
Genome-Wide Identification of PYL/RCAR ABA Receptors and Functional Analysis of LbPYL10 in Heat Tolerance in Goji (Lycium barbarum)

Abstract
:1. Introduction
2. Results
2.1. Heat Stress–Induced ABA Accumulation in L. barbarum
2.2. Characterization of the LbPYL Gene Family
2.3. Phylogeny Analysis
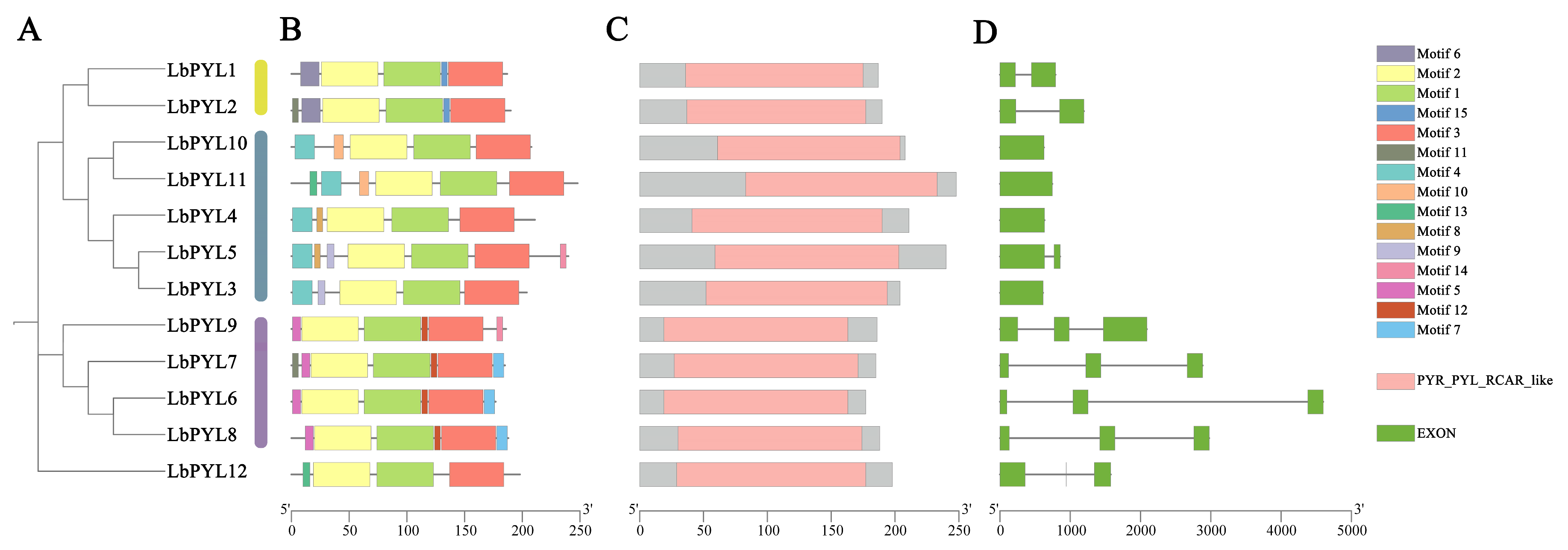
2.4. Gene Structure, Conserved Motifs, and Domain Analysis
2.5. Analysis of Cis–Elements in the LbPYL Promoters
2.6. Tissue–Specific Expression
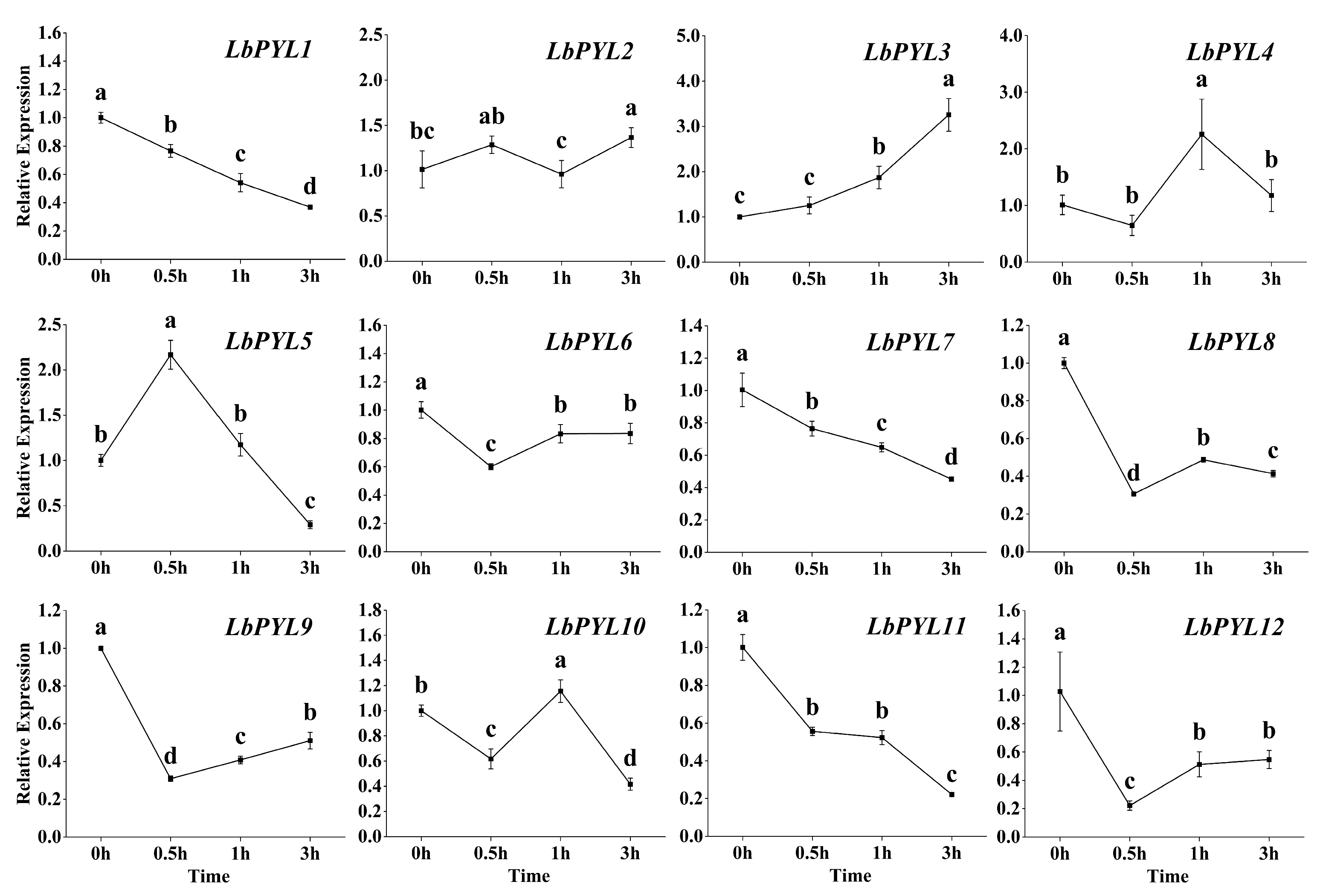
2.7. Transcriptional Response to Exogenous ABA
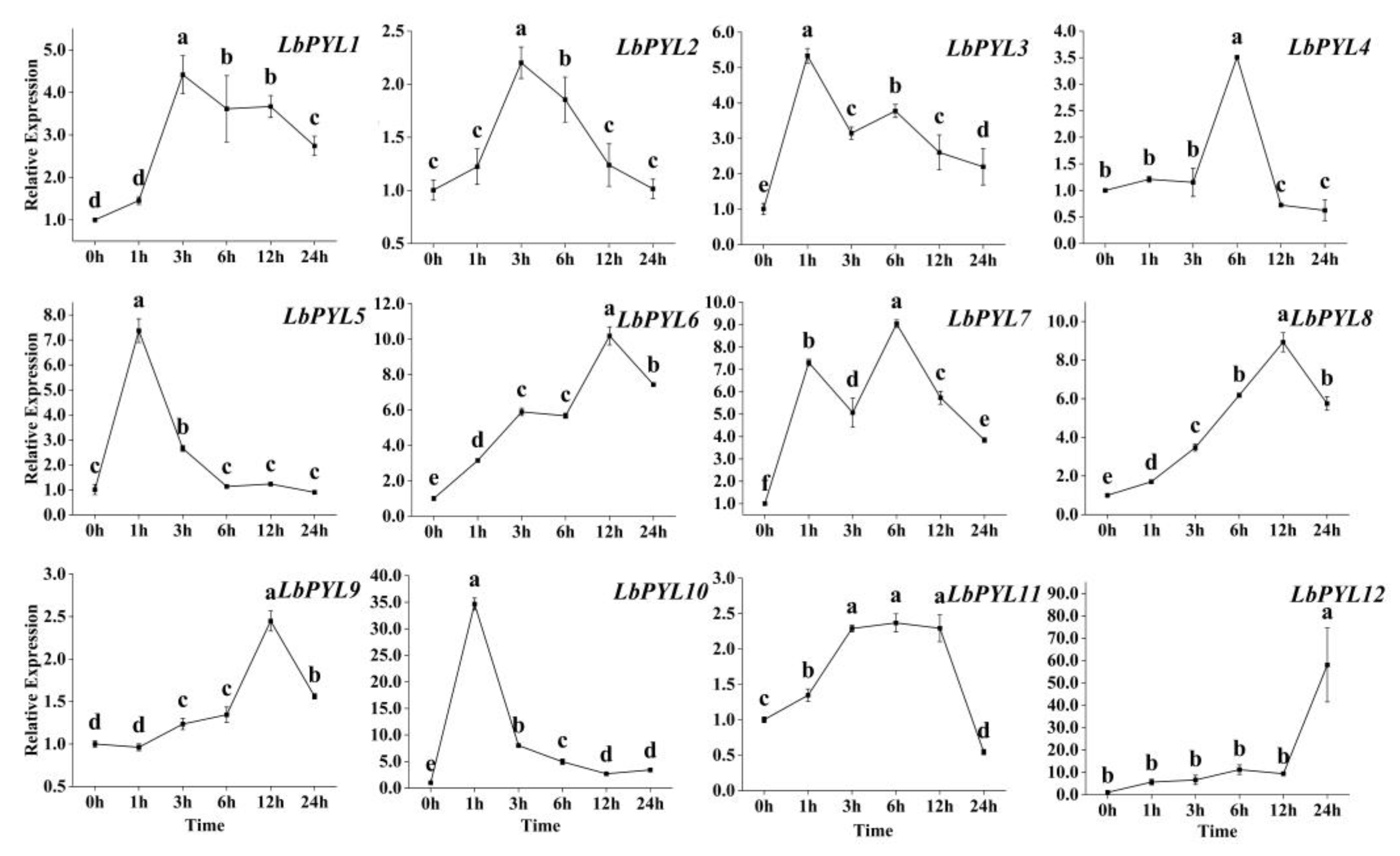
2.8. Transcriptional Response to Heat Stress
2.9. Functional Validation of LbPYL10 in Heat Stress Tolerance
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Heat Stress Assay
4.3. ABA Treatment
4.4. ABA Content Determination
4.5. Virus–Induced LbPYL10 Gene Silencing
4.6. Identification and Chromosomal Location of LbPYL Genes
4.7. Phylogenetic Tree Analysis of LbPYLs in L. barbarum
4.8. Analysis of Gene Exon–Intron Structures and Protein Conserved Motifs
4.9. Analysis of Cis–Elements in the LbPYL Promoters
4.10. Quantitative Real–Time PCR (qRT–PCR) Assay
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yoshida, T.; Christmann, A.; Yamaguchi-Shinozaki, K.; Grill, E.; Fernie, A.R. Revisiting the basal role of ABA-Roles outside of stress. Trends Plant Sci. 2019, 24, 625–635. [Google Scholar] [CrossRef]
- Sun, Y.; Pri-Tal, O.; Michaeli, D.; Mosquna, A. Evolution of abscisic acid signaling module and its perception. Front. Plant Sci. 2020, 11, 934. [Google Scholar] [CrossRef]
- Ma, Y.; Szostkiewicz, I.; Korte, A.; Moes, D.; Yang, Y.; Christmann, A.; Grill, E. Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science 2009, 324, 1064–1068. [Google Scholar] [CrossRef]
- Park, S.; Fung, P.; Nishimura, N.; Jensen, D.R.; Fujii, H.; Zhao, Y.; Lumba, S.; Santiago, J.; Rodrigues, A.; Chow, T.F.; et al. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 2009, 324, 1068–1071. [Google Scholar] [CrossRef]
- Fan, W.; Zhao, M.; Li, S.; Bai, X.; Li, J.; Meng, H.; Mu, Z. Contrasting transcriptional responses of PYR1/PYL/RCAR ABA receptors to ABA or dehydration stress between maize seedling leaves and roots. BMC Plant Biol. 2016, 16, 99–112. [Google Scholar] [CrossRef]
- Bai, G.; Xie, H.; Yao, H.; Li, F.; Chen, X.; Zhang, Y.; Xiao, B.; Yang, J.; Li, Y.; Yang, D.H. Genome-wide identification and characterization of ABA receptor PYL/RCAR gene family reveals evolution and roles in drought stress in Nicotiana Tabacum. BMC Genom. 2019, 20, 575. [Google Scholar] [CrossRef]
- Nian, L.; Zhang, X.; Yi, X.; Liu, X.; Ain, N.U.; Yang, Y.; Li, X.; Haider, F.U.; Zhu, X. Genome-wide identification of ABA receptor PYL/RCAR gene family and their response to cold stress in Medicago sativa L. Physiol. Mol. Biol. Plants 2021, 27, 1979–1995. [Google Scholar] [CrossRef]
- Fujii, H.; Chinnusamy, V.; Rodrigues, A.; Rubio, S.; Antoni, R.; Park, S.Y.; Cutler, S.R.; Sheen, J.; Rodriguez, P.L.; Zhu, J.K. In vitro reconstitution of an abscisic acid signalling pathway. Nature 2009, 462, 660–664. [Google Scholar] [CrossRef]
- Dittrich, M.; Mueller, H.M.; Bauer, H.; Peirats-Llobet, M.; Rodriguez, P.L.; Geilfus, C.M.; Carpentier, S.C.; Al Rasheid, K.A.S.; Kollist, H.; Merilo, E.; et al. The role of Arabidopsis ABA receptors from the PYR/PYL/RCAR family in stomatal acclimation and closure signal integration. Nat. Plants 2019, 5, 1002–1011. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, J.; Poree, F.; Schaeufele, R.; Helmke, H.; Frackenpohl, J.; Lehr, S.; von Koskull-Döring, P.; Christmann, A.; Schnyder, H.; et al. Abscisic acid receptors and coreceptors modulate plant water use efficiency and water productivity. Plant Physiol. 2019, 180, 1066–1080. [Google Scholar] [CrossRef]
- Mega, R.; Abe, F.; Kim, J.S.; Tsuboi, Y.; Tanaka, K.; Kobayashi, H.; Sakata, Y.; Hanada, K.; Tsujimoto, H.; Kikuchi, J.; et al. Tuning water-use efficiency and drought tolerance in wheat using abscisic acid receptors. Nat. Plants 2019, 5, 153–159. [Google Scholar] [CrossRef]
- Mao, H.; Jian, C.; Cheng, X.; Chen, B.; Mei, F.; Li, F.; Zhang, Y.; Li, S.; Du, L.; Li, T.; et al. The wheat ABA receptor gene TaPYL1-1B contributes to drought tolerance and grain yield by increasing water-use efficiency. Plant Biotechnol. J. 2022, 20, 846–861. [Google Scholar] [CrossRef]
- Hewage, K.A.H.; Yang, J.F.; Wang, D.; Hao, G.F.; Yang, G.F.; Zhu, J.K. Chemical manipulation of abscisic acid signaling: A new approach to abiotic and biotic stress management in agriculture. Adv. Sci. 2020, 7, 2001265. [Google Scholar] [CrossRef]
- Lozano-Juste, J.; Infantes, L.; Garcia-Maquilon, I.; Ruiz-Partida, R.; Merilo, E.; Benavente, J.L.; Velazquez-Campoy, A.; Coego, A.; Bono, M.; Forment, J.; et al. Structure-guided engineering of a receptor-agonist pair for inducible activation of the ABA adaptive response to drought. Sci. Adv. 2023, 9, e9948. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Peterson, F.C.; Mosquna, A.; Yao, J.; Volkman, B.F.; Cutler, S.R. Agrochemical control of plant water use using engineered abscisic acid receptors. Nature 2015, 520, 545–548. [Google Scholar] [CrossRef] [PubMed]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 20, 17–62. [Google Scholar] [CrossRef] [PubMed]
- Guihur, A.; Rebeaud, M.E.; Goloubinoff, P. How do plants feel the heat and survive? Trends Biochem. Sci. 2022, 47, 824–838. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Euring, D.; Cha, J.Y.; Lin, Z.; Lu, M.; Huang, L.J.; Kim, W.Y. Plant hormone-mediated regulation of heat tolerance in response to global climate change. Front. Plant Sci. 2021, 11, 627969. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.R.; Feng, B.H.; Chen, T.T.; Tao, L.X.; Fu, G.F. Role of abscisic acid in thermal acclimation of plants. J. Plant Biol. 2018, 61, 255–264. [Google Scholar] [CrossRef]
- Huang, Y.C.; Niu, C.Y.; Yang, C.R.; Jinn, T.L. The heat stress factor HSFA6b connects ABA signaling and ABA-mediated heat responses. Plant Physiol. 2016, 172, 1182–1199. [Google Scholar] [CrossRef]
- Li, H.; Liu, S.S.; Yi, C.Y.; Wang, F.; Zhou, J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Hydrogen peroxide mediates abscisic acid-induced HSP70 accumulation and heat tolerance in grafted cucumber plants. Plant Cell Environ. 2014, 37, 2768–2780. [Google Scholar] [CrossRef]
- Wang, X.; Zhuang, L.; Shi, Y.; Huang, B. Up-regulation of HSFA2c and HSPs by ABA contributing to improved heat tolerance in tall fescue and Arabidopsis. Int. J. Mol. Sci. 2017, 18, 1981. [Google Scholar] [CrossRef]
- Hu, X.; Carver, B.F.; Powers, C.; Yan, L.; Zhu, L.; Chen, C. Effectiveness of Genomic Selection by Response to Selection for Winter Wheat Variety Improvement. Plant Genome 2019, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Rezaul, I.M.; Baohua, F.; Tingting, C.; Weimeng, F.; Caixia, Z.; Longxing, T.; Guanfu, F. Abscisic acid prevents pollen abortion under high-temperature stress by mediating sugar metabolism in rice spikelets. Physiol. Plant. 2019, 165, 644–663. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, N.; Sehar, Z.; Fatma, M.; Umar, S.; Sofo, A.; Khan, N.A. Nitric oxide and abscisic acid mediate heats tress tolerance through regulation of osmolytes and antioxidants to protect photosynthesis and growth in wheat plants. Antioxidants 2022, 11, 372. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhong, X.; Liao, J.; Ji, P.; Yang, J.; Cao, Z.; Duan, X.; Xiong, J.; Wang, Y.; Xu, C.; et al. Exogenous abscisic acid improves grain filling capacity under heat stress by enhancing antioxidative defense capability in rice. BMC Plant Biol. 2023, 23, 619. [Google Scholar] [CrossRef]
- Yao, R.; Heinrich, M.; Weckerle, C.S. The genus Lycium as food and medicine: A botanical, ethnobotanical and historical review. J. Ethnopharmacol. 2018, 212, 50–66. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.L.; Li, Y.L.; Fan, Y.F.; Li, Z.; Yoshida, K.; Wang, J.Y.; Ma, X.K.; Wang, N.; Mitsuda, N.; Kotake, T.; et al. Wolfberry genomes and the evolution of Lycium (Solanaceae). Commun. Biol. 2021, 4, 671. [Google Scholar] [CrossRef]
- Gong, H.; Rehman, F.; Ma, Y.; Zeng, S.; Yang, T.; Huang, J.; Li, Z.; Wu, D.; Wang, Y. Germplasm resources and strategy for genetic breeding of Lycium species: A Review. Front. Plant Sci. 2022, 13, 802936. [Google Scholar] [CrossRef]
- He, W.; Liu, M.; Qin, X.; Liang, A.; Chen, Y.; Yin, Y.; Qin, K.; Mu, Z. Genome-Wide Identification and Expression Analysis of the Aquaporin Gene Family in Lycium barbarum during Fruit Ripening and Seedling Response to Heat Stress. Curr. Issues Mol. Biol. 2022, 44, 5933–5948. [Google Scholar] [CrossRef] [PubMed]
- González-Guzmán, M.; Rodríguez, L.; Lorenzo-Orts, L.; Pons, C.; Sarrión-Perdigones, A.; Fernández, M.A.; Peirats-Llobet, M.; Forment, J.; Moreno-Alvero, M.; Cutler, S.R.; et al. Tomato PYR/PYL/RCAR abscisic acid receptors show high expression in root, differential sensitivity to the abscisic acid agonist quinabactin, and the capability to enhance plant drought resistance. J. Exp. Bot. 2014, 65, 4451–4464. [Google Scholar] [CrossRef] [PubMed]
- Waadt, R.; Seller, C.A.; Hsu, P.K.; Takahashi, Y.; Munemasa, S.; Schroeder, J.I. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 680–694. [Google Scholar] [CrossRef] [PubMed]
- Neill, E.M.; Byrd, M.C.R.; Billman, T.; Brandizzi, F.; Stapleton, A.E. Plant growth regulators interact with elevated temperature to alter heat stress signaling via the unfolded protein response in maize. Sci. Rep. 2019, 9, 10392. [Google Scholar] [CrossRef]
- Sharma, L.; Priya, M.; Kaushal, N.; Bhandhari, K.; Chaudhary, S.; Dhankher, O.P.; Prasad, P.V.V.; Siddique, K.H.M.; Nayyar, H. Plant growth-regulating molecules as thermoprotectants: Functional relevance and prospects for improving heat tolerance in food crops. J. Exp. Bot. 2020, 71, 569–594. [Google Scholar] [CrossRef]
- Li, Q.; Shen, C.; Zhang, Y.; Zhou, Y.; Niu, M.; Wang, H.L.; Lian, C.; Tian, Q.; Mao, W.; Wang, X.; et al. PePYL4 enhances drought tolerance by modulating water-use efficiency and ROS scavenging in Populus. Tree Physiol. 2023, 43, 102–117. [Google Scholar] [CrossRef]
- Barros, J.A.; Cavalcanti, J.H.F.; Medeiros, D.; Nunes-Nesi, A.; Avin-Wittenberg, T.; Fernie, A.R.; Araújo, W.L. Autophagy deficiency compromises alternative pathways of respiration following energy deprivation in Arabidopsis thaliana. Plant Physiol. 2017, 175, 62–76. [Google Scholar] [CrossRef]
- Wada, S.; Ishida, H.; Izumi, M.; Yoshimoto, K.; Ohsumi, Y.; Mae, T.; Makino, A. Autophagy plays a role in chloroplast degradation during senescence in individually darkened leaves. Plant Physiol. 2008, 149, 885–893. [Google Scholar] [CrossRef]
- Kóta, Z.; Horváth, L.I.; Droppa, M.; Horváth, G.; Farkas, T.; Páli, T. Protein assembly and heat stability in developing thylakoid membranes during greening. Proc. Natl. Acad. Sci. USA 2002, 99, 12149–12154. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhu, L.; Wang, Q.; Hou, X. Autophagy-Related 2 Regulates Chlorophyll Degradation under Abiotic Stress Conditions in Arabidopsis. Int. J. Mol. Sci. 2020, 21, 4515. [Google Scholar] [CrossRef] [PubMed]
- Pruzinská, A.; Tanner, G.; Anders, I.; Roca, M.; Hörtensteiner, S. Chlorophyll breakdown: Pheophorbide a oxygenase is a Rieske-type iron-sulfur protein, encoded by the accelerated cell death 1 gene. Proc. Natl. Acad. Sci. USA 2003, 100, 15259–15264. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Kim, D.; Kim, Y.S.; Hörtensteiner, S.; Paek, N.C. Arabidopsis STAYGREEN-LIKE (SGRL) promotes abiotic stress-induced leaf yellowing during vegetative growth. FEBS Lett. 2014, 588, 3830–3837. [Google Scholar] [CrossRef]
- Chen, P.; Sun, Y.F.; Kai, W.B.; Liang, B.; Zhang, Y.S.; Zhai, X.W.; Jiang, L.; Du, Y.W.; Leng, P. Interactions of ABA signaling core components (SlPYLs, SlPP2Cs, and SlSnRK2s) in tomato (Solanum lycopersicon). J. Plant Physiol. 2016, 205, 67–74. [Google Scholar] [CrossRef]
- Sun, S.; Fan, W.; Mu, Z. The spatio-temporal specificity of PYR1/PYL/RCAR ABA receptors in response to developmental and environmental cues. Plant Signal. Behav. 2017, 12, e1214793. [Google Scholar] [CrossRef] [PubMed]
- Tischer, S.V.; Wunschel, C.; Papacek, M.; Kleigrewe, K.; Hofmann, T.; Christmann, A.; Grill, E. Combinatorial interaction network of abscisic acid receptors and coreceptors from Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2017, 114, 10280–10285. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.G.; Fu, F.L.; Yu, H.Q.; Hu, T.; Zhang, Y.Y.; Tao, Y.; Zhu, J.K.; Zhao, Y.; Li, W.C. Interaction network of core ABA signaling components in maize. Plant Mol. Biol. 2018, 96, 245–263. [Google Scholar] [CrossRef] [PubMed]
- Lee, U.; Wie, C.; Fernandez, B.O.; Feelisch, M.; Vierling, E. Modulation of nitrosative stress by S-nitrosoglutathione reductase is critical for thermotolerance and plant growth in Arabidopsis. Plant Cell 2008, 20, 786–802. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.Q.; Li, X.; Niu, L.; Jameson, P.; Zhou, W.B. Transcription-associated metabolomic adjustments in maize occur during combined drought and cold stress. Plant Physiol. 2021, 186, 677–695. [Google Scholar] [CrossRef]
- Liu, Y.L.; Sun, W.; Zeng, S.H.; Huang, W.J.; Liu, D.; Hu, W.M.; Shen, X.F.; Wang, Y. Virus-induced gene silencing in two novel functional plants, Lycium barbarum L. and Lycium ruthenicum Murr. Sci. Hortic. 2014, 170, 267–274. [Google Scholar] [CrossRef]
- Chen, C.J.; Wu, Y.; Li, J.W.; Wang, X.; Zeng, Z.H.; Xu, J.; Liu, Y.L.; Feng, J.T.; Chen, H.; He, Y.H.; et al. TBtools-II: A “one for all, all for one” bioinformatics platform for biological big-data mining. Mol. Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.M.; Chen, Y.R.; Cai, G.J.; Cai, R.L.; Hu, Z.; Wang, H. Tree Visualization by One Table (tvBOT): A web application for visualizing, modifying and annotating phylogenetic trees. Nucleic Acids Res. 2023, 51, 587–592. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Version | Gene Length (bp) | CDS Length (bp) | Deduced Protein | ||
|---|---|---|---|---|---|---|
| Size (aa) | MW (kDa) | PI | ||||
| LbPYL1 | XM_060355288.1 | 1356 | 564 | 187 | 21.045 | 6.16 |
| LbPYL2 | XM_060315403.1 | 1640 | 573 | 190 | 21.305 | 5.74 |
| LbPYL3 | XM_060316745.1 | 970 | 615 | 204 | 22.646 | 6.23 |
| LbPYL4 | XM_060357010.1 | 992 | 636 | 211 | 24.401 | 6.29 |
| LbPYL5 | XM_060322965.1 | 1315 | 723 | 240 | 26.651 | 5.52 |
| LbPYL6 | XM_060317057.1 | 5513 | 534 | 177 | 20.229 | 6.38 |
| LbPYL7 | XM_060347829.1 | 3489 | 558 | 185 | 21.078 | 5.75 |
| LbPYL8 | XM_060350890.1 | 3670 | 567 | 188 | 21.459 | 6.30 |
| LbPYL9 | XM_060362463.1 | 2092 | 561 | 186 | 20.762 | 5.96 |
| LbPYL10 | XM_060346502.1 | 1178 | 627 | 208 | 22.874 | 8.18 |
| LbPYL11 | XM_060320401.1 | 1103 | 747 | 248 | 27.636 | 8.65 |
| LbPYL12 | XM_060337877.1 | 2052 | 597 | 198 | 22.276 | 5.34 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Liu, J.; Chen, Y.; Liang, A.; He, W.; Qin, X.; Qin, K.; Mu, Z. Genome-Wide Identification of PYL/RCAR ABA Receptors and Functional Analysis of LbPYL10 in Heat Tolerance in Goji (Lycium barbarum). Plants 2024, 13, 887. https://doi.org/10.3390/plants13060887
Li Z, Liu J, Chen Y, Liang A, He W, Qin X, Qin K, Mu Z. Genome-Wide Identification of PYL/RCAR ABA Receptors and Functional Analysis of LbPYL10 in Heat Tolerance in Goji (Lycium barbarum). Plants. 2024; 13(6):887. https://doi.org/10.3390/plants13060887
Chicago/Turabian StyleLi, Zeyu, Jiyao Liu, Yan Chen, Aihua Liang, Wei He, Xiaoya Qin, Ken Qin, and Zixin Mu. 2024. "Genome-Wide Identification of PYL/RCAR ABA Receptors and Functional Analysis of LbPYL10 in Heat Tolerance in Goji (Lycium barbarum)" Plants 13, no. 6: 887. https://doi.org/10.3390/plants13060887
APA StyleLi, Z., Liu, J., Chen, Y., Liang, A., He, W., Qin, X., Qin, K., & Mu, Z. (2024). Genome-Wide Identification of PYL/RCAR ABA Receptors and Functional Analysis of LbPYL10 in Heat Tolerance in Goji (Lycium barbarum). Plants, 13(6), 887. https://doi.org/10.3390/plants13060887