


Chemodiversity of Arctic Plant Dryas oxyodonta: LC-MS Profile and Antioxidant Activity




Abstract
:1. Introduction
2. Results and Discussion
2.1. Metabolome of Dryas oxyodonta
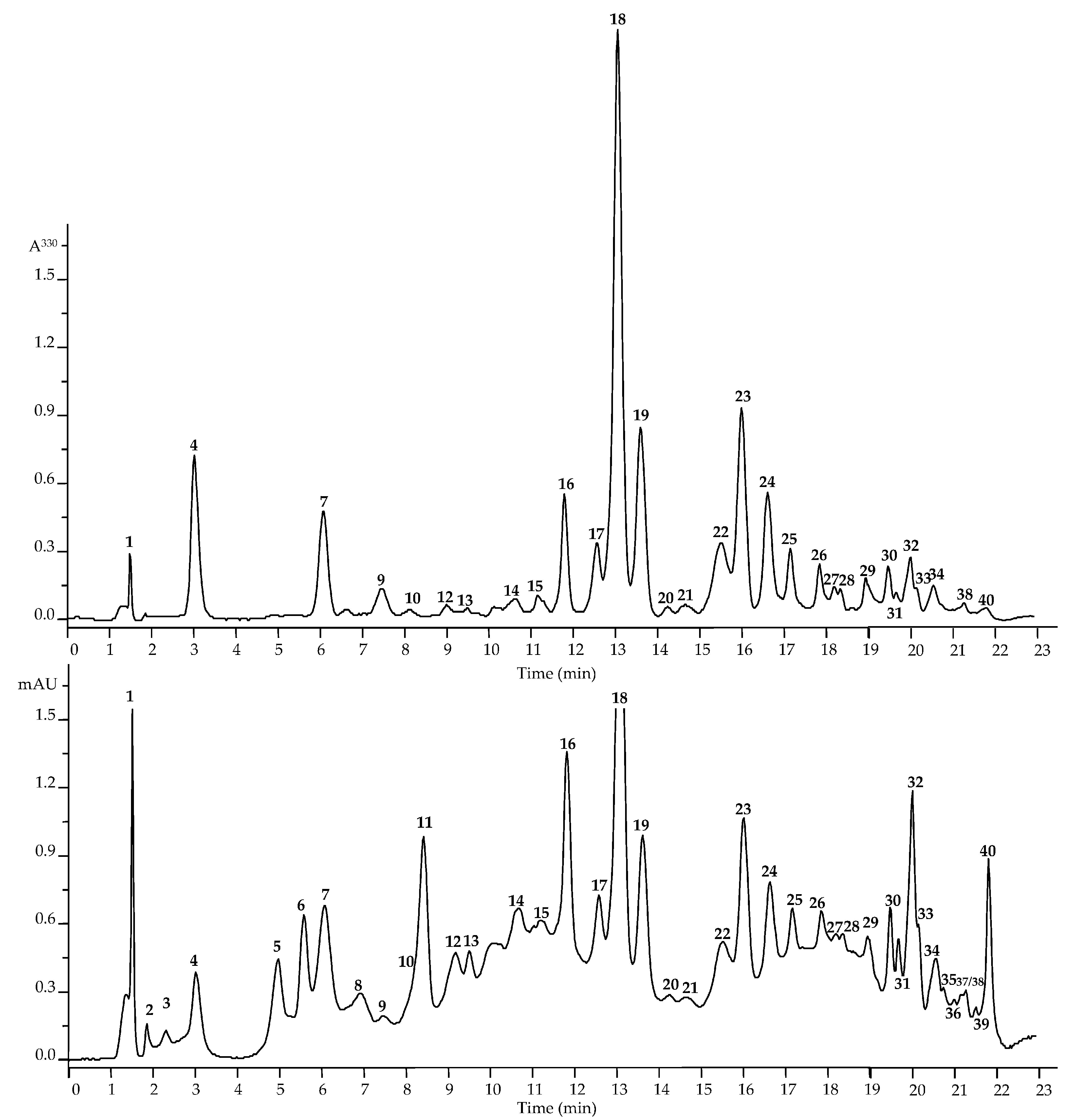
2.1.1. Gallotannins, Hydroxycinnamates, Procyanidins, and Catechins
2.1.2. Flavonoids
2.1.3. Triterpenes
2.2. Chemodiversity Significance of D. oxyodonta Metabolites for the Dryas Genus
2.3. Quantitative Analysis of Metabolites in Dryas oxyodonta Extracts
2.4. Antioxidant Activity of D. oxyodonta Extracts
3. Materials and Methods
3.1. Plant Material
3.2. Reagents
3.3. Extract Preparation
3.4. Liquid Chromatography–Mass Spectrometry Detection of Metabolites in D. oxyodonta Extracts
3.5. HPLC-PDA-ESI-tQ-MS Quantification of Metabolites in D. oxyodonta Extracts
3.6. Antioxidant Activity of D. oxyodonta Extracts
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Challice, J.S. Rosaceae chemotaxonomy and the origins of the Pomoideae. Bot. J. Linn. Soc. 1974, 69, 239–259. [Google Scholar] [CrossRef]
- Takhtajan, A. Diversity and Classification of Flowering Plants; Columbia University Press: New York, NY, USA, 1997; pp. 1–620. [Google Scholar]
- Shulaev, V.; Korban, S.S.; Sosinski, B.; Abbott, A.G.; Aldwinckle, H.S.; Folta, K.M.; Iezzoni, A.; Main, D.; Arús, P.; Dandekar, A.M.; et al. Multiple models for Rosaceae genomics. Plant Physiol. 2008, 147, 985–1003. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Hatano, T.; Iwasaki, M.; Kubo, M.; Orime, T.; Yoshizaki, M.; Naruhashi, N. Hydrolysable tannins as chemotaxonomic markers in the Rosaceae. Phytochemistry 1992, 31, 3091–3096. [Google Scholar] [CrossRef]
- Potter, D.; Eriksson, T.; Evans, R.C.; Oh, S.; Smedmark, J.E.E.; Morgan, D.R.; Kerr, M.; Robertson, K.R.; Arsenault, M.; Dickinson, T.A.; et al. Phylogeny and classification of Rosaceae. Plant Syst. Evol. 2007, 266, 5–43. [Google Scholar] [CrossRef]
- Watson, L.; Dallwitz, M.J. The families of flowering plants. Interactive identification and information retrieval on CD-ROM version 1.0 1993, and colour illustrated manual. Nord. J. Bot. 1994, 14, 486. [Google Scholar]
- Sedelnikov, V.P. High-mountain vegetation of North Asia: Dryad tundras. Contemp. Probl. Ecol. 2015, 8, 267–277. [Google Scholar] [CrossRef]
- McGraw, J.B.; Antanovics, J. Experimental ecology of Dryas octopetala ecotypes: I. Ecotypic differentiation and life cycle stages of selection. J. Ecol. 1983, 71, 879–897. [Google Scholar] [CrossRef]
- Komarov, V.L. Flora of USSR; AN SSSR: Moscow, Russia, 1941; Volume 10, p. 273. [Google Scholar]
- Plants of the World Online. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:724676-1 (accessed on 12 March 2024).
- Makarov, A.A. Plant Medical Remedies of Yakut Traditional Medicine; YaGU: Yakutsk, Russia, 1974; pp. 26–47. [Google Scholar]
- Batorova, S.M.; Yakovlev, G.P.; Aseeva, T.A. Reference-Book of Traditional Tibetan Medicine Herbs; Nauka: Novosibirsk, Russia, 2013; pp. 84–95. [Google Scholar]
- Pangon, J.F.; Jay, M.; Voirin, B. Les flavonoides du Dryas octopetala. Phytochemistry 1974, 13, 1883–1885. [Google Scholar] [CrossRef]
- Servettaz, O.; Colombo, M.L.; De Bernardi, M.; Uberti, E.; Vidari, G.; Vita-Finzi, P. Flavonol glycosides from Dryas octopetala. J. Nat. Prod. 1984, 47, 809–814. [Google Scholar] [CrossRef]
- Petelka, J.; Plagg, B.; Säumel, I.; Zerbe, S. Traditional medicinal plants in South Tyrol (northern Italy, southern Alps): Biodiversity and use. J. Ethnobiol. Ethnomed. 2020, 16, 74. [Google Scholar] [CrossRef]
- Manninen, M.; Karonen, M.; Salminen, J.-P. Chemotaxonomic markers for the leaf buds of common Finnish trees and shrubs: A rapid UHPLC MS fingerprinting tool for species identification. Molecules 2022, 27, 6810. [Google Scholar] [CrossRef]
- Kashchenko, N.I.; Olennikov, D.N. Phenolome of Asian agrimony tea (Agrimonia asiatica Juz., Rosaceae): LC-MS profile, α-glucosidase inhibitory potential and stability. Foods 2020, 9, 1348. [Google Scholar] [CrossRef]
- Kashchenko, N.I.; Olennikov, D.N.; Chirikova, N.K. Metabolites of Siberian raspberries: LC-MS profile, seasonal variation, antioxidant activity and, thermal stability of Rubus matsumuranus phenolome. Plants 2021, 10, 2317. [Google Scholar] [CrossRef] [PubMed]
- Kashchenko, N.I.; Olennikov, D.N.; Chirikova, N.K. Metabolites of Geum aleppicum and Sibbaldianthe bifurca: Diversity and α-glucosidase inhibitory potential. Metabolites 2023, 13, 689. [Google Scholar] [CrossRef] [PubMed]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis: Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Vallverdú-Queralt, A.; Jáuregui, O.; Medina-Remón, A.; Andrés-Lacueva, C.; Lamuela-Raventós, R.M. Improved characterization of tomato polyphenols using liquid chromatography/electrospray ionization linear ion trap quadrupole Orbitrap mass spectrometry and liquid chromatography/electrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 2986–2992. [Google Scholar] [CrossRef] [PubMed]
- Olennikov, D.N.; Chirikova, N.K.; Vasilieva, A.G.; Fedorov, I.A. LC-MS profile, gastrointestinal and gut microbiota stability and antioxidant activity of Rhodiola rosea herb metabolites: A comparative study with subterranean organs. Antioxidants 2020, 9, 526. [Google Scholar] [CrossRef] [PubMed]
- Rockenbach, I.I.; Jungfer, E.; Ritter, C.; Santiago-Schübel, B.; Thiele, B.; Fett, R.; Galensa, R. Characterization of flavan-3-ols in seeds of grape pomace by CE, HPLC-DAD-MSn and LC-ESI-FTICR-MS. Food Res. Int. 2012, 48, 848–855. [Google Scholar] [CrossRef]
- Qu, C.; Fu, F.; Lu, K.; Zhang, K.; Wang, R.; Xu, X.; Wang, M.; Lu, J.; Wan, H.; Zhanglin, T.; et al. Differential accumulation of phenolic compounds and expression of related genes in black- and yellow-seeded Brassica napus. J. Exp. Bot. 2013, 64, 2885–2898. [Google Scholar] [CrossRef] [PubMed]
- Boso, S.; Gago, P.; Santiago, J.-L.; Álvarez-Acero, I.; Martinez Bartolomé, M.-A.; Martínez, M.-C. Polyphenols in the waste water produced during the hydrodistillation of ‘Narcea roses’ cultivated in the Cibea river valley (Northern Spain). Horticulturae 2022, 8, 376. [Google Scholar] [CrossRef]
- Iwashina, T.; Katoh, N. Qualitative and quantitative variation of anthocyanins and flavonols among the different organs of Cercidiphyllum japonicum. Bull. Natl. Mus. Nat. Sci. 2018, 44, 105–114. [Google Scholar]
- Yuan, W.; Wang, J.; An, X.; Dai, M.; Jiang, Z.; Zhang, L.; Yu, S.; Huang, X. UPLC-MS/MS method for the determination of hyperoside and application to pharmacokinetics study in rat after different administration routes. Chromatographia 2021, 84, 249–256. [Google Scholar] [CrossRef]
- Fernandes, P.A.R.; Ferreira, S.S.; Bastos, R.; Ferreira, I.; Cruz, M.T.; Pinto, A.; Coelho, E.; Passos, C.P.; Coimbra, M.A.; Cardoso, S.M.; et al. Apple pomace extract as a sustainable food ingredient. Antioxidants 2019, 8, 189. [Google Scholar] [CrossRef] [PubMed]
- Zymone, K.; Raudone, L.; Žvikas, V.; Jakštas, V.; Janulis, V. Phytoprofiling of Sorbus L. inflorescences: A valuable and promising resource for phenolics. Plants 2022, 11, 3421. [Google Scholar] [CrossRef] [PubMed]
- Owczarek, A.; Kołodziejczyk-Czepas, J.; Marczuk, P.; Siwek, J.; Wąsowicz, K.; Olszewska, M.A. Bioactivity potential of Aesculus hippocastanum L. flower: Phytochemical profile, antiradical capacity and protective effects on human plasma components under oxidative/nitrative stress in vitro. Pharmaceuticals 2021, 14, 1301. [Google Scholar] [CrossRef] [PubMed]
- López-Angulo, G.; Montes-Avila, J.; Díaz-Camacho, S.P.; Vega-Aviña, R.; López-Valenzuela, J.Á.; Delgado-Vargas, F. Comparison of terpene and phenolic profiles of three wild species of Echeveria (Crassulaceae). J. Appl. Bot. Food Qual. 2018, 91, 145–154. [Google Scholar] [CrossRef]
- Clifford, M.N.; Scalbert, A. Ellagitannins–Nature, occurrence and dietary burden. J. Sci. Food Agric. 2000, 80, 1118–1125. [Google Scholar] [CrossRef]
- Camann, J.; Denzel, K.; Schilling, G.; Gross, G.G. Biosynthesis of gallotannins: β-Glucogallin-dependent formation of 1,2,3,4,6-pentagalloylglucose by enzymatic galloylation of 1,2,3,6-tetragalloylglucose. Arch. Biochem. Biophys. 1989, 273, 58–63. [Google Scholar] [CrossRef]
- Ishimaru, K.; Hirose, M.; Takahashi, K.; Koyama, K.; Shimomura, K. Tannin production in root culture of Sanguisorba officinalis. Phytochemistry 1990, 29, 3827–3830. [Google Scholar] [CrossRef]
- Schulenburg, K.; Feller, A.; Hoffmann, T.; Schecker, J.H.; Martens, S.; Schwab, W. Formation of β-glucogallin, the precursor of ellagic acid in strawberry and raspberry. J. Exp. Bot. 2016, 67, 2299–2308. [Google Scholar] [CrossRef]
- Eriksson, T.; Hibbs, M.S.; Yoder, A.D.; Delwiche, C.F.; Donoghue, M.J. The phylogeny of Rosoideae (Rosaceae) based on sequences of the internal transcribed spacers (its) of nuclear ribosomal DNA and the TRNL/F region of chloroplast DNA. Int. J. Plant Sci. 2003, 164, 197–211. [Google Scholar] [CrossRef]
- Wilkes, S.; Glasl, H. Isolation, characterization, and systematic significance of 2-pyrone-4,6-dicarboxylic acid in Rosaceae. Phytochemistry 2001, 58, 441–449. [Google Scholar] [CrossRef]
- Morgan, D.R.; Soltis, D.E.; Robertson, K.R. Systematic and evolutionary implications of rbcL sequence variation in Rosaceae. Am. J. Bot. 1994, 81, 890–903. [Google Scholar] [CrossRef]
- Hameed, A.; Liu, Z.; Wu, H.; Zhong, B.; Ciborowski, M.; Suleria, H.A.R. A comparative and comprehensive characterization of polyphenols of selected fruits from the Rosaceae family. Metabolites 2022, 12, 271. [Google Scholar] [CrossRef]
- Neelam; Khatkar, A.; Sharma, K.K. Phenylpropanoids and its derivatives: Biological activities and its role in food, pharmaceutical and cosmetic industries. Crit. Rev. Food Sci. Nutr. 2020, 60, 2655–2675. [Google Scholar] [CrossRef] [PubMed]
- Vasco, C.; Riihinen, K.; Ruales, J.; Kamal-Eldin, A. Phenolic compounds in Rosaceae fruits from Ecuador. J. Agric. Food Chem. 2009, 57, 1204–1212. [Google Scholar] [CrossRef] [PubMed]
- Patras, M.A.; Jaiswal, R.; McDougall, G.J.; Kuhnert, N. Profiling and quantification of regioisomeric caffeoyl glucoses in berry fruits. J. Agric. Food Chem. 2018, 66, 1096–1104. [Google Scholar] [CrossRef]
- Hussein, S.A.M.; Ayoub, N.A.; Nawwar, M.A.M. Caffeoyl sugar esters and an ellagitannin from Rubus sanctus. Phytochemistry 2003, 63, 905–911. [Google Scholar] [CrossRef]
- Fecka, I.; Bednarska, K.; Włodarczyk, M. Fragaria × ananassa cv. Senga Sengana leaf: An agricultural waste with antiglycation potential and high content of ellagitannins, flavonols, and 2-pyrone-4,6-dicarboxylic acid. Molecules 2022, 27, 5293. [Google Scholar] [CrossRef]
- Jiang, L.; Lu, M.; Rao, T.; Liu, Z.; Wu, X.; An, H. Comparative analysis of fruit metabolome using widely targeted metabolomics reveals nutritional characteristics of different Rosa roxburghii genotypes. Foods 2022, 11, 850. [Google Scholar] [CrossRef]
- Granica, S.; Kluge, H.; Horn, G.; Matkowski, A.; Kiss, A.K. The phytochemical investigation of Agrimonia eupatoria L. and Agrimonia procera Wallr. as valid sources of Agrimoniae herba—The pharmacopoeial plant material. J. Pharm. Biomed. Anal. 2015, 114, 272–279. [Google Scholar] [CrossRef]
- Ortega-Vidal, J.; Ruiz-Martos, L.; Salido, S.; Altarejos, J. Proanthocyanidins in pruning wood extracts of four European plum (Prunus domestica L.) cultivars and their hLDHA inhibitory activity. Chem. Biodivers. 2023, 20, e202200931. [Google Scholar] [CrossRef]
- Sunagawa, T.; Shimizu, T.; Kanda, T.; Tagashira, M.; Sami, M.; Shirasawa, T. Procyanidins from apples (Malus pumila Mill.) extend the lifespan of Caenorhabditis elegans. Planta Med. 2011, 77, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.E.; Cho, J.Y.; Lee, Y.G.; Jeong, H.Y.; Lee, H.J.; Moon, J.H. Isolation of five proanthocyanidins from pear (Pyrus pyrifolia Nakai) fruit peels. Food Sci. Biotechnol. 2017, 26, 1209–1215. [Google Scholar] [CrossRef] [PubMed]
- Uto, T.; Sakamoto, A.; Tung, N.H.; Fujiki, T.; Kishihara, K.; Oiso, S.; Kariyazono, H.; Morinaga, O.; Shoyama, Y. Anti-proliferative activities and apoptosis induction by triterpenes derived from Eriobotrya japonica in human leukemia cell lines. Int. J. Mol. Sci. 2013, 14, 4106–4120. [Google Scholar] [CrossRef] [PubMed]
- Quan, X.X.; Huang, Y.Y.; Chen, L.; Yuan, J.Q. Traditional uses, phytochemical, pharmacology, quality control and modern applications of two important Chinese medicines from Rosa laevigata Michx.: A review. Front. Pharmacol. 2022, 13, 1012265. [Google Scholar] [CrossRef] [PubMed]
- Bate-Smith, E.C. Investigation of the chemistry and taxonomy of sub-tribe Quillajeae of the Rosaceae using comparisons of fresh and herbarium material. Phytochemistry 1965, 4, 535–539. [Google Scholar] [CrossRef]
- Sołtys, A.; Galanty, A.; Podolak, I. Ethnopharmacologically important but underestimated genus Sorbus: A comprehensive review. Phytochem. Rev. 2020, 19, 491–526. [Google Scholar] [CrossRef]
- Baek, Y.S.; Song, N.Y.; Nam, T.G.; Kim, D.-O.; Kang, H.-C.; Kwon, O.-K.; Baek, N.-I. Flavonoids from Fragaria ananassa calyx and their antioxidant capacities. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 787–793. [Google Scholar] [CrossRef]
- Koehler, D.L.; Smith, D.M. Hybridization between Cowania mexicana var. stansburiana and Purshia glandulosa (Rosaceae). Madroño 1981, 28, 13–25. [Google Scholar]
- Jay, M.; De Luca, V.; Ibrahim, R. Meta-methylation of flavonol rings A (8-) and B (3’-) is catalysed by two distinct O-methyltransferases in Lotus corniculatus. Z. Naturforsch. C 1983, 38, 413–417. [Google Scholar] [CrossRef]
- García-Calderón, M.; Pérez-Delgado, C.M.; Palove-Balang, P.; Betti, M.; Márquez, A.J. Flavonoids and isoflavonoids biosynthesis in the model legume Lotus japonicus; connections to nitrogen metabolism and photorespiration. Plants 2020, 9, 774. [Google Scholar] [CrossRef]
- Kharlamova, N.; Sukhova, M.; Chlachula, J. Present climate development in Southern Siberia: A 55-year weather observations record. IOP Conf. Ser. Earth Environ. Sci. 2019, 395, 012027. [Google Scholar] [CrossRef]
- Watanabe, T.; Matsuyama, H.; Kuzhevskaia, I.; Nechepurenko, O.; Chursin, V.; Zemtsov, V. Long-Term trends of extreme climate indexes in the Southern part of Siberia in comparison with those of surrounding regions. Atmosphere 2023, 14, 1131. [Google Scholar] [CrossRef]
- Patanè, G.T.; Putaggio, S.; Tellone, E.; Barreca, D.; Ficarra, S.; Maffei, C.; Calderaro, A.; Laganà, G. Catechins and proanthocyanidins involvement in metabolic syndrome. Int. J. Mol. Sci. 2023, 24, 9228. [Google Scholar] [CrossRef]
- Speisky, H.; Shahidi, F.; Costa de Camargo, A.; Fuentes, J. Revisiting the oxidation of flavonoids: Loss, conservation or enhancement of their antioxidant properties. Antioxidants 2022, 11, 133. [Google Scholar] [CrossRef]
- Singh, P.; Singh, S.M.; D’Souza, L.M.; Wahidullah, S. Phytochemical profiles and antioxidant potential of four Arctic vascular plants from Svalbard. Polar Biol. 2012, 35, 1825–1836. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kashchenko, N.I. Spireasalicin, a new acylated quercetin glycoside from Spiraea salicifolia. Chem. Nat. Comp. 2017, 53, 1038–1044. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kruglova, M.Y. A new quercetin glycoside and other phenolic compounds from the genus Filipendula. Chem. Nat. Comp. 2013, 49, 610–616. [Google Scholar] [CrossRef]
- LabSolutions. Available online: https://www.shimadzu.eu/labsolutions-0 (accessed on 27 February 2024).
- Olennikov, D.N.; Chirikova, N.K.; Kashchenko, N.I.; Nikolaev, V.M.; Kim, S.-W.; Vennos, C. Bioactive phenolics of the genus Artemisia (Asteraceae): HPLC-DAD-ESI-TQ-MS/MS profile of the Siberian species and their inhibitory potential against α-amylase and α-glucosidase. Front. Pharmacol. 2018, 9, 756. [Google Scholar] [CrossRef] [PubMed]
- Olennikov, D.N.; Zilfikarov, I.N.; Penzina, T.A. Use of microcolumn HPLC for analysis of aloenin in Aloe arborescens raw material and related drugs. Pharm. Chem. J. 2013, 47, 494–497. [Google Scholar] [CrossRef]
- Asker, M.M.S.; Shawky, B.T. Structural characterization and antioxidant activity of an extracellular polysaccharide isolated from Brevibacterium otitidis BTS 44. Food Chem. 2010, 123, 315–320. [Google Scholar] [CrossRef]
- Ding, H.; Chou, T.; Liang, C. Antioxidant and antimelanogenic properties of rosmarinic acid methyl ester from Origanum vulgare. Food Chem. 2010, 123, 254–262. [Google Scholar] [CrossRef]
- Yao, Y.; Chen, S.; Li, H. An improved system to evaluate superoxide-scavenging effects of bioflavonoids. ChemistryOpen 2021, 10, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Luqman, S.; Kumar, R. Importance of deoxyribose degradation assay for evaluating hydroxyl radical scavenging activity of Punica extract. Int. J. Food Prop. 2012, 15, 942–948. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| No. | tR, min | UV Pattern a | Molecular Formula | ESI-MS, m/z (Deprotonated ion [M − H]−: Daughter Ions) | Compound [Ref.] | IL b |
|---|---|---|---|---|---|---|
| 1 | 1.48 | A | C15H18O9 | 341: 179 | 6-O-Caffeoyl glucose [21] | 1 |
| 2 | 1.92 | B | C13H16O10 | 331: 169 | Galloyl glucose [22] | 2 |
| 3 | 2.29 | B | C13H16O10 | 331: 169 | 1-O-Galloyl glucose [22] | 1 |
| 4 | 3.02 | A | C15H18O9 | 341: 179 | 1-O-Caffeoyl glucose [21] | 1 |
| 5 | 4.96 | C | C30H26O12 | 577 | Procyanidin B1 [23] | 1 |
| 6 | 5.63 | C | C15H14O6 | 289 | (+)-Catechin [22] | 1 |
| 7 | 6.08 | D | C15H18O8 | 325: 163 | 1-O-p-Coumaroyl glucose [21] | 1 |
| 8 | 6.97 | C | C30H26O12 | 577 | Procyanidin B2 [23] | 1 |
| 9 | 7.38 | A | C16H20O9 | 355: 193 | 1-O-Feruloyl glucose [21] | 1 |
| 10 | 8.08 | A | C24H24O14 | 503: 341, 179 | 1,6-Di-O-caffeoyl glucose [21] | 1 |
| 11 | 8.31 | C | C15H14O6 | 289 | (−)-Epicatechin [22] | 1 |
| 12 | 9.00 | E | C33H40O22 | 787: 625, 463, 301 | Quercetin 3-O-trihexoside [24] | 2 |
| 13 | 9.42 | E | C32H38O21 | 757: 625, 595, 463, 301 | Quercetin 3-O-pentoside-O-dihexoside [25] | 2 |
| 14 | 10.67 | E | C27H30O17 | 625: 463, 301 | Quercetin 3-O-dihexoside [24] | 2 |
| 15 | 11.12 | E | C26H28O16 | 595: 463, 301 | Quercetin 3-O-pentoside-O-hexoside [25] | 2 |
| 16 | 11.87 | F | C22H22O13 | 493: 331 | Corniculatusin O-hexoside [26] | 2 |
| 17 | 12.76 | F | C21H20O12 | 463: 331 | Corniculatusin O-pentoside [26] | 2 |
| 18 | 13.0.8 | E | C21H20O12 | 463: 301 | Quercetin 3-O-galactoside (hyperoside) [27] | 1 |
| 19 | 13.67 | E | C20H18O11 | 433: 301 | Quercetin 3-O-arabinopyranoside (guaijaverin) [28] | 1 |
| 20 | 14.22 | G | C22H22O12 | 477: 315 | Sexangularetin O-hexoside [29] | 2 |
| 21 | 14.79 | G | C21H20O11 | 447: 315 | Sexangularetin O-pentoside [29] | 2 |
| 22 | 15.55 | E | C20H18O11 | 433: 301 | Quercetin 3-O-xyloside (reynoutrin) [28] | 1 |
| 23 | 16.01 | E | C20H18O11 | 433: 301 | Quercetin 3-O-arabinofuranoside (avicularin) [28] | 1 |
| 24 | 16.71 | H | C21H20O11 | 447: 285 | Kaempferol 3-O-galactoside (trifolin) [30] | 1 |
| 25 | 17.11 | H | C20H18O10 | 417: 285 | Kaempferol 3-O-arabinofuranoside (juglanin) [30] | 1 |
| 26 | 17.89 | I | C35H34O19 | 757: 625, 595, 463, 301 | Quercetin O-caffeoyl-O-pentoside-O-hexoside [31] | 2 |
| 27 | 18.09 | J | C31H28O16 | 655: 493, 331 | Corniculatusin O-caffeoyl-O-hexoside [26] | 2 |
| 28 | 18.19 | J | C30H26O15 | 625: 463, 331 | Corniculatusin O-caffeoyl-O-pentoside [26] | 2 |
| 29 | 18.97 | I | C30H26O15 | 625: 463, 301 | Quercetin O-caffeoyl-O-hexoside [31] | 2 |
| 30 | 19.42 | I | C29H24O14 | 595: 433, 301 | Quercetin O-caffeoyl-O-pentoside [31] | 2 |
| 31 | 19.63 | K | C31H28O15 | 639: 477, 315 | Sexangularetin O-caffeoyl-O-hexoside [29] | 2 |
| 32 | 19.94 | K | C30H26O14 | 609: 447, 315 | Sexangularetin O-caffeoyl-O-pentoside [29] | 2 |
| 33 | 20.02 | I | C29H24O14 | 595: 433, 301 | Quercetin O-caffeoyl-O-pentoside [31] | 2 |
| 34 | 20.53 | I | C29H24O14 | 595: 433, 301 | Quercetin O-caffeoyl-O-pentoside [31] | 2 |
| 35 | 20.82 | L | C30H48O5 | 487 | Tormentic acid [19] | 1 |
| 36 | 21.02 | L | C30H48O4 | 471 | Corosolic acid [19] | 1 |
| 37 | 21.12 | L | C30H48O3 | 455 | Ursolic acid [19] | 1 |
| 38 | 21.20 | M | C30H26O14 | 609: 447, 285 | Kaempferol O-caffeoyl-O-hexoside [30] | 2 |
| 39 | 21.53 | L | C30H48O3 | 455 | Ursolic acid isomer [19] | 2 |
| 40 | 21.89 | M | C29H24O13 | 579: 417, 285 | Kaempferol O-caffeoyl-O-pentoside [30] | 2 |
| No. | Compound | Sakha 1000 m (n = 14) | Buryatia 1900 m (n = 14) | Altai 2300 m (n = 12) | |||
|---|---|---|---|---|---|---|---|
| Flowers | Leaves | Flowers | Leaves | Flowers | Leaves | ||
| 1 | 6-O-Caffeoyl glucose | 0.93 ± 0.02 | 0.90 ± 0.02 | 1.03 ± 0.02 | tr. | tr. | tr. |
| 2 | Galloyl glucose | 1.87 ± 0.03 | 5.11 ± 0.11 | 1.69 ± 0.04 | 2.83 ± 0.05 | tr. | tr. |
| 3 | 1-O-Galloyl glucose | 2.14 ± 0.04 | 3.73 ± 0.07 | 1.53 ± 0.03 | 1.65 ± 0.03 | 0.93 ± 0.02 | tr. |
| 4 | 1-O-Caffeoyl glucose | 3.75 ± 0.06 | 0.93 ± 0.02 | 4.83 ± 0.10 | 1.27 ± 0.02 | 1.11 ± 0.02 | 1.24 ± 0.02 |
| 5 | Procyanidin B1 | 55.11 ± 0.99 | 73.84 ± 1.40 | 42.14 ± 0.88 | 40.83 ± 0.86 | 21.15 ± 0.38 | 20.53 ± 0.41 |
| 6 | (+)-Catechin | 14.82 ± 0.27 | 45.77 ± 0.96 | 26.84 ± 0.51 | 39.14 ± 0.82 | 14.22 ± 0.27 | 22.83 ± 0.43 |
| 7 | 1-O-p-Coumaroyl glucose | 2.63 ± 0.04 | 1.73 ± 0.04 | 3.79 ± 0.08 | 0.61 ± 0.01 | 0.27 ± 0.01 | tr. |
| 8 | Procyanidin B2 | 19.25 ± 0.35 | 29.16 ± 0.61 | 10.06 ± 0.19 | 11.29 ± 0.24 | 9.57 ± 0.19 | 8.27 ± 0.17 |
| 9 | 1-O-Feruloyl glucose | 0.83 ± 0.02 | tr. | 0.93 ± 0.02 | tr. | 0.52 ± 0.01 | tr. |
| 10 | 1,6-Di-O-caffeoyl glucose | 0.24 ± 0.00 | tr. | 0.12 ± 0.00 | tr. | 0.14 ± 0.00 | tr. |
| 11 | (−)-Epicatechin | 26.84 ± 0.51 | 52.17 ± 1.04 | 42.03 ± 0.84 | 73.15 ± 1.61 | 22.82 ± 0.46 | 35.11 ± 0.70 |
| 12 | Quercetin 3-O-trihexoside | 1.01 ± 0.02 | tr. | 0.12 ± 0.00 | tr. | 1.09 ± 0.02 | 0.14 ± 0.00 |
| 13 | Quercetin 3-O-pentoside-O-dihexoside | 0.67 ± 0.01 | tr. | 0.09 ± 0.00 | tr. | 1.72 ± 0.04 | 0.21 ± 0.00 |
| 14 | Quercetin 3-O-dihexoside | 1.43 ± 0.03 | tr. | 0.53 ± 0.01 | tr. | 4.35 ± 0.09 | 0.41 ± 0.01 |
| 15 | Quercetin 3-O-pentoside-O-hexoside | 1.24 ± 0.02 | tr. | 0.72 ± 0.02 | tr. | 2.09 ± 0.04 | 0.52 ± 0.01 |
| 16 | Corniculatusin 3-O-galactoside | 7.83 ± 0.15 | 2.73 ± 0.05 | 5.79 ± 0.12 | 0.50 ± 0.01 | 0.11 ± 0.00 | 0.08 ± 0.00 |
| 17 | Corniculatusin 3-O-arabinoside | 6.33 ± 0.11 | 1.29 ± 0.03 | 4.25 ± 0.09 | 0.41 ± 0.01 | 0.23 ± 0.00 | 0.09 ± 0.00 |
| 18 | Quercetin 3-O-galactoside (hyperoside) | 43.27 ± 0.69 | 8.21 ± 0.16 | 58.33 ± 1.22 | 12.63 ± 0.21 | 27.63 ± 0.58 | 2.63 ± 0.06 |
| 19 | Quercetin 3-O-arabinopyranoside (guaijaverin) | 10.52 ± 0.19 | 5.27 ± 0.10 | 14.63 ± 0.28 | 6.24 ± 0.12 | 22.85 ± 0.48 | 11.81 ± 0.26 |
| 20 | Sexangularetin 3-O-galactoside | 1.52 ± 0.03 | 0.40 ± 0.01 | 0.84 ± 0.01 | tr. | 0.28 ± 0.01 | 0.15 ± 0.00 |
| 21 | Sexangularetin 3-O-arabinoside | 2.16 ± 0.04 | 0.52 ± 0.01 | 0.92 ± 0.02 | tr. | 0.37 ± 0.01 | 0.06 ± 0.00 |
| 22 | Quercetin 3-O-xyloside (reynoutrin) | 5.27 ± 0.10 | tr. | 6.83 ± 0.14 | 0.83 ± 0.01 | 11.64 ± 0.23 | 9.83 ± 0.17 |
| 23 | Quercetin 3-O-arabinofuranoside (avicularin) | 9.83 ± 0.17 | 1.53 ± 0.03 | 14.95 ± 0.31 | 3.86 ± 0.08 | 27.82 ± 0.53 | 5.22 ± 0.09 |
| 24 | Kaempferol 3-O-galactoside (trifolin) | 4.03 ± 0.08 | 0.93 ± 0.02 | 5.29 ± 0.09 | 1.95 ± 0.04 | 9.65 ± 0.18 | 2.10 ± 0.04 |
| 25 | Kaempferol 3-O-arabinofuranoside (juglanin) | 0.52 ± 0.01 | tr. | 1.89 ± 0.04 | 0.52 ± 0.01 | 3.76 ± 0.07 | 1.52 ± 0.03 |
| 26 | Quercetin O-caffeoyl-O-pentoside-O-hexoside | 0.41 ± 0.01 | tr. | 1.22 ± 0.02 | 0.21 ± 0.00 | 1.73 ± 0.03 | tr. |
| 27 | Corniculatusin O-caffeoyl-O-hexoside | 2.67 ± 0.05 | 0.63 ± 0.01 | 0.72 ± 0.02 | tr. | tr. | 0.27 ± 0.00 |
| 28 | Corniculatusin O-caffeoyl-O-pentoside | 1.54 ± 0.03 | 1.14 ± 0.03 | 0.70 ± 0.01 | tr. | 0.44 ± 0.01 | 0.19 ± 0.00 |
| 29 | Quercetin O-caffeoyl-O-hexoside | 0.24 ± 0.00 | tr. | 0.63 ± 0.01 | 0.08 ± 0.00 | 1.14 ± 0.02 | tr. |
| 30 | Quercetin O-caffeoyl-O-pentoside | 0.12 ± 0.00 | tr. | 0.69 ± 0.01 | 0.05 ± 0.00 | 1.52 ± 0.03 | tr. |
| 31 | Sexangularetin O-caffeoyl-O-hexoside | 0.53 ± 0.01 | 0.04 ± 0.00 | 0.08 ± 0.00 | tr. | 0.02 ± 0.00 | tr. |
| 32 | Sexangularetin O-caffeoyl-O-pentoside | 2.67 ± 0.06 | 0.22 ± 0.00 | 1.10 ± 0.02 | 0.09 ± 0.00 | tr. | tr. |
| 33 | Quercetin O-caffeoyl-O-pentoside | tr. | tr. | 0.54 ± 0.01 | tr. | 0.97 ± 0.02 | tr. |
| 34 | Quercetin O-caffeoyl-O-pentoside | tr. | tr. | 0.63 ± 0.01 | tr. | 1.49 ± 0.03 | tr. |
| 35 | Tormentic acid | 6.85 ± 0.14 | 7.39 ± 0.14 | 2.94 ± 0.06 | 3.85 ± 0.08 | 1.41 ± 0.03 | 2.53 ± 0.05 |
| 36 | Corosolic acid | 2.74 ± 0.06 | 3.67 ± 0.07 | 0.53 ± 0.01 | 1.02 ± 0.02 | 0.92 ± 0.02 | 1.14 ± 0.02 |
| 37 | Ursolic acid | 2.22 ± 0.04 | 1.29 ± 0.03 | 0.60 ± 0.01 | 0.42 ± 0.01 | 0.57 ± 0.01 | 1.69 ± 0.03 |
| 38 | Kaempferol O-caffeoyl-O-hexoside | tr. | tr. | 0.63 ± 0.01 | tr. | 0.90 ± 0.02 | 0.52 ± 0.01 |
| 39 | Ursolic acid isomer | 1.14 ± 0.02 | 0.93 ± 0.02 | 0.83 ± 0.02 | 0.92 ± 0.02 | 1.14 ± 0.02 | 0.95 ± 0.02 |
| 40 | Kaempferol O-caffeoyl-O-pentoside | tr. | tr. | 0.69 ± 0.01 | tr. | 2.11 ± 0.04 | 0.92 ± 0.02 |
| Total cinnamoyl glucoses | 8.38 | 3.56 | 10.70 | 1.88 | 2.04 | 1.24 | |
| Total galloyl glucoses | 4.01 | 8.84 | 3.22 | 4.48 | 0.93 | tr. | |
| Total procyanidins | 74.36 | 103.00 | 52.20 | 52.12 | 30.72 | 28.80 | |
| Total catechins | 41.66 | 97.94 | 68.87 | 112.29 | 37.04 | 57.94 | |
| Total flavonoids, including: | 103.81 | 22.91 | 122.81 | 27.37 | 123.91 | 36.67 | |
| Kaempferol glucosides | 4.55 | 0.93 | 8.50 | 2.47 | 16.42 | 5.06 | |
| Quercetin glucosides | 74.01 | 15.01 | 99.91 | 23.90 | 106.04 | 30.77 | |
| Sexangularetin glucosides | 6.88 | 1.18 | 2.94 | 0.09 | 0.67 | 0.21 | |
| Corniculatusin glucosides | 18.37 | 5.79 | 11.46 | 0.91 | 0.78 | 0.63 | |
| Total phenolics | 232.22 | 236.25 | 257.80 | 198.14 | 194.64 | 124.65 | |
| Total triterpenes | 12.95 | 13.28 | 4.90 | 6.21 | 4.04 | 6.31 | |
| Extracts | DPPH• a | ABTS•+ a | O2•– a | •OH a |
|---|---|---|---|---|
| SFE | 9.23 ± 0.18 | 5.04 ± 0.10 | 26.13 ± 0.52 | 14.92 ± 0.29 |
| SLE | 11.08 ± 0.22 | 7.12 ± 0.14 | 37.16 ± 0.74 | 8.08 ± 0.16 |
| BFE | 8.67 ± 0.17 | 3.99 ± 0.08 | 21.59 ± 0.43 | 12.78 ± 0.25 |
| BLE | 11.90 ± 0.23 | 4.36 ± 0.09 | 41.14 ± 0.82 | 19.36 ± 0.39 |
| AFE | 15.01 ± 0.30 | 8.07 ± 0.16 | 58.22 ± 1.16 | 36.05 ± 0.72 |
| ALE | 18.33 ± 0.36 | 10.85 ± 0.21 | 73.01 ± 1.46 | 49.32 ± 0.99 |
| Trolox b | 10.17 ± 0.20 | 4.67 ± 0.09 | 109.28 ± 2.18 | 16.37 ± 0.33 |
| Compound | Regression Equation a | r2 | Syx | LOD/LOQ (µg/mL) | Linear Range (µg/mL) | RSD% (Intra-Day) | RSD% (Inter-Day) | Recovery of Spiked Sample REC% | |
|---|---|---|---|---|---|---|---|---|---|
| a | b × 106 | ||||||||
| 6-O-Caffeoyl glucose | 1.3387 | −0.0284 | 0.9981 | 9.50 × 10−2 | 0.23/0.71 | 1.0–100.0 | 1.45 | 1.92 | 101.27 |
| 1-O-Galloyl glucose | 1.3586 | −0.0663 | 0.9987 | 9.69 × 10−2 | 0.24/0.71 | 1.0–100.0 | 0.97 | 1.16 | 98.34 |
| 1-O-Caffeoyl glucose | 1.5824 | −0.1078 | 0.9965 | 16.25 × 10−2 | 0.34/1.03 | 1.0–100.0 | 0.89 | 1.29 | 100.64 |
| Procyanidin B1 | 1.3722 | −0.0829 | 0.9973 | 9.93 × 10−2 | 0.24/0.72 | 1.0–100.0 | 1.09 | 1.36 | 99.12 |
| (+)-Catechin | 0.9562 | −0.0521 | 0.9971 | 7.79 × 10−2 | 0.27/0.82 | 1.0–100.0 | 1.23 | 1.54 | 100.07 |
| 1-O-p-Coumaroyl glucose | 1.4238 | −0.0891 | 0.9901 | 7.33 × 10−2 | 0.17/0.52 | 1.0–100.0 | 1.38 | 1.88 | 101.14 |
| Procyanidin B2 | 1.3620 | −0.0820 | 0.9961 | 9.91 × 10−2 | 0.21/0.72 | 1.0–100.0 | 1.47 | 1.64 | 100.78 |
| 1-O-Feruloyl glucose | 1.5152 | −0.0523 | 0.9979 | 12.67 × 10−2 | 0.28/0.84 | 1.0–100.0 | 1.33 | 1.61 | 100.11 |
| 1,6-Di-O-caffeoyl glucose | 1.7552 | −0.0569 | 0.9982 | 8.89 × 10−2 | 0.18/0.51 | 1.0–100.0 | 1.40 | 1.98 | 100.39 |
| (−)-Epicatechin | 1.0828 | −0.0456 | 0.9973 | 6.85 × 10−2 | 0.21/0.63 | 1.0–100.0 | 1.12 | 1.42 | 99.23 |
| Hyperoside | 1.4689 | −0.3641 | 0.9990 | 5.69 × 10−2 | 0.12/0.38 | 1.0–100.0 | 1.22 | 1.43 | 101.22 |
| Guaijaverin | 1.3436 | −0.4406 | 0.9981 | 17.58 × 10−2 | 0.43/1.31 | 1.0–100.0 | 0.99 | 1.60 | 102.55 |
| Reynoutrin | 1.5364 | −0.3614 | 0.9927 | 10.07 × 10−2 | 0.22/0.66 | 1.0–100.0 | 1.34 | 1.78 | 102.03 |
| Avicularin | 1.4041 | −0.3270 | 0.9992 | 14.02 × 10−2 | 0.33/1.00 | 1.0–100.0 | 1.31 | 1.57 | 99.83 |
| Trifolin | 2.0859 | −0.9171 | 0.9980 | 6.18 × 10−2 | 0.03/0.09 | 1.0–100.0 | 1.23 | 1.83 | 101.23 |
| Juglanin | 2.2126 | −0.5160 | 0.9987 | 8.11 × 10−2 | 0.12/0.37 | 1.0–100.0 | 1.08 | 1.60 | 98.33 |
| Tormentic acid | 1.5330 | −0.0863 | 0.9985 | 4.15 × 10−2 | 0.09/0.27 | 1.0–100.0 | 1.39 | 1.78 | 100.09 |
| Corosolic acid | 2.3312 | −0.4563 | 0.9803 | 14.92 × 10−2 | 0.21/0.64 | 1.0–100.0 | 1.24 | 1.85 | 101.40 |
| Ursolic acid | 1.2820 | −0.9634 | 0.9697 | 11.64 × 10−2 | 0.30/0.91 | 1.0–100.0 | 1.41 | 1.91 | 99.54 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kashchenko, N.I.; Olennikov, D.N.; Chirikova, N.K. Chemodiversity of Arctic Plant Dryas oxyodonta: LC-MS Profile and Antioxidant Activity. Plants 2024, 13, 868. https://doi.org/10.3390/plants13060868
Kashchenko NI, Olennikov DN, Chirikova NK. Chemodiversity of Arctic Plant Dryas oxyodonta: LC-MS Profile and Antioxidant Activity. Plants. 2024; 13(6):868. https://doi.org/10.3390/plants13060868
Chicago/Turabian StyleKashchenko, Nina I., Daniil N. Olennikov, and Nadezhda K. Chirikova. 2024. "Chemodiversity of Arctic Plant Dryas oxyodonta: LC-MS Profile and Antioxidant Activity" Plants 13, no. 6: 868. https://doi.org/10.3390/plants13060868
APA StyleKashchenko, N. I., Olennikov, D. N., & Chirikova, N. K. (2024). Chemodiversity of Arctic Plant Dryas oxyodonta: LC-MS Profile and Antioxidant Activity. Plants, 13(6), 868. https://doi.org/10.3390/plants13060868