
Distribution Pattern of Suitable Areas and Corridor Identification of Endangered Ephedra Species in China


 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Contemporary Distribution Patterns of Endangered Ephedra Species
2.2. Future Distribution Patterns of Endangered Ephedra Species
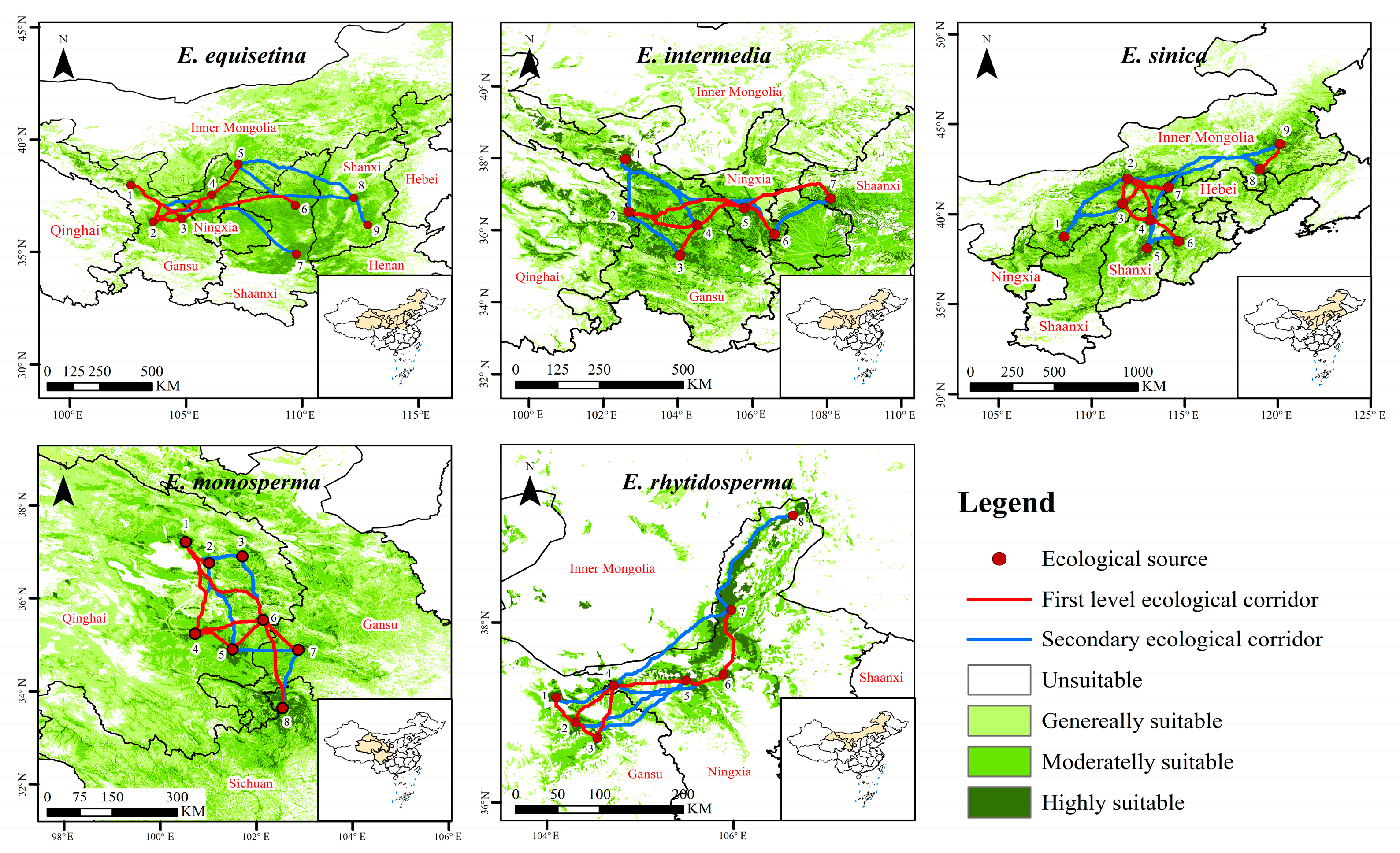
2.3. Corridor Identification of Endangered Ephedra Species
3. Discussion
4. Materials and Methods
4.1. Data Sources and Data Processing
4.2. Suitable Habitat Evaluation
4.3. MCR Modeling for Corridor Identification
5. Conclusions
- (1)
- Under the current climate scenario, four species, namely E. equisetina, E. intermedia, E. sinica, and E. monosperma, inhabit over 16% of their suitable area in China. Conversely, E. rhytidosperma occupies a comparatively smaller proportion of its suitable areas in China at only 0.05%. Altitude was the most critical factor limiting the growth of E. intermedia and E. monosperma, while salinity played a pivotal role in constraining the growth of E. equisetina and E. rhytidosperma. In contrast, the distribution of E. sinica exhibited a stronger dependence on precipitation factors.
- (2)
- Under the future climate scenario, the expansion of the suitable area of E. equisetina, E. intermedia, and E. sinica was maximized under the SSP585 scenario (2090s). The suitable area of E. monosperma shrank, with the greatest degree of shrinkage in the SSP585 scenario (2050s), by 12.42%. In contrast, E. rhytidosperma loses its suitable area under future climate scenarios.
- (3)
- By employing the MCR model and gravity model, we successfully identified 71 crucial ecological corridors for five Ephedra species, all of which are strategically located away from anthropogenic surfaces. These corridors play a pivotal role in ensuring the long-term survival of Ephedra species. Identifying these ecological corridors provides valuable insights into the conservation and management strategies for Ephedra species.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Srivastava, V.; Griess, V.C.; Keena, M.A. Assessing the Potential Distribution of Asian Gypsy Moth in Canada: A Comparison of Two Methodological Approaches. Sci. Rep. 2020, 10, 22. [Google Scholar] [CrossRef] [PubMed]
- Box, E.O.; Fujiwara, K. A Comparative Look at Bioclimatic Zonation, Vegetation Types, Tree Taxa and Species Richness in Northeast Asia. Bot. Pacifica 2012, 1, 5–20. [Google Scholar] [CrossRef]
- Ge, X.J.; Hsu, T.W.; Hung, K.H.; Lin, C.J.; Huang, C.C.; Huang, C.C.; Chiang, Y.C.; Chiang, T.Y. Inferring Multiple Refugia and Phylogeographical Patterns in Pinus massoniana Based on Nucleotide Sequence Variation and DNA Fingerprinting. PLoS ONE 2012, 7, e43717. [Google Scholar] [CrossRef] [PubMed]
- Hoover, D.L.; Knapp, A.K.; Smith, M.D. Resistance and resilience of a grassland ecosystem to climate extremes. Ecology 2014, 95, 2646–2656. [Google Scholar] [CrossRef]
- Olsen, S.L.; Töpper, J.P.; Skarpaas, O.; Vandvik, V.; Klanderud, K. From facilitation to competition: Temperature-driven shift in dominant plant interactions affects population dynamics in seminatural grasslands. Glob. Chang. Biol. 2016, 22, 1915–1926. [Google Scholar] [CrossRef] [PubMed]
- Xue, T.T.; Gadagkar, S.R.; Albright, T.P.; Yang, X.D.; Li, J.; Xia, C.Y.; Wu, J.Y.; Yu, S.X. Prioritizing conservation of biodiversity in an alpine region: Distribution pattern and conservation status of seed plants in the Qinghai-Tibetan Plateau. Glob. Ecol. Conserv. 2021, 32, e01885. [Google Scholar] [CrossRef]
- Wang, A.L.; Li, W.W. Genetic diversity of Rheum tanguticum (Polygonaceae), an endangered species on Qinghai-Tibetan Plateau. Biochem. Syst. Ecol. 2016, 69, 132–137. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- Fang, J.Y.; Zhu, J.L.; Shi, Y. The responses of ecosystems to global warming. Chin. Sci. Bull. 2018, 63, 136–140. [Google Scholar] [CrossRef]
- Cai, Q.Q.; An, M.T.; Yu, J.H.; Tian, L. Suitable habitat prediction of Pinus kwangtungensis in China under climate change. J. Shaanxi Norm. Univ. 2023, 52, 90–102. [Google Scholar] [CrossRef]
- Bálint, M.; Domisch, S.; Engelhardt, C.H.M.; Haase, P.; Lehrian, S.; Sauer, J.; Theissinger, K.; Pauls, S.U.; Nowak, C. Cryptic biodiversity loss linked to global climate change. Nat. Clim. Chang. 2011, 1, 313–318. [Google Scholar] [CrossRef]
- He, Y.Z.; Huang, W.D.; Zhao, X.; Lv, P. Review on the impact of climate change on plant diversity. J. Desert Res. 2021, 41, 59–66. [Google Scholar] [CrossRef]
- Liu, M.X.; Li, L.; Wang, S.Y.; Xiao, S.R.; Mi, J.L. Forecasting the future suitable growth areas and constructing ecological corridors for the vulnerable species Ephedra sinica in China. J. Nat. Conserv. 2023, 73, 126401. [Google Scholar] [CrossRef]
- Tracy, J.L.; Kantola, T.; Baum, K.A.; Coulson, R.N. Modeling fall migration pathways and spatially identifying potential migratory hazards for the eastern monarch butterfly. Landsc. Ecol. 2019, 34, 443–458. [Google Scholar] [CrossRef]
- Tsiftsis, S.; Djordjevic, V.; Tsiripidis, I. Neottia cordata (Orchidaceae) at its southernmost distribution border in Europe: Threat status and effectiveness of Natura 2000 Network for its conservation. J. Nat. Conserv. 2019, 48, 27–35. [Google Scholar] [CrossRef]
- Xu, W.M.; Du, Q.L.; Yan, S.; Cao, Y.; Liu, X.; Guan, D.X.; Ma, L.Q. Geographical distribution of As-hyperaccumulator Pteris vittata in China: Environmental factors and climate changes. Sci. Total Environ. 2022, 803, 149864. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.H.; Pan, B.R.; Zhuo, L.; Li, W.J.; Liu, H.L. Impact of future climate change on distribution pattern of Tamarix taklamakanensis and its conservation revelation. Arid. Zone Res. 2018, 35, 150–155. [Google Scholar] [CrossRef]
- Zhang, K.L.; Yao, L.J.; Meng, J.S.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef]
- Duan, Y.Z.; Wang, H.T.; Wang, C.; Du, Z.Y. Potential distribution of endangered plant Helianthemum songaricum in China under climate change. J. Plant Resour. Environ. 2020, 2, 55–68. [Google Scholar] [CrossRef]
- Lu, K.; He, Y.M.; Mao, W.; Zy, D.U.; Wang, L.J.; Liu, G.M.; Feng, W.J.; Duan, Y.Z. Potential geographical distribution and changes of Artemisia ordosicain China under future climate change. J. Appl. Ecol. 2020, 31, 3758–3766. [Google Scholar] [CrossRef]
- Liu, Y.; Miao, C.; Wang, H.S. Influence of climate change on distribution of suitable areas of larix plantation in China. Acta Ecol. Sin. 2023, 43, 9686–9698. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Zhao, B.J.; Huang, T.S.; Chen, H.; Yue, J.J.; Tian, Y.L. Responses of the Distribution Pattern of the Suitable Habitat of Juniperus tibetica Komarov to Climate Change on the Qinghai-Tibet Plateau. Forests 2023, 14, 434. [Google Scholar] [CrossRef]
- Ma, S.M.; Wei, B.; Li, X.C.; Ji, X.R. The impacts of climate change on the potential distribution of Haloxylon ammodendron. Chin. J. Ecol. 2017, 36, 1243. [Google Scholar] [CrossRef]
- Zhang, D.F.; Zhang, Q.; Guo, J.; Sun, C.Z.; Xie, C.X. Research on the global ecological suitability and characteristics of regions with Angelica sinensis based on the MaxEnt model. Acta Ecol. Sin. 2017, 37, 5111–5120. [Google Scholar] [CrossRef]
- Rong, W.W.; Huang, X.; Niu, P.X.; Liu, J.Q.; Yan, R.; Su, J.J.; Hu, S.C.; Chu, G.M. Potentially suitable areas for traditional Chinese medicinal material Ephedra equisetina based on MaxEnt model. Acta Ecol. Sin. 2023, 43, 8631–8646. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, P.; Zou, H.; Ji, X.; Wang, Z.; Liu, Z. Adaptive Distribution and Vulnerability Assessment of Endangered Maple Species on the Tibetan Plateau. Forests 2024, 15, 491. [Google Scholar] [CrossRef]
- Li, M.; Zhang, Y.; Yang, Y.; Wang, T.; Wu, C.; Zhang, X. Prediction of Historical, Current, and Future Configuration of Tibetan Medicinal Herb Gymnadenia orchidis Based on the Optimized MaxEnt in the Qinghai–Tibet Plateau. Plants 2024, 13, 645. [Google Scholar] [CrossRef]
- Jantz, P.; Goetz, S.; Laporte, N. Carbon stock corridors to mitigate climate change and promote biodiversity in the tropics. Nat. Clim. Chang. 2014, 4, 138–142. [Google Scholar] [CrossRef]
- Fei, Y.Q.; Huang, A.M.; Luo, Y.; Zhao, L. Ecological restoration of abandoned mines within giant panda habitat based on Maxent model and circuit theory. J. Environ. Eng. Technol. 2024, 14, 1–11. [Google Scholar] [CrossRef]
- Ye, Y.P.; Wang, S.N. Estimating urban suitable ecological land based on the minimum cumulative resistance model: A case study in Nanjing, China. In Proceedings of the IOP Conference Series: Earth and Environmental Science, Macao, China, 16–19 July 2019. [Google Scholar]
- Li, Z.; Ding, Y.; Wang, Y.L.; Chen, J.; Wu, F.M. Construction of Ecological Security Pattern in Mountain Rocky Desertification Area Based on MCR Model: A Case Study of Nanchuan, Chongqing. J. Ecol. Rural Environ. 2020, 36, 1046–1054. [Google Scholar] [CrossRef]
- Su, X.K.; Liu, S.L.; Dong, S.K.; Zhang, Y.; Wu, X.Y.; Zhao, H.D.; Zhao, Z.Z.; Sha, W. Effects of potential mining activities on migration corridors of Chiru (Pantholops hodgsonii) in the Altun National Nature Reserve, China. J. Nat. Conserv. 2015, 28, 119–126. [Google Scholar] [CrossRef]
- Zhuge, H.J.; Lin, D.Q.; Li, X.W. Identification of ecological corridors for Tibetan antelope and assessment of their human disturbances in the alpine desert of Qinghai-Tibet Plateau. Chin. J. Appl. Ecol. 2015, 26, 2504–2510. [Google Scholar] [CrossRef]
- Chen, S.L.; Gilbert, M.G. Flora of China Editorial Committee (2006) Flora of China; Science Press and Missouri Botanical Garden Press: Beijing, China, 2006; Volume 22. [Google Scholar]
- Shi, B.; Dou, J.D.; Huang, W.; Li, X.W. Community characteristics of Ephedra rhytidosperma in Helan Mountain of Ningxia, China. Chin. J. Plant Ecol. 2022, 46, 362–367. [Google Scholar] [CrossRef]
- Zhao, L.; Yang, B.; Liu, W.D.; Ma, L.; Gao, J.H. Sex ratio and spatial pattern of Ephedra rhytidosperma in Helan Mountain of Ningxia. Acta Ecol. Sin. 2022, 42, 10297–10304. [Google Scholar] [CrossRef]
- Xiu, L.M.; Miura, A.B.; Yamamoto, K.; Kobayashi, T.; Song, Q.H.; Kitamura, H.; Cyong, J.C. Pancreatic islet regeneration by ephedrine in mice with streptozotocin-induced diabetes. Am. J. Chin. Med. 2001, 29, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Tao, C.; Du, C.G. Medicinal herbs for the management of desertification. J. Agric. Biotechnol. Ecol. 2010, 3, 124–132. [Google Scholar]
- Ma, X.H.; Lu, Y.Y.; Huang, D.D.; Zhu, T.T.; Lv, P.L.; Jin, L. Ecology suitability study of Ephedra intermedia. China J. Chin. Mater. Medica 2017, 42, 2068–2071. [Google Scholar] [CrossRef]
- He, P.; Li, J.Y.; Li, Y.F.; Xu, N.; Gao, Y.; Guo, L.F.; Huo, T.T.; Peng, C.; Meng, F.Y. Habitat protection and planning for three Ephedra using the MaxEnt and Marxan models. Ecol. Indic. 2021, 133, 108399. [Google Scholar] [CrossRef]
- Nzei, J.M.; Ngarega, B.K.; Mwanzia, V.M.; Musili, P.M.; Wang, Q.F.; Chen, J.M. The past, current, and future distribution modeling of four water lilies (Nymphaea) in Africa indicates varying suitable habitats and distribution in climate change. Aquat. Bot. 2021, 173, 103416. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, Z.Y.; Abdukeyum, N.; Ling, Y.B. Potential Geographical Distribution of Medicinal Plant Ephedra sinica Stapf under Climate Change. Forests 2022, 13, 2149. [Google Scholar] [CrossRef]
- Guo, L.; Gao, Y.; He, P.; He, Y.; Meng, F. Modeling for Predicting the Potential Geographical Distribution of Three Ephedra Herbs in China. Plants 2023, 12, 787. [Google Scholar] [CrossRef]
- Cianfaglione, K.; Crișan, F.; Gafta, D. Soil Quality Enhances Seed Germination Success in Ephedra major—A Pilot Experiment. Plants 2023, 12, 438. [Google Scholar] [CrossRef] [PubMed]
- Remya, K.; Ramachandran, A.; Jayakumar, S. Predicting the current and future suitable habitat distribution of Myristica dactyloides Gaertn. using MaxEnt model in the Eastern Ghats, India. Ecol. Eng. 2015, 82, 184–188. [Google Scholar] [CrossRef]
- Sun, J.; Wang, N.A.; Niu, Z. Effect of Soil Environment on Species Diversity of Desert Plant Communities. Plants 2023, 12, 3465. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Cao, G.C.; Rong, Z.L.; Li, H.F. Prediction of potential distribution of Ophiocordyceps sinensis in Chain based on Maxent model. Ecol. Sci. 2023, 42, 202–212. [Google Scholar] [CrossRef]
- Chen, M.Y.; Zhang, L.L.; Cao, B.; Bai, C.K. Predictive suitable distributions and principal environment variables of six medicinal plants of Scutellaria based on MaxEnt and GIS. J. Tradit. Chin. Vet. Med. 2022, 41, 7–11. [Google Scholar] [CrossRef]
- Song, H.; Feng, G.; Tian, C.Y.; Zhang, F.S. Osmotic adjustment traits of Suaeda physophora, Haloxylon ammodendron and Haloxylon persicum in field or controlled conditions. Plant Sci. 2006, 170, 113–119. [Google Scholar] [CrossRef]
- Jiang, L.; Wu, D.; Li, W.; Liu, Y.; Li, E.; Li, X.; Yang, G.; He, X. Variations in Physiological and Biochemical Characteristics of Kalidium foliatum Leaves and Roots in Two Saline Habitats in Desert Region. Forests 2024, 15, 148. [Google Scholar] [CrossRef]
- Cantón, Y.; Del Barrio, G.; Solé-Benet, A.; Lázaro, R. Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain. Catena 2004, 55, 341–365. [Google Scholar] [CrossRef]
- Polley, H.W.; Tischler, C.R.; Jobnson, H.B. Elevated atmospheric CO2 magnifies intra-specific variation in seedling growth of honey mesquite: An assessment of relative growth rates. Rangel. Ecol. Manag. 2006, 59, 128–134. [Google Scholar] [CrossRef]
- Wang, Y.X.; Chen, X.J.; Lou, S.N.; Hu, A.; Ren, J.F. Woody-plant encroachment in grasslands: A review of mechanisms and aftereffects. Acta Pratacult. Sin. 2018, 27, 219–227. [Google Scholar] [CrossRef]
- Kang, H.M.; Li, H.H.; Xu, D.H.; Qin, P.; Liu, M.L. Effects of elevated CO2 and temperature on Caragana jubata (Alpineshrub) growth and antioxidant systems. Acta Ecol. Sin. 2020, 40, 367–376. [Google Scholar] [CrossRef]
- Hong, C.C.; Wang, B.Z.; Bai, Y.H.; Zhu, Y.J.; Zhang, K.B.; Yang, X.H. Spatial and temporal variations of potential suitable areas of caragana roborovskyi in China under climate change scenarios. Acta Bot. Boreali-Occident. Sin. 2023, 43, 0856–0866. [Google Scholar] [CrossRef]
- Schöb, C.; Kammer, P.M.; Choler, P.; Veit, H. Small-scale plant species distribution in snowbeds and its sensitivity to climate change. Plant Ecol. 2009, 200, 91–104. [Google Scholar] [CrossRef]
- Wang, C.H.; Zhang, S.N.; Zhang, F.M.; Li, K.C.; Yang, K. On the increase of precipitation in the Northwestern China under the global warming. Adv. Earth Sci. 2021, 36, 980–989. [Google Scholar] [CrossRef]
- Li, B.F.; Chen, Y.N.; Chen, Z.S.; Xiong, H.G.; Lian, L.S. Why does precipitation in northwest China show a significant increasing trend from 1960 to 2010? Atmos. Res. 2016, 167, 275–284. [Google Scholar] [CrossRef]
- Fan, X.W.; Miao, C.Y.; Duan, Q.Y.; Shen, C.W.; Wu, Y. The Performance of CMIP6 Versus CMIP5 in Simulating Temperature Extremes Over the Global Land Surface. J. Geophys. Res.-Atmos. 2020, 125, e2020JD033031. [Google Scholar] [CrossRef]
- Root, T.L.; Price, J.T.; Hall, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J.A. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef]
- Xu, M.L.; Xue, X. Analysis on the effects of climate warming on growth and phenology of alpine plants. J. Arid. Land Resour. Environ. 2013, 27, 137–141. [Google Scholar] [CrossRef]
- Wang, D.; You, Q.L.; Jiang, Z.H.; Wu, W.B.; Jiao, Y. Analysis of extreme temperature changes in China based on the homogeneity-adjusteddata. Plateau Meteorol. 2016, 35, 1352–1363. [Google Scholar] [CrossRef]
- Fan, X.; Pan, J.W.; He, S.T. Prediction of the potential distribution of Rosa roxburghii under the background of climate change based on MaxEnt model. Acta Bot. Boreali-Occident. Sin. 2021, 41, 159–167. [Google Scholar] [CrossRef]
- Li, Q.W.; Li, S.; Cao, M.C.; Xu, H.G. Habitat Suitability Evaluation and Corridor Design of Muntiacus crinifrons in Qianjiangyuan National Park. J. Ecol. Rural. Environ. 2021, 37, 778–785. [Google Scholar] [CrossRef]
- Li, Q.L. Community Characteristics and Conservation Strategies of Endangered Plant adiantum nelumboides X. C. Zhang in Three Gorges Reservoir Region; Southwest University: Chongqing, China, 2018. [Google Scholar]
- Zhao, Y.F.; Pan, B.R. Potential geographical distributions of Tugarinovia in China under climate change scenarios. Arid. Zone Res. 2023, 40, 949–957. [Google Scholar] [CrossRef]
- Zhai, X.Y.; Shen, Y.F.; Zhu, S.H.; Tu, Z.H. Potential impacts of climate change in future on the geographical distributions of relic Liriodendron chinense. J. Trop. Subtrop. Bot. 2021, 29, 151–161. [Google Scholar] [CrossRef]
- Li, F.; Ye, Y.P.; Song, B.W.; Wang, R.S. Evaluation of urban suitable ecological land based on the minimum cumulative resistance model: A case study from Changzhou, China. Ecol. Model. 2015, 318, 194–203. [Google Scholar] [CrossRef]
- Li, R.; Jiao, Y.M.; Liu, X.; Liu, Z.L.; Gao, X. Suitability evaluation and corridor design of habitats for Green Peafowl based on MaxEnt Model. Chin. J. Ecol. 2019, 38, 919–926. [Google Scholar] [CrossRef]
- Akbaba, B.; Bulut, S. Inventory of Large Mammal Species in the Ilgaz Mountains (ankr): A Major Ecological Corridor in Anatolia. Hitte J. Sci. Eng. 2020, 7, 73–80. [Google Scholar] [CrossRef]
- Kong, L.Q.; Xu, W.H.; Xiao, Y.; Pimm, S.L.; Shi, H.; Ouyang, Z.Y. Spatial models of giant pandas under current and future conditions reveal extinction risks. Nat. Ecol. Evol. 2021, 5, 1309. [Google Scholar] [CrossRef]
- Yang, W.; Tong, Y.F.; Ma, T.; Wu, G.L.; Li, S.P. Study on characters of N, P and K of aeolian soil and fertilization for Ephedra sinica growing base. Pratacult. Sci. 2008, 25, 19–25. [Google Scholar]
- Shi, N.N.; Han, Y.; Wang, Q.; Quan, Z.J.; Luo, Z.L. Construction and optimization of ecological network for protected areas in Qinghai province. Chin. J. Ecol. 2018, 37, 1910–1916. [Google Scholar] [CrossRef]
- Rushdi, A.M.A.; Hassan, A.K. Reliability of migration between habitat patches with heterogeneous ecological corridors. Ecol. Model. 2015, 304, 1–10. [Google Scholar] [CrossRef]
- Li, L.; Shi, Z.H.; Yin, W.; Zhu, D.; Ng, S.L.; Cai, C.F.; Lei, A.L. A fuzzy analytic hierarchy process (FAHP) approach to eco-environmental vulnerability assessment for the danjiangkou reservoir area, China. Ecol. Model. 2009, 220, 3439–3447. [Google Scholar] [CrossRef]
- Tang, Y.H.; Gao, C.; Wu, X.F. Urban Ecological Corridor Network Construction: An Integration of the Least Cost Path Model and the InVEST Model. ISPRS Int. Geo-Inf. 2020, 9, 33. [Google Scholar] [CrossRef]
- Perrino, E.V.; Mahmoud, Z.N.A.; Valerio, F.; Tomaselli, V.; Wagensommer, R.P.; Trani, A. Synecology of Lagoecia cuminoides L. in Italy and evaluation of functional compounds presence in its water or hydroalcoholic extracts. Sci. Rep. 2023, 13, 20906. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.K.; Qin, H.N.; Jin, X.H.; Zhang, Z.X.; Yang, Q.W.; Hong, D.Y.; Li, D.Z.; Li, K.F.; Yuan, L.C.; Zhou, Z.H. On the necessity, principle, and process of updating the List of National Key Protected Wild Plants. Biodivers. Sci. 2021, 29, 1577–1582. [Google Scholar] [CrossRef]
- Wang, S.; Xie, Y. China Species Red List; Higher Education Press: Beijing, China, 2004. [Google Scholar]
- Warren, D.L.; Beaumont, L.J.; Dinnage, R.; Baumgartner, J.B. New methods for measuring ENM breadth and overlap in environmental space. Ecography 2019, 42, 444–446. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 2010, 33, 607–611. [Google Scholar] [CrossRef]
- Yang, H.F.; Zhong, X.N.; Deng, S.Q.; Xu, H. Assessment of the impact of LUCC on NPP and its influencing factors in the Yangtze River basin, China. Catena 2021, 206, 105542. [Google Scholar] [CrossRef]
- Moreno, R.; Zamora, R.; Molina, J.R.; Vasquez, A.; Herrera, M.A. Predictive modeling of microhabitats for endemic birds in South Chilean temperate forests using Maximum entropy (Maxent). Ecol. Inform. 2011, 6, 364–370. [Google Scholar] [CrossRef]
- Ward, D.F. Modelling the potential geographic distribution of invasive ant species in New Zealand. Biol. Invasions 2007, 9, 723–735. [Google Scholar] [CrossRef]
- Yang, Z.B.; Bai, Y.; Alatalo, J.M.; Huang, Z.D.; Yang, F.; Pu, X.Y.; Wang, R.B.; Yang, W.; Guo, X.Y. Spatio-temporal variation in potential habitats for rare and endangered plants and habitat conservation based on the maximum entropy model. Sci. Total Environ. 2021, 784, 147080. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yuan, H.; Zou, H.; Zhu, X.; Zhang, Y.; Wang, Z.; Liu, Z. Global Warming Drives Expansion of Endangered Spruce Forest on the Tibetan Plateau. Sustainability 2024, 16, 2164. [Google Scholar] [CrossRef]
- Chen, Y.G.; Le, X.G.; Chen, Y.H.; Chen, W.X.; Du, J.G.; Zhong, Q.L.; Cheng, D.L. Identification of the potential distribution area of Cunninghamia lanceolata in China under climate change based on the MaxEnt model. Chin. J. Appl. Ecol. 2022, 33, 1207–1214. [Google Scholar] [CrossRef]
- Van der Windt, H.J.; Swart, J.A.A. Ecological corridors, connecting science and politics: The case of the Green River in the Netherlands. J. Appl. Ecol. 2008, 45, 124–132. [Google Scholar] [CrossRef]
- Teng, Y.; Zhang, Z.; Zhang, S.L.; Yang, Y.X.; He, W.; Wang, N.; Zhang, Z.Y.; Bao, W.D. Habital suitabilility analysis and ecological corridor designs for red deer (Cervus elaphus) in the southern Greater Khingan Mountains. Acta Ecol. Sin. 2022, 42, 5990–6000. [Google Scholar] [CrossRef]
- Bastille-Rousseau, G.; Wittemyer, G. Characterizing the Landscape of Movement to Identify Critical Wildlife Habitat and Corridors; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2021. [Google Scholar] [CrossRef]
- Brandao, M.M.; Vieira, F.D.; Nazareno, A.G.; de Carvalho, D. Genetic diversity of neotropical tree Myrcia splendens (Myrtaceae) in a fragment-corridor system in the Atlantic rainforest. Flora 2015, 216, 35–41. [Google Scholar] [CrossRef]
- Yao, C.Y.; He, Y.M.; Cheng, J.X.; Zhang, T.Y.; Pan, H.Y.; Ma, H.J. Evaluation of ecological security pattern and optimization suggestions in MinjiangRiver Basin based on MCR model and gravity model. Acta Ecol. Sin. 2023, 43, 7083–7096. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Generally Suitable Areas /×104 km2 | Moderately Suitable Areas /×104 km2 | Highly Suitable Areas /×104 km2 | Total Suitable Areas | |
|---|---|---|---|---|---|
| /×104 km2 | Percentage/% | ||||
| E. equisetina | 107.90 | 47.29 | 16.40 | 171.60 | 17.9 |
| E. intermedia | 134.55 | 62.24 | 25.30 | 222.10 | 16.1 |
| E. sinica | 89.77 | 50.84 | 14.11 | 154.72 | 16.1 |
| E. monosperma | 124.43 | 34.16 | 30.51 | 161.64 | 16.8 |
| E. rhytidosperma | 1.79 | 2.05 | 0.64 | 4.48 | 0.05 |
| Species | Driving Factor | Contribution |
|---|---|---|
| E. equisetina | T_CACO3: topsoil calcium carbonate | 16 |
| Elevation | 14.5 | |
| Bio6: min temperature of coldest month | 11.7 | |
| Bio2: mean diurnal range | 10.4 | |
| Bio12: annual precipitation | 10 | |
| E. intermedia | Elevation | 19.8 |
| T_BS: topsoil base saturation | 14.2 | |
| Bio15: precipitation seasonality | 10.6 | |
| Bio9: mean temperature of driest quarter | 9.5 | |
| T_CACO3: topsoil calcium carbonate | 7.8 | |
| E. sinica | Bio13: precipitation of wettest month | 19.1 |
| T_CACO3: topsoil calcium carbonate | 15.8 | |
| Bio19: precipitation of coldest quarter | 13 | |
| Elevation | 10.9 | |
| Bio15: precipitation seasonality | 10.9 | |
| E. monosperma | Elevation | 33.1 |
| Bio2: mean diurnal range | 12.4 | |
| Slope | 8.9 | |
| Bio19: precipitation of coldest quarter | 8.8 | |
| Bio12: annual precipitation | 6.7 | |
| E. rhytidosperma | T_PH_H2O: topsoil pH (H2O) | 38.6 |
| Bio12: annual precipitation | 15.5 | |
| Bio11: mean temperature of coldest quarter | 13.3 | |
| Aspect | 10.2 | |
| T_CASO4: topsoil gypsum | 7.9 |
| Symbol | Environmental Factors | Unit | Symbol | Environmental Factors | Unit |
|---|---|---|---|---|---|
| Bio1 | Annual mean temperature | °C | Bio19 | Precipitation of coldest quarter | mm |
| Bio2 | Mean diurnal range | °C | Elev | Elevation | m |
| Bio3 | Isothermality | \ | Aspect | Aspect | ° |
| Bio4 | Temperature seasonality | \ | Slope | Slope | ° |
| Bio5 | Max temperature of warmest month | °C | T_PH_H2O | Topsoil pH (H2O) | −log (H+) |
| Bio6 | Min temperature of coldest month | °C | T_GRAVEL | Topsoil gravel content | % |
| Bio7 | Temperature annual range | °C | T_SILT | Topsoil silt fraction | % |
| Bio8 | Mean temperature of wettest quarter | °C | T_CLAY | Topsoil clay fraction | % |
| Bio9 | Mean temperature of driest quarter | °C | T_SAND | Topsoil sand fraction | % |
| Bio10 | Mean temperature of warmest quarter | °C | T_OC | Topsoil organic carbon | % |
| Bio11 | Mean temperature of coldest quarter | °C | T_CEC_CLAY | Topsoil CEC (clay) | cmol/kg |
| Bio12 | Annual precipitation | mm | T_CEC_SOIL | Topsoil CEC (soil) | cmol/kg |
| Bio13 | Precipitation of wettest month | mm | T_BS | Topsoil base saturation | % |
| Bio14 | Precipitation of driest month | mm | T_TEB | Topsoil TEB | cmol/kg |
| Bio15 | Precipitation seasonality | \ | T_CACO3 | Topsoil calcium carbonate | % |
| Bio16 | Precipitation of wettest quarter | mm | T_CASO4 | Topsoil gypsum | % |
| Bio17 | Precipitation of driest quarter | mm | T_ESP | Topsoil sodicity (ESP) | % |
| Bio18 | Precipitation of warmest quarter | mm | T_ECE | Topsoil salinity (Elco) | dS/m |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Li, J.; Zou, H.; Wang, Z.; Zhu, X.; Zhang, Y.; Liu, Z. Distribution Pattern of Suitable Areas and Corridor Identification of Endangered Ephedra Species in China. Plants 2024, 13, 890. https://doi.org/10.3390/plants13060890
Zhang H, Li J, Zou H, Wang Z, Zhu X, Zhang Y, Liu Z. Distribution Pattern of Suitable Areas and Corridor Identification of Endangered Ephedra Species in China. Plants. 2024; 13(6):890. https://doi.org/10.3390/plants13060890
Chicago/Turabian StyleZhang, Huayong, Jiangpeng Li, Hengchao Zou, Zhongyu Wang, Xinyu Zhu, Yihe Zhang, and Zhao Liu. 2024. "Distribution Pattern of Suitable Areas and Corridor Identification of Endangered Ephedra Species in China" Plants 13, no. 6: 890. https://doi.org/10.3390/plants13060890
APA StyleZhang, H., Li, J., Zou, H., Wang, Z., Zhu, X., Zhang, Y., & Liu, Z. (2024). Distribution Pattern of Suitable Areas and Corridor Identification of Endangered Ephedra Species in China. Plants, 13(6), 890. https://doi.org/10.3390/plants13060890