholar] [CrossRef]
- Zaret, M.M.; Kuhs, M.A.; Anderson, J.C.; Seabloom, E.W.; Borer, E.T.; Kinkel, L.L. Seasonal shifts from plant diversity to consumer control of grassland productivity. Ecol. Lett. 2022, 25, 1215–1224. [Google Scholar] [CrossRef]
- Zaret, M.M.; Kinkel, L.L.; Borer, E.T.; Seabloom, E.W. Soil nutrients cause threefold increase in pathogen and herbivore impacts on grassland plant biomass. J. Ecol. 2023, 111, 1629–1640. [Google Scholar] [CrossRef]
- Seabloom, E.W.; Borer, E.T.; Kinkel, L.L. No evidence for trade-offs in plant responses to consumer food web manipulations. Ecology 2018, 99, 1953–1963. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.Y.; Schuldt, A.; Hönig, L.; Yang, B.; Liu, X.J.; Bruelheide, H.; Ma, K.P.; Schmid, B.; Niklaus, P.A. Effects of enemy exclusion on biodiversity–productivity relationships in a subtropical forest experiment. J. Ecol. 2022, 110, 2167–2178. [Google Scholar] [CrossRef]
- Borer, E.T.; Seabloom, E.W.; Tilman, D. Plant diversity controls arthropod biomass and temporal stability. Ecol. Lett. 2012, 15, 1457–1464. [Google Scholar] [CrossRef]
- Porazinska, D.L.; Farrer, E.C.; Spasojevic, M.J.; de Mesquita, C.P.B.; Sartwell, S.A.; Smith, J.G.; White, C.T.; King, A.J.; Suding, K.N.; Schmidt, S.K. Plant diversity and density predict belowground diversity and function in an early successional alpine ecosystem. Ecology 2018, 99, 1942–1952. [Google Scholar] [CrossRef] [PubMed]
- Keesing, F.; Holt, R.D.; Ostfeld, R.S. Effects of species diversity on disease risk. Ecol. Lett. 2006, 9, 485–498. [Google Scholar] [CrossRef]
- Jing, X.; Sanders, N.J.; Shi, Y.; Chu, H.Y.; Classen, A.T.; Zhao, K.; Chen, L.T.; Shi, Y.; Jiang, Y.X.; He, J.S. The links between ecosystem multifunctionality and above- and belowground biodiversity are mediated by climate. Nat. Commun. 2015, 6, 8159. [Google Scholar] [CrossRef]
- Duffy, J.E.; Cardinale, B.J.; Mcintyre, P.B.; Thebault, E.; Loreau, M. The functional role of biodiversity in ecosystems: Incorporating trophic complexity. Ecol. Lett. 2007, 10, 522–538. [Google Scholar] [CrossRef]
- Galloway, J.N.; Dentener, F.J.; Capone, D.G.; Boyer, E.W.; Howarth, R.W.; Seitzinger, S.P.; Asner, G.P.; Cleveland, C.C.; Green, P.A.; Holland, E.A.; et al. Nitrogen cycles: Past, present, and future. Biogeochemistry 2004, 70, 153–226. [Google Scholar] [CrossRef]
- Ackerman, D.; Millet, D.B.; Chen, X. Global estimates of inorganic nitrogen deposition across four decades. Glob. Biogeochem. Cycles 2019, 33, 100–107. [Google Scholar] [CrossRef]
- Wilcots, M.E.; Schroeder, K.M.; Delancey, L.C.; Kjaer, S.J.; Hobbie, S.E.; Seabloom, E.W.; Borer, E.T. Realistic rates of nitrogen addition increase carbon flux rates but do not change soil carbon stocks in a temperate grassland. Glob. Chang. Biol. 2022, 28, 4819–4831. [Google Scholar] [CrossRef]
- Ke, Y.G.; Yu, Q.; Wang, H.Q.; Zhao, Y.; Jia, X.T.; Yang, Y.D.; Zhang, Y.L.; Zhou, W.; Wu, H.H.; Xu, C.; et al. The potential bias of nitrogen deposition effects on primary productivity and biodiversity. Glob. Chang. Biol. 2022, 29, 1054–1061. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.F.; Wu, J.G.; Clark, C.M.; Naeem, S.; Pan, Q.M.; Huang, J.H.; Zhang, L.X.; Han, X.G. Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functionsing: Evidence from inner Mongolia Grasslands. Glob. Chang. Biol. 2010, 16, 358–372. [Google Scholar] [CrossRef]
- Seabloom, E.W.; Adler, P.B.; Alberti, J.; Biederman, L.; Buckley, Y.M.; Cadotte, M.W.; Collins, S.L.; Dee, L.; Fay, P.A.; Firn, J.; et al. Increasing effects of chronic nutrient enrichment on plant diversity loss and ecosystem productivity over time. Ecology 2021, 102, e03218. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.H.; Wang, H.; Sun, J.; Niu, S.L. Global evidence on nitrogen saturation of terrestrial ecosystem net primary productivity. Environ. Res. Lett. 2016, 11, 024012. [Google Scholar] [CrossRef]
- Fridley, J.D. Resource availability dominates and alters the relationship between species diversity and ecosystem productivity in experimental plant communities. Oecologia 2002, 132, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Grime, J.P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am. Nat. 1977, 111, 1221–1226. [Google Scholar] [CrossRef]
- Frey, S.D.; Knorr, M.; Parrent, J.L.; Simpson, R.T. Chronic nitrogenenrichment affects the structure and function of the soil microbialcommunity in temperate hardwood and pine forests. Forest Ecol. Manag. 2004, 196, 159–171. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Jin, Y.H.; Xu, J.W.; He, H.S.; Tao, Y.; Yang, Z.P.; Bai, Y.Y. Effects of exogenous N and endogenous nutrients on alpine tundra litter decomposition in an area of high nitrogen deposition. Sci. Total Environ. 2021, 805, 150388. [Google Scholar] [CrossRef]
- Liu, C.C.; Li, Y.; Yan, P.; He, N.P. How to Improve the Predictions of Plant Functional Traits on ecosystem functioning? Front. Plant Sci. 2021, 12, 622260. [Google Scholar] [CrossRef]
- Tatarko, A.R.; Knops, J.M.H. Nitrogen addition and ecosystem functioning: Both species abundances and traits alter community structure and function. Ecosphere 2018, 9, e02087. [Google Scholar] [CrossRef]
- Yan, P.; He, N.P.; Yu, K.L.; Xu, L.; Van Meerbeek, K. Integrating multiple plant functional traits to predict ecosystem productivity. Commun. Biol. 2023, 6, 239. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Chen, D.; Hu, X.; Fang, X.; Li, Q.; Huang, Q.; Sun, F.; Zhou, J.; Bai, Y.; Zhang, J.; et al. Nitrogen deposition drives the intricate changes of fine root traits. Glob. Ecol. Conserv. 2023, 43, e02443. [Google Scholar] [CrossRef]
- Price, P.W. The plant vigor hypothesis and herbivore attack. Oikos 1991, 62, 244–251. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Mommer, L.; De Vries, F.T. Going belowground: Root traits as drivers of ecosystem processes. Trends Ecol. Evol. 2014, 29, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Bruno, J.F.; Boyer, K.E.; Duffy, J.E.; Lee, S.C. Relative and interactive effects of plant and grazer richness in a benthic marine community. Ecology 2008, 89, 2518–2528. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. From tropics to tundra: Global convergence in plant functioning. Proc. Natl. Acad. Sci. USA 1997, 94, 13730–13734. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Lemaire, G.; Gastal, F. Nitrogen, plant growth and crop yield. In Plant Nitrogen; Lea, P.J., Morot-Gaudry, J.F., Eds.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 343–367. [Google Scholar]
- Diaz, S.; Cabido, M. Vive la différence: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Faucon, M.P.; Houben, D.; Lambers, H. Plant functional traits: Soil and ecosystem services. Trends Plant Sci. 2017, 22, 385–394. [Google Scholar] [CrossRef]
- Zhou, Z.H.; Wang, C.K.; Zheng, M.H.; Jiang, L.F.; Luo, Y.Q. Patterns and mechanisms of response by soil microbial communities to nitrogen addition. Soil Biol. Biochem. 2017, 115, 433–441. [Google Scholar] [CrossRef]
- Averill, C.; Waring, B. Nitrogen limitation of decomposition and decay: How can it occur? Glob. Chang. Biol. 2018, 24, 1417–1427. [Google Scholar] [CrossRef]
- Han, W.J.; Chang, J.; Jiang, H.; Niu, S.D.; Liu, Y.; Xu, J.M.; Wu, J.Z.; Ge, Y. Plant species diversity affects plant nutrient pools by affecting plant biomass and nutrient concentrations in high-nitrogen ecosystems. Basic Appl. Ecol. 2021, 56, 213–225. [Google Scholar] [CrossRef]
- Tilman, D. Biodiversity: Population Versus Ecosystem Stability. Ecology 1996, 77, 350–363. [Google Scholar] [CrossRef]
- Knops, J.M.H.; Bradley, K.L.; Wedin, D.A. Mechanisms of plant species impacts on ecosystem nitrogen cycling. Ecol. Lett. 2002, 5, 454–466. [Google Scholar] [CrossRef]
- Haddad, N.M.; Tilman, D.; Haarstad, J.; Ritchie, M.; Knops, J.M.H. Contrasting effects of plant richness and composition on insect communities: A field experiment. Am. Nat. 2001, 158, 17–35. [Google Scholar] [CrossRef]
- Luo, B.; Du, Y.Y.; Han, W.J.; Geng, Y.; Wang, Q.; Duan, Y.Y.; Ren, Y.; Liu, D.; Chang, J.; Ge, Y. Reduce health damage cost of greenhouse gas and ammonia emissions by assembling plant diversity in floating constructed wetlands treating wastewater—ScienceDirect. J. Clean. Prod. 2020, 244, 118927. [Google Scholar] [CrossRef]
- Loreau, M.; Hector, A. Partitioning selection and complementarity in biodiversity experiments. Nature 2001, 412, 72–76. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Aboveground Biomass | Belowground Biomass | Total Biomass | |||
|---|---|---|---|---|---|---|
| p Value | Change | p Value | Change | p Value | Change | |
| Control | ||||||
| P. frutescens | 0.223 | ns | 0.135 | ns | 0.061 | ns |
| B. striata | 0.357 | ns | <0.001 | ↑ 381.67% | 0.058 | ns |
| B. pilosa | <0.001 | ↑ 343.78% | 0137 | ns | 0.818 | ns |
| Insecticide | ||||||
| P. frutescens | 0.121 | ns | 0.109 | ns | 0.003 | ↓ 49.72% |
| B. striata | 0.942 | ns | <0.001 | ↑ 356.69% | 0.071 | ns |
| B. pilosa | <0.001 | ↑ 300.77% | 0.216 | ns | 0.189 | ns |
| Fungicide | ||||||
| P. frutescens | 0.554 | ns | 0.152 | ns | 0.126 | ns |
| B. striata | 0.102 | ns | <0.001 | ↑ 252.87% | 0.645 | ns |
| B. pilosa | <0.001 | ↑ 472.00% | 1.000 | ns | 0.001 | ↑ 110.19% |
| Insecticide + Fungicide | ||||||
| P. frutescens | 0.567 | ns | 0.056 | ns | 0.144 | ns |
| B. striata | 0.397 | ns | 0.003 | ↑ 138.16% | 0.765 | ns |
| B. pilosa | <0.001 | ↑ 506.20% | 0.635 | ns | <0.001 | ↑ 155.78% |
| Source of Variation | Aboveground Biomass | Belowground Biomass | Total Biomass | |||
|---|---|---|---|---|---|---|
| p Value | Change | p Value | Change | p Value | Change | |
| Control | ||||||
| P. frutescens | 0.235 | ns | 0.487 | ns | 0.095 | ns |
| B. striata | 0.016 | ↓ 58.09% | <0.001 | ↑ 170.89% | 0.357 | ns |
| B. pilosa | <0.001 | ↑ 735.36% | 0.040 | ↓ 46.65% | 0.002 | ↑ 122.13% |
| Insecticide | ||||||
| P. frutescens | 0.247 | ns | 0.002 | ↓ 75.01% | 0.009 | ↓ 52.30% |
| B. striata | 0.111 | ns | 0.178 | ns | 0.114 | ns |
| B. pilosa | <0.001 | ↑ 425.05% | 0.516 | ns | <0.001 | ↑ 220.75% |
| Fungicide | ||||||
| P. frutescens | 0.316 | ns | 0.134 | ns | 0.069 | ns |
| B. striata | 0.464 | ns | 0.018 | ↑ 174.19% | 0.555 | ns |
| B. pilosa | <0.001 | ↑ 270.44% | 0.237 | ns | 0.003 | ↑ 85.24% |
| Insecticide + Fungicide | ||||||
| P. frutescens | 0.432 | ns | 0.014 | ↓ 60.88% | 0.980 | ns |
| B. striata | 0.077 | ns | 0.236 | ns | 0.148 | ns |
| B. pilosa | <0.001 | ↑ 355.43% | 0.347 | ns | <0.001 | ↑ 211.94% |
| Source of Variation | Leaf Area | Root Length | Root Diameter | Root Tip Number | ||||
|---|---|---|---|---|---|---|---|---|
| p Value | Change | p Value | Change | p Value | Change | p Value | Change | |
| Control | ||||||||
| P. frutescens | 0.136 | ns | 0.063 | ns | 0.208 | ns | 0.007 | ↑ 62.01% |
| B. striata | <0.001 | ↑ 95.19% | 0.002 | ↓ 52.62% | 0.012 | ↑ 51.69% | 0.016 | ↓ 37.89% |
| B. pilosa | 0.133 | ns | 0.341 | ns | 0.037 | ↓ 25.67% | 0.138 | ns |
| Insecticide | ||||||||
| P. frutescens | 0.123 | ns | 0.311 | ns | 0.035 | ↓ 21.59% | 0.480 | ns |
| B. striata | 0.006 | ↑ 96.78% | <0.001 | ↓ 40.80% | <0.001 | ↑ 53.61% | 0.010 | ↓ 34.62% |
| B. pilosa | 0.004 | ↓ 57.27% | 0.235 | ns | 0.494 | ns | 0.589 | ns |
| Fungicide | ||||||||
| P. frutescens | 0.177 | ns | 0.180 | ns | 0.255 | ns | 0.032 | ↓ 25.80% |
| B. striata | 0.002 | ↑ 129.53% | 0.413 | ns | <0.001 | ↑ 59.98% | 0.432 | ns |
| B. pilosa | 0.368 | ns | 0.036 | ↑ 41.28% | 0.619 | ns | 0.378 | ns |
| Insecticide + Fungicide | ||||||||
| P. frutescens | 0.744 | ns | 0.119 | ns | 0.268 | ns | 0.052 | ns |
| B. striata | 0.002 | ↑ 81.35% | 0.135 | ns | <0.001 | ↑ 51.78% | 0.039 | ↓ 21.53% |
| B. pilosa | 0.275 | ns | 0.876 | ns | 0.109 | ns | 0.445 | ns |
| Source of Variation | Leaf Area | Root Length | Root Diameter | Root Tip Number | ||||
|---|---|---|---|---|---|---|---|---|
| p Value | Change | p Value | Change | p Value | Change | p Value | Change | |
| Control | ||||||||
| P. frutescens | 0.111 | ns | 0.542 | ns | 0.693 | ns | 0.913 | ns |
| B. striata | 0.025 | ↑ 55.64% | 0.007 | ↓ 53.30% | <0.001 | ↑ 73.57% | <0.001 | ↓ 48.59% |
| B. pilosa | 0.762 | ns | <0.001 | ↑ 225.29% | <0.001 | ↓ 36.05% | 0.001 | ↑ 90.01% |
| Insecticide | ||||||||
| P. frutescens | 0.023 | ↓ 34.55% | 0.865 | ns | 0.077 | ns | 0.678 | ns |
| B. striata | 0.090 | ns | 0.008 | ↓ 64.61% | <0.001 | ↑ 67.27% | <0.001 | ↓ 41.13% |
| B. pilosa | 0.664 | ns | 0.018 | ↑ 79.98% | 0.900 | ns | 0.667 | ns |
| Fungicide | ||||||||
| P. frutescens | 0.180 | ns | 0.824 | ns | 0.406 | ns | 0.826 | ns |
| B. striata | 0.832 | ns | 0.014 | ↓ 36.42% | <0.001 | ↑ 62.31% | 0.048 | ↓ 34.31% |
| B. pilosa | 0.781 | ns | 0.003 | ↑ 94.93% | 0.046 | ↓ 27.22% | 0.239 | ns |
| Insecticide + Fungicide | ||||||||
| P. frutescens | 0.273 | ns | 0.432 | ns | 0.252 | ns | 0.622 | ns |
| B. striata | 0.810 | ns | <0.001 | ↓ 64.46% | <0.001 | ↑ 90.17% | <0.001 | ↓ 57.71% |
| B. pilosa | 0.026 | ↓ 26.22% | 0.015 | ↑ 76.82% | 0.164 | ns | 0.180 | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Yang, L.; Shen, K.; Cao, H.; Lin, Y.; Liu, J.; Han, W. Nitrogen Addition and Heterotroph Exclusion Affected Plant Species Diversity–Biomass Relationship by Affecting Plant Functional Traits. Plants 2024, 13, 258. https://doi.org/10.3390/plants13020258
Xu X, Yang L, Shen K, Cao H, Lin Y, Liu J, Han W. Nitrogen Addition and Heterotroph Exclusion Affected Plant Species Diversity–Biomass Relationship by Affecting Plant Functional Traits. Plants. 2024; 13(2):258. https://doi.org/10.3390/plants13020258
Chicago/Turabian StyleXu, Xile, Luping Yang, Kai Shen, Huijuan Cao, Yishi Lin, Jinliang Liu, and Wenjuan Han. 2024. "Nitrogen Addition and Heterotroph Exclusion Affected Plant Species Diversity–Biomass Relationship by Affecting Plant Functional Traits" Plants 13, no. 2: 258. https://doi.org/10.3390/plants13020258
APA StyleXu, X., Yang, L., Shen, K., Cao, H., Lin, Y., Liu, J., & Han, W. (2024). Nitrogen Addition and Heterotroph Exclusion Affected Plant Species Diversity–Biomass Relationship by Affecting Plant Functional Traits. Plants, 13(2), 258. https://doi.org/10.3390/plants13020258