
Genome-Wide Identification of Expansins in Rubus chingii and Profiling Analysis during Fruit Ripening and Softening

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Identification of Expansin Family Members in Rubus chingii
2.2. Gene Structures and Protein Motifs of RchEXPs
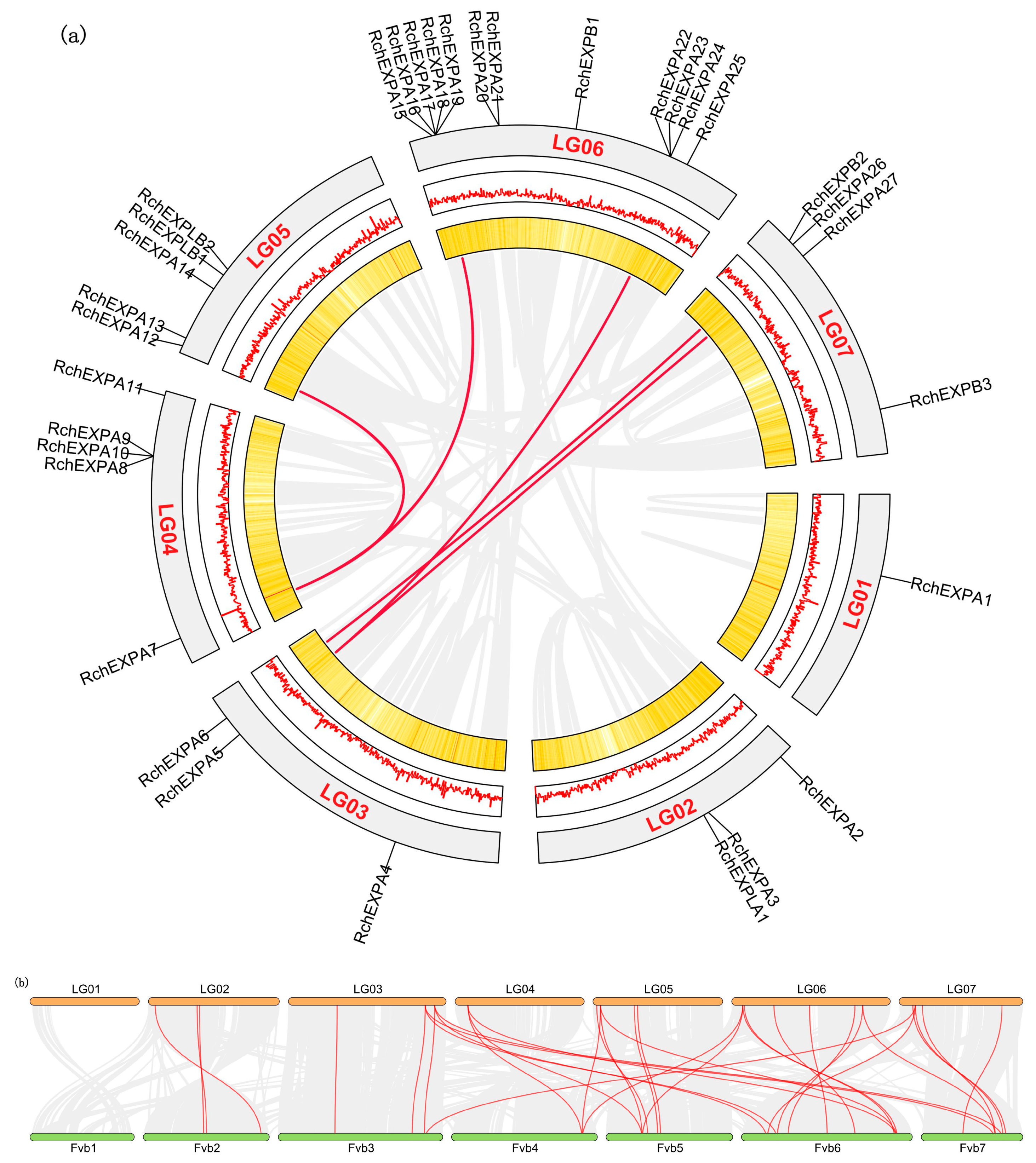
2.3. Chromosomal Location and Synteny Analyses of RchEXPs
2.4. Cis-Regulatory Elements and Interaction Analyses of RchEXPs
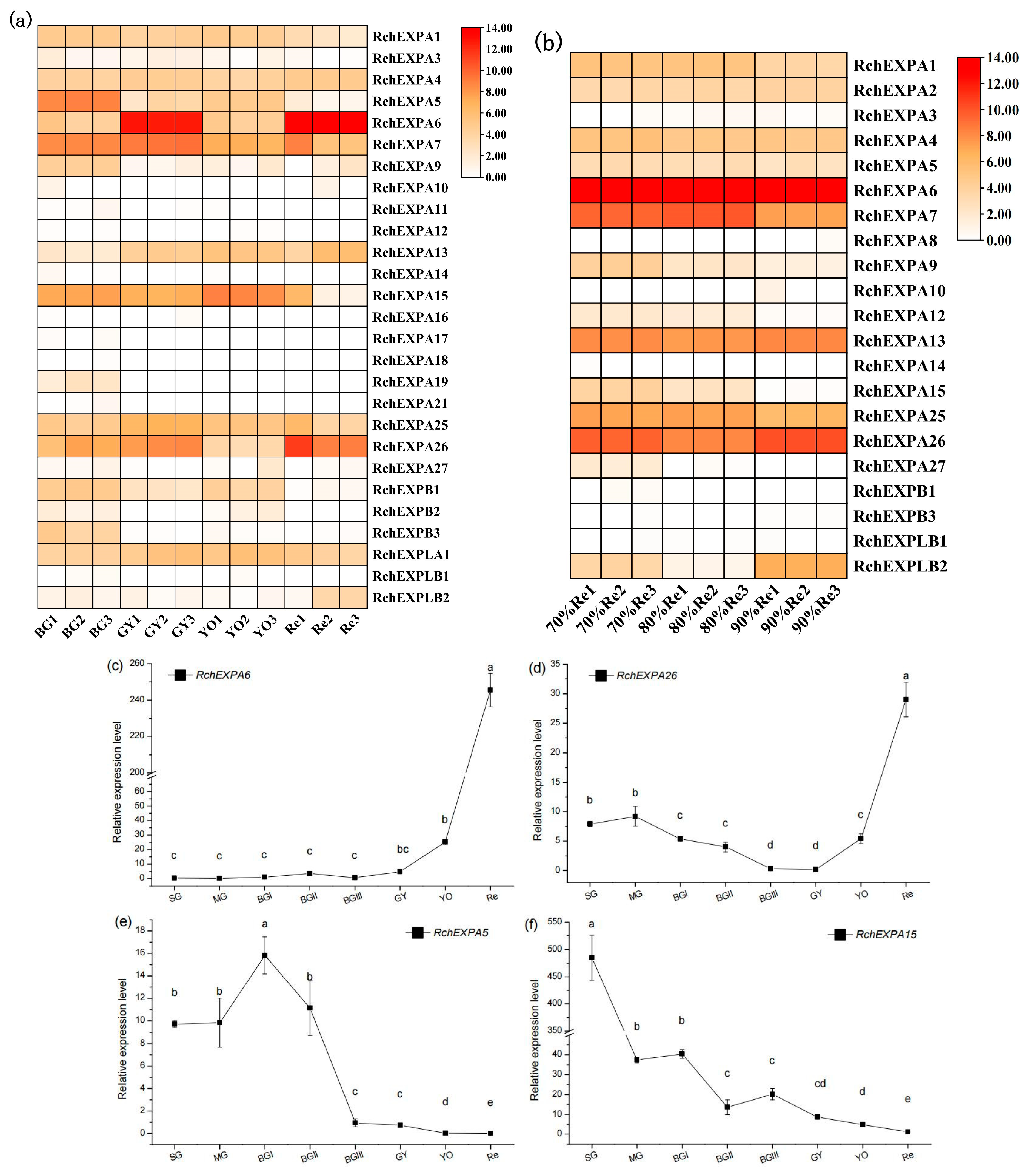
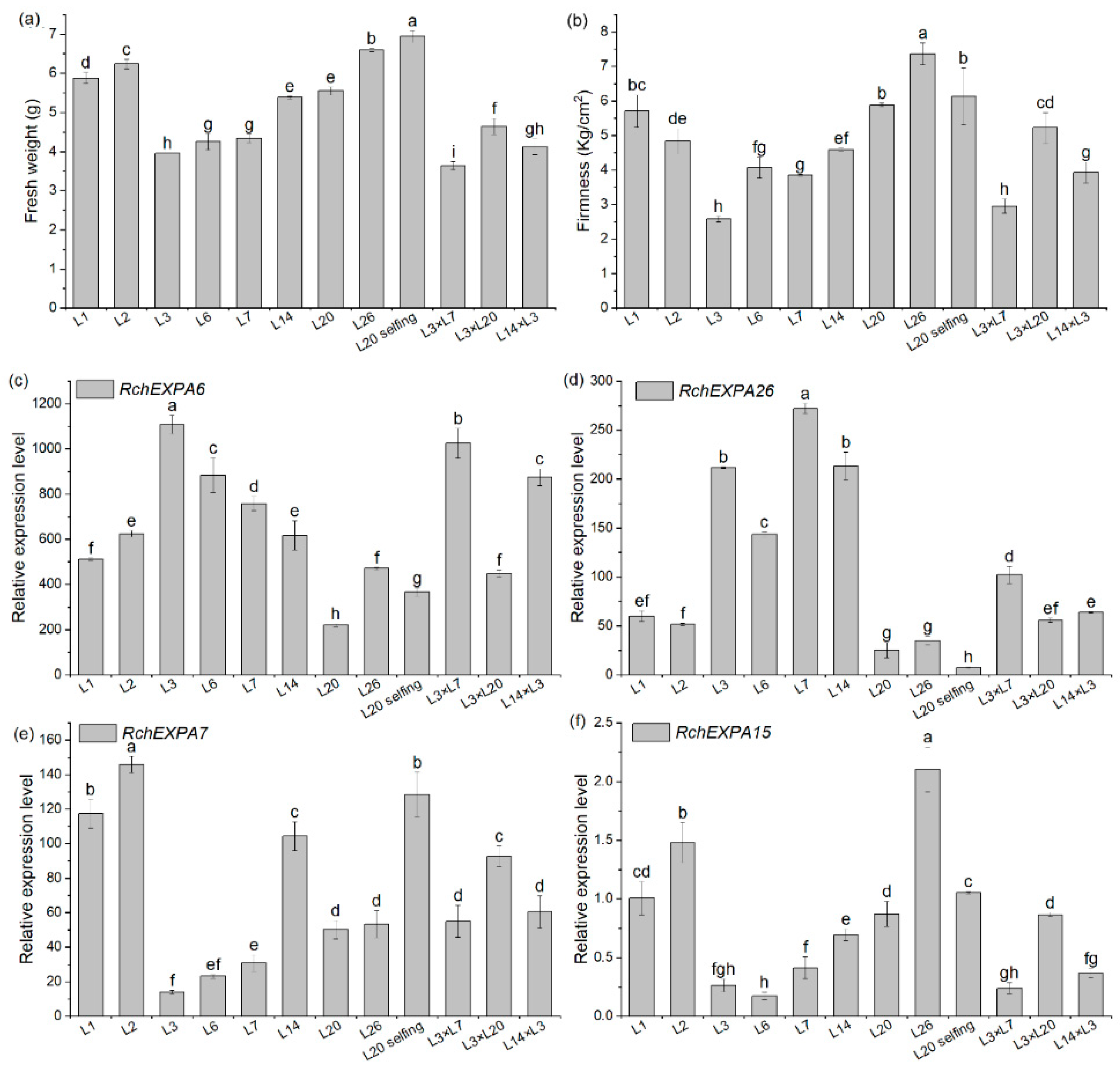
2.5. Expression Profiles of RchEXPs in Different Fruit Ripening and Softening Stages of R. chingii
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material
5.2. Genome-Wide Identification of Expansin Family in R. chingii and Phylogenetic Analysis
5.3. Motifs, Gene Structures, Chromosomal Location, and Synteny Analysis of Expansins in R. chingii
5.4. Cis-Regulatory Elements Analysis and Protein–Protein Interaction Network Construction of Expansins in R. chingii
5.5. Expression Analyses of RchExps during Developmental Stages
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sheng, J.Y.; Wang, S.Q.; Liu, K.H.; Zhu, B.; Zhang, Q.Y.; Qin, L.P.; Wu, J.J. Rubus chingii Hu: An overview of botany, traditional uses, phytochemistry, and pharmacology. Chin. J. Nat. Med. 2020, 18, 401–416. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Jiang, J.; Li, X.; Xie, Y.; Jin, Z.; Wang, X.; Li, Y.; Zhong, Y.; Lin, J.; Yang, W. Bioactive compounds and fruit quality of Chinese raspberry, Rubus chingii Hu varied with genotype and phenological phase. Sci. Hortic. 2021, 281, 109951. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia Commission. Fupenzi, Rubi Fructus. Pharmacopoeia of the People’s Republic of China (Edition 2020); China Medical Science Press: Beijing, China, 2020. [Google Scholar]
- Yu, G.; Luo, Z.; Wang, W.; Li, Y.; Zhou, Y.; Shi, Y. Rubus chingii Hu: A review of the phytochemistry and pharmacology. Front. Pharmacol. 2019, 10, 799. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Jiang, J.; Shu, L.; Li, X.; Huang, J.; Qian, B.; Wang, X.; Li, X.; Chen, J.; Xu, H. Combined transcriptomic and metabolic analyses reveal potential mechanism for fruit development and quality control of Chinese raspberry (Rubus chingii Hu). Plant Cell Rep. 2021, 40, 1923–1946. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lei, T.; Han, G.; Yue, J.; Zhang, X.; Yang, Q.; Ruan, H.; Gu, C.; Zhang, Q.; Qian, T.; et al. The chromosome-scale reference genome of Rubus chingii Hu provides insight into the biosynthetic pathway of hydrolyzable tannins. Plant J. 2021, 107, 1466–1477. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Hu, Y.; Li, J.; Cai, J.; Qiu, Y.; Dong, C. Anti-inflammatory effect of a novel pectin polysaccharide from Rubus chingii Hu on colitis mice. Front. Nutr. 2022, 9, 868657. [Google Scholar] [CrossRef]
- Wang, J.; Xiao, X.; Zhou, N.; Zhao, M.; Lang, S.; Ren, Q.; Wang, D.; Fu, H. Rubochingosides A—J, labdane-type diterpene glycosides from leaves of Rubus chingii. Phytochemistry 2023, 210, 113670. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, D.; Zhu, B.; Wang, S.; Xu, Y.; Zhang, C.; Yang, H.; Wang, S.; Liu, P.; Qin, L.; et al. Rubus chingii Hu. unripe fruits extract ameliorates carbon tetrachloride-induced liver fibrosis and improves the associated gut microbiota imbalance. Chin. Med. 2022, 17, 56. [Google Scholar] [CrossRef]
- Zhong, J.; Wang, Y.; Li, C.; Yu, Q.; Xie, J.; Dong, R.; Xie, Y.; Li, B.; Tian, J.; Chen, Y. Natural variation on free, esterified, glycosylated and insoluble-bound phenolics of Rubus chingii Hu: Correlation between phenolic constituents and antioxidant activities. Food Res. Int. 2022, 162, 112043. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Z.; Guo, Q.; Gao, X.; Ma, Q.; Xue, Z.; Ferri, N.; Zhang, M.; Chen, H. Identification of ellagitannins in the unripe fruit of Rubus chingii Hu and evaluation of its potential antidiabetic activity. J. Agric. Food Chem. 2019, 67, 7025–7039. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y.; Jin, L.; Chen, Z.; Jiang, J.; Jackson, A. Development of fruit color in Rubus chingii Hu (Chinese raspberry): A story about novel offshoots of anthocyanin and carotenoid biosynthesis. Plant Sci. 2021, 311, 110996. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.M.; Bassil, N.V.; Dossett, M.; Leigh Worthington, M.; Graham, J. Genetic and genomic resources for Rubus breeding: A roadmap for the future. Hortic. Res. 2019, 6, 116. [Google Scholar] [CrossRef]
- Li, B.J.; Grierson, D.; Shi, Y.; Chen, K.S. Roles of abscisic acid in regulating ripening and quality of strawberry, a model non-climacteric fruit. Hortic. Res. 2022, 9, uhac089. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Manghwar, H.; Hu, W. Study on supergenus Rubus L.: Edible, medicinal, and phylogenetic characterization. Plants 2022, 11, 1211. [Google Scholar] [CrossRef]
- VanBuren, R.; Bryant, D.; Bushakra, J.M.; Vining, K.J.; Edger, P.P.; Rowley, E.R.; Priest, H.D.; Michael, T.P.; Lyons, E.; Filichkin, S.A.; et al. The genome of black raspberry (Rubus occidentalis). Plant J. 2016, 87, 535–547. [Google Scholar] [CrossRef]
- Hua, Y.; Dai, B.; Luo, Y.; Ding, Y. Integrated analysis of multiple metabolome and transcriptome revealed the accumulation of flavonoids and associated molecular regulation mechanisms in Rubus chingii Hu at different developmental stages. Plant Physiol. Biochem. 2023, 204, 108085. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Huang, J.; Ruan, H.; Qian, W.; Fang, Z.; Gu, C.; Zhang, N.; Liang, Y.; Wang, Z.; Gao, L.; et al. Competition between FLS and DFR regulates the distribution of flavonols and proanthocyanidins in Rubus chingii Hu. Front. Plant Sci. 2023, 14, 1134993. [Google Scholar] [CrossRef]
- Valenzuela-Riffo, F.; Parra-Palma, C.; Ramos, P.; Morales-Quintana, L. Molecular and structural insights into FaEXPA5, an alpha-expansin protein related with cell wall disassembly during ripening of strawberry fruit. Plant Physiol. Biochem. 2020, 154, 581–589. [Google Scholar] [CrossRef]
- Ren, Y.; Li, B.; Jia, H.; Yang, X.; Sun, Y.; Shou, J.; Jiang, G.; Shi, Y.; Chen, K. Comparative analysis of fruit firmness and genes associated with cell wall metabolisms in three cultivated strawberries during ripening and postharvest. Food Qual. Saf. 2023, 7, fyad020. [Google Scholar] [CrossRef]
- Mu, Q.; Li, X.; Luo, J.; Pan, Q.; Li, Y.; Gu, T. Characterization of expansin genes and their transcriptional regulation by histone modifications in strawberry. Planta 2021, 254, 21. [Google Scholar] [CrossRef]
- Jin, K.M.; Zhuo, R.Y.; Xu, D.; Wang, Y.J.; Fan, H.J.; Huang, B.Y.; Qiao, G.R. Genome-wide identification of the expansin gene family and its potential association with drought stress in moso bamboo. Int. J. Mol. Sci. 2020, 21, 9491. [Google Scholar] [CrossRef] [PubMed]
- Sampedro, J.; Cosgrove, D.J. The expansin superfamily. Genome Biol. 2005, 6, 242. [Google Scholar] [CrossRef] [PubMed]
- Marowa, P.; Ding, A.; Kong, Y. Expansins: Roles in plant growth and potential applications in crop improvement. Plant Cell Rep. 2016, 35, 949–965. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Zou, X.; Gao, Q. Genome-wide identification of expansin in Fragaria vesca and expression profiling analysis of the FvEXPs in different fruit development. Gene 2022, 814, 146162. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Plant expansins: Diversity and interactions with plant cell walls. Curr. Opin. Plant Biol. 2015, 25, 162–172. [Google Scholar] [CrossRef]
- Samalova, M.; Gahurova, E.; Hejatko, J. Expansin-mediated developmental and adaptive responses: A matter of cell wall biomechanics? Quant. Plant Biol. 2022, 13, e11. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, B.; Li, C.; Lei, C.; Kong, C.; Yang, Y.; Gong, M. A comprehensive expression analysis of the expansin gene family in potato (Solanum tuberosum) discloses stress-responsive expansin-like B genes for drought and heat tolerances. PLoS ONE 2019, 14, e0219837. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, G.; An, J.; Li, Q.; Chen, Y.; Zhao, X.; Wu, J.; Wang, Y.; Hao, Q.; Wang, W. Expansin gene TaEXPA2 positively regulates drought tolerance in transgenic wheat (Triticum aestivum L.). Plant Sci. 2020, 298, 110596. [Google Scholar] [CrossRef]
- Bashline, L.; Lei, L.; Li, S.; Gu, Y. Cell wall, cytoskeleton, and cell expansion in higher plants. Mol. Plant 2014, 7, 586–600. [Google Scholar] [CrossRef]
- Culbertson, A.T.; Smith, A.L.; Cook, M.D.; Zabotina, O.A. Truncations of xyloglucan xylosyltransferase 2 provide insights into the roles of the N- and C-terminus. Phytochemistry 2016, 128, 12–19. [Google Scholar] [CrossRef]
- Wang, T.; Park, Y.B.; Caporini, M.A.; Rosay, M.; Zhong, L.; Cosgrove, D.J.; Hong, M. Sensitivity-enhanced solid-state NMR detection of expansin’s target in plant cell walls. Proc. Natl. Acad. Sci. USA 2013, 110, 16444–16449. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Chen, F.; Luo, J.; Zeng, W.; Xu, W. The biosynthesis of plant cell wall xylan and its application. Chin. Bull. Bot. 2023, 58, 316–334. (In Chinese) [Google Scholar] [CrossRef]
- Minoia, S.; Boualem, A.; Marcel, F.; Troadec, C.; Quemener, B.; Cellini, F.; Petrozza, A.; Vigouroux, J.; Lahaye, M.; Carriero, F.; et al. Induced mutations in tomato SlExp1 alter cell wall metabolism and delay fruit softening. Plant Sci. 2016, 242, 195–202. [Google Scholar] [CrossRef]
- Dotto, M.C.; Martínez, G.A.; Civello, P.M. Expression of expansin genes in strawberry varieties with contrasting fruit firmness. Plant Physiol. Biochem. 2006, 44, 301–307. [Google Scholar] [CrossRef]
- Valenzuela-Riffo, F.; Gaete-Eastman, C.; Stappung, Y.; Lizana, R.; Herrera, R.; Moya-León, M.A.; Morales-Quintana, L. Comparative in silico study of the differences in the structure and ligand interaction properties of three alpha-expansin proteins from Fragaria chiloensis fruit. J. Biomol. Struct. Dyn. 2019, 37, 3245–3258. [Google Scholar] [CrossRef]
- Figueroa, N.E.; Gatica-Meléndez, C.; Figueroa, C.R. Ethylene application at the immature stage of Fragaria chiloensis fruit represses the anthocyanin biosynthesis with a concomitant accumulation of lignin. Food Chem 2021, 358, 129913. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Chromosome Location | Length (aa) | pI | MW | Subcellular Ocalization |
|---|---|---|---|---|---|---|
| LG01.812 | RchEXPA1 | LG01:8,877,139–8,884,215 | 361 | 6.99 | 39,910.71 | Cell wall |
| LG02.327 | RchEXPA2 | LG02:1,544,766–1,543,798 | 256 | 8.86 | 27,373.94 | Cell wall |
| LG02.2248 | RchEXPA3 | LG02:11,412,133–11,413,361 | 252 | 8.00 | 27,683.61 | Cell wall |
| LG03.2086 | RchEXPA4 | LG03:11,399,062–11,406,813 | 482 | 9.30 | 53,196.43 | Cell wall |
| LG03.4509 | RchEXPA5 | LG03:32,076,184–32,078,242 | 259 | 9.60 | 27,888.73 | Cell wall |
| LG03.4886 | RchEXPA6 | LG03:34,333,208–34,334,320 | 253 | 8.12 | 27,197.39 | Cell wall |
| LG04.560 | RchEXPA7 | LG04:2,963,719–2,975,241 | 327 | 8.83 | 35,055.07 | Cell wall |
| LG04.3299 | RchEXPA8 | LG04:23,036,026–23,036,555 | 125 | 7.55 | 12,970.17 | Cell wall |
| LG04.3300 | RchEXPA9 | LG04:23,053,077–23,054,497 | 133 | 5.03 | 13,924.83 | Cell wall |
| LG04.3301 | RchEXPA10 | LG04:23,039,084–23,040,442 | 132 | 8.16 | 13,990.05 | Cell wall |
| LG04.4234 | RchEXPA11 | LG04:29,721,537–29,722,605 | 252 | 7.52 | 27,668.67 | Cell wall |
| LG05.148 | RchEXPA12 | LG05:803,994–804,890 | 235 | 9.02 | 26,237.85 | Cell wall |
| LG05.336 | RchEXPA13 | LG05:1,698,742–1,700,885 | 289 | 9.49 | 31,609.97 | Cell wall |
| LG05.1456 | RchEXPA14 | LG05:7,815,734–7,817,299 | 261 | 9.32 | 29,004.09 | Cell wall |
| LG06.511 | RchEXPA15 | LG06:2,552,263–2,555,905 | 273 | 9.37 | 30,024.84 | Cell wall |
| LG06.587 | RchEXPA16 | LG06:2,934,998–2,935,958 | 262 | 9.00 | 29,008.40 | Cell wall |
| LG06.588 | RchEXPA17 | LG06:2,937,934–2,938,949 | 252 | 8.09 | 27,558.47 | Cell wall |
| LG06.589 | RchEXPA18 | LG06:2,940,922–2,941,932 | 252 | 7.55 | 27,639.52 | Cell wall |
| LG06.590 | RchEXPA19 | LG06:2,943,293–2,944,142 | 250 | 8.80 | 27,593.51 | Cell wall |
| LG06.1815 | RchEXPA20 | LG06:9,876,023–9,877,025 | 250 | 8.01 | 26,551.37 | Cell wall |
| LG06.1816 | RchEXPA21 | LG06:9,877,714–9,879,031 | 252 | 8.65 | 26,995.71 | Cell wall |
| LG06.4033 | RchEXPA22 | LG06:28,894,710–28,897,718 | 302 | 9.07 | 33,583.85 | Cell wall |
| LG06.4034 | RchEXPA23 | LG06:28,900,123–28,902,589 | 271 | 5.77 | 30,149.98 | Cell wall |
| LG06.4035 | RchEXPA24 | LG06:28,905,229–28,906,521 | 264 | 8.75 | 29,355.39 | Cell wall |
| LG06.4347 | RchEXPA25 | LG06:30,752,267–30,753,827 | 260 | 9.46 | 27,967.92 | Cell wall |
| LG07.721 | RchEXPA26 | LG07:3,727,233–3,728,227 | 252 | 7.53 | 26,779.83 | Cell wall |
| LG07.1027 | RchEXPA27 | LG07:5,399,695–5,401,139 | 258 | 9.01 | 27,605.21 | Cell wall |
| LG06.2871 | RchEXPB1 | LG06:18,236,794–18,237,946 | 261 | 8.98 | 28,257.54 | Cell wall |
| LG07.600 | RchEXPB2 | LG07:3,173,494–3,174,761 | 270 | 5.35 | 28,845.35 | Cell wall |
| LG07.3409 | RchEXPB3 | LG07:24,111,895–24,113,983 | 286 | 4.39 | 30,073.51 | Cell wall |
| LG02.2351 | RchEXPLA1 | LG02:11,988,434–11,989,907 | 260 | 8.45 | 28,204.14 | Cell wall |
| LG05.1739 | RchEXPLB1 | LG05:9,646,051–9,653,623 | 247 | 6.87 | 27,091.51 | Cell wall |
| LG05.1872 | RchEXPLB2 | LG05:10,406,187–10,407,678 | 257 | 4.88 | 28,062.50 | Cell wall |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Shen, D.; Shi, Y.; Chen, Y.; He, H.; Jiang, J.; Wang, F.; Jiang, J.; Wang, X.; Li, X.; et al. Genome-Wide Identification of Expansins in Rubus chingii and Profiling Analysis during Fruit Ripening and Softening. Plants 2024, 13, 431. https://doi.org/10.3390/plants13030431
Chen Z, Shen D, Shi Y, Chen Y, He H, Jiang J, Wang F, Jiang J, Wang X, Li X, et al. Genome-Wide Identification of Expansins in Rubus chingii and Profiling Analysis during Fruit Ripening and Softening. Plants. 2024; 13(3):431. https://doi.org/10.3390/plants13030431
Chicago/Turabian StyleChen, Zhen, Danwei Shen, Yujie Shi, Yiquan Chen, Honglian He, Junfeng Jiang, Fan Wang, Jingyong Jiang, Xiaoyan Wang, Xiaobai Li, and et al. 2024. "Genome-Wide Identification of Expansins in Rubus chingii and Profiling Analysis during Fruit Ripening and Softening" Plants 13, no. 3: 431. https://doi.org/10.3390/plants13030431
APA StyleChen, Z., Shen, D., Shi, Y., Chen, Y., He, H., Jiang, J., Wang, F., Jiang, J., Wang, X., Li, X., & Zeng, W. (2024). Genome-Wide Identification of Expansins in Rubus chingii and Profiling Analysis during Fruit Ripening and Softening. Plants, 13(3), 431. https://doi.org/10.3390/plants13030431