

Evaluating Guayule (Parthenium argentatum A. Gray) Germplasm Grown in Spain: Rubber and Resin along Three Production Cycles
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
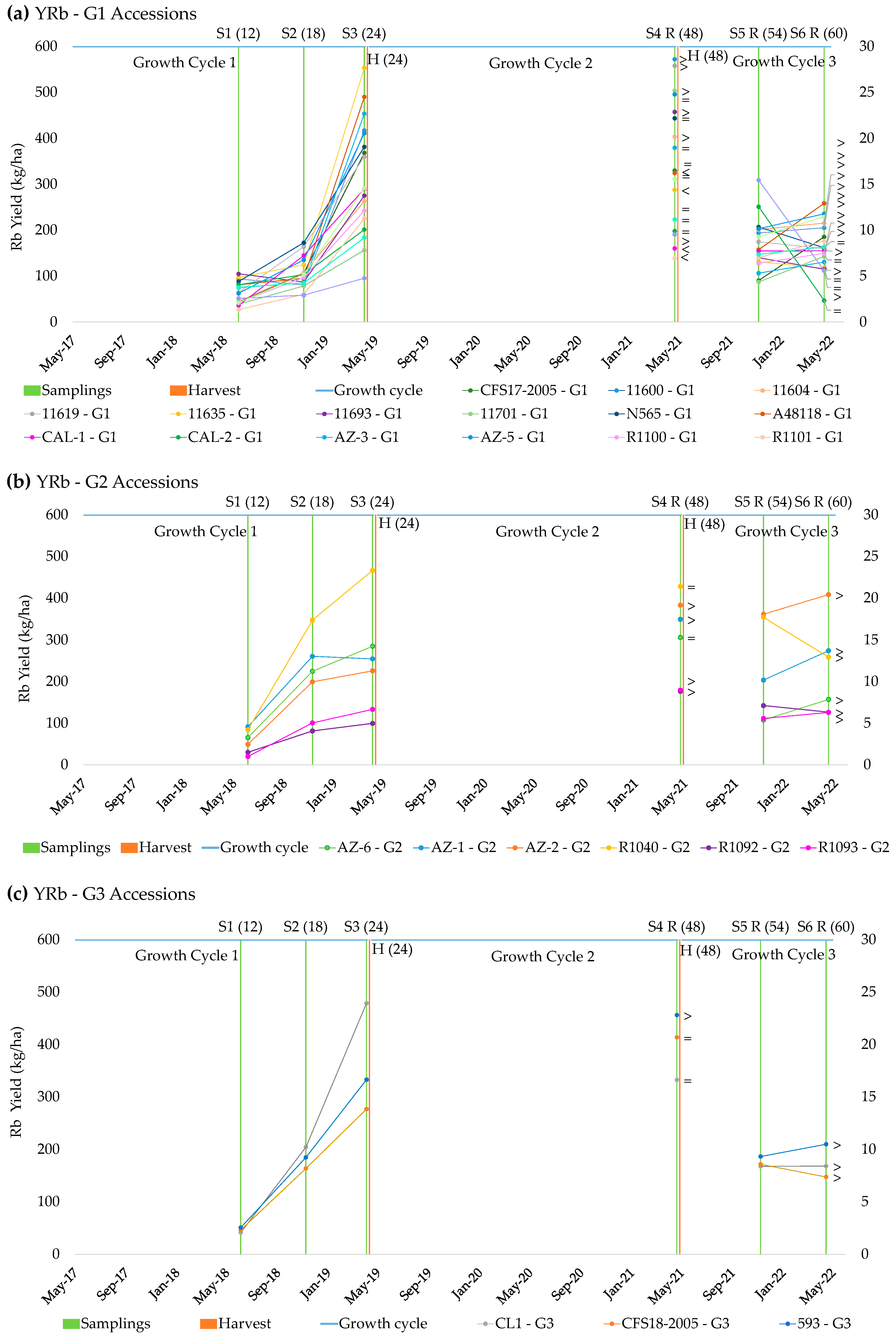
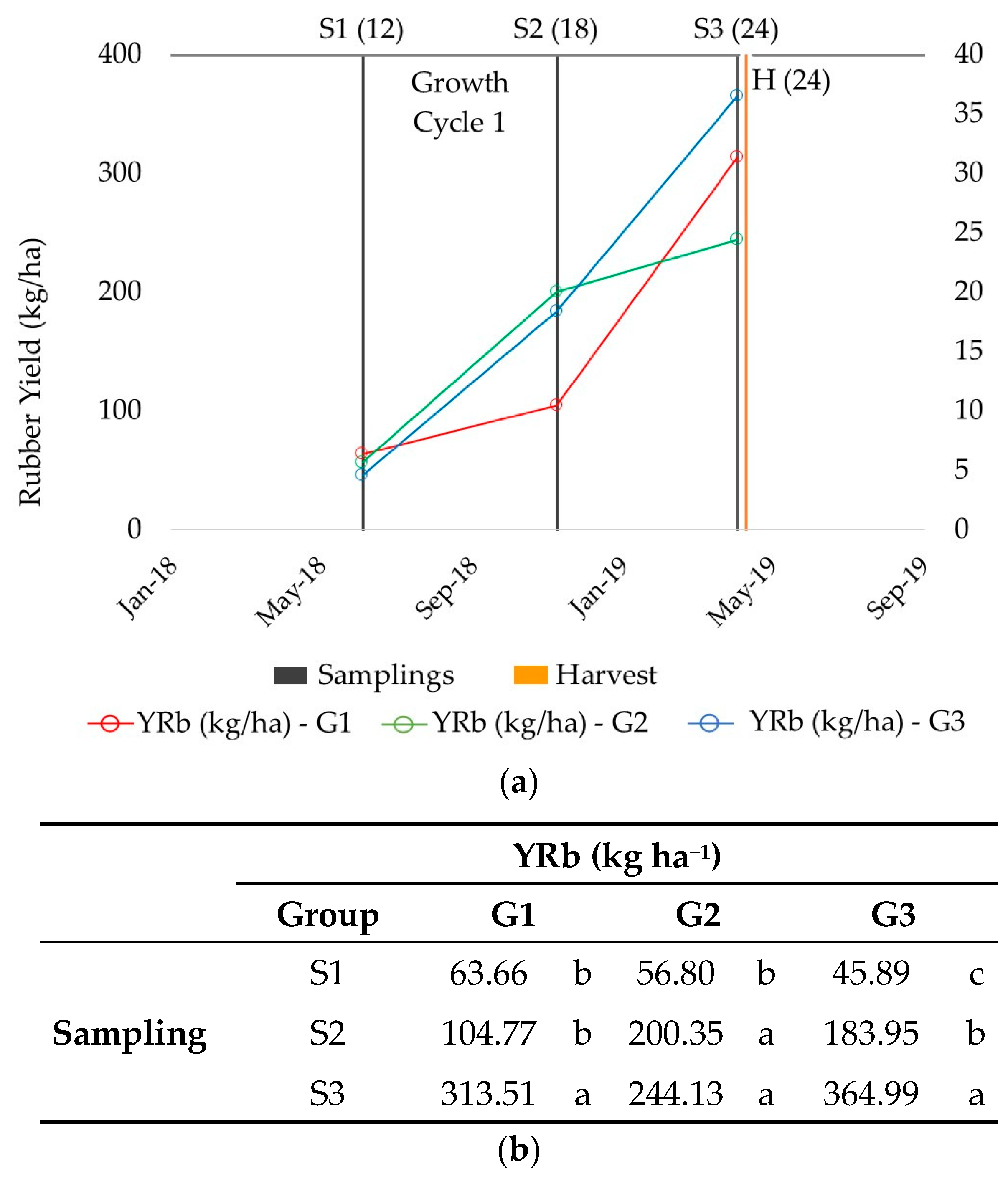
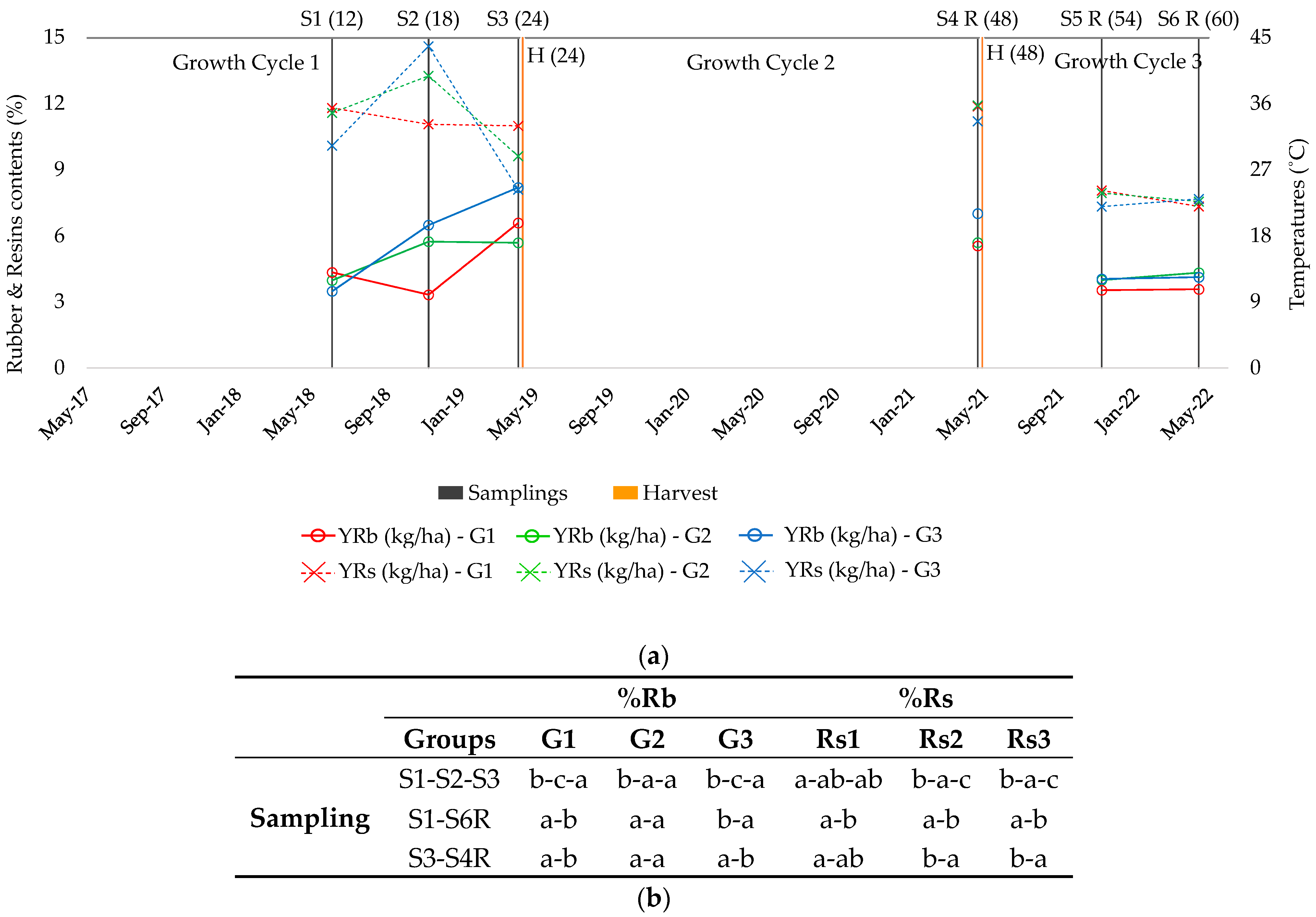
2.1. Rubber Production
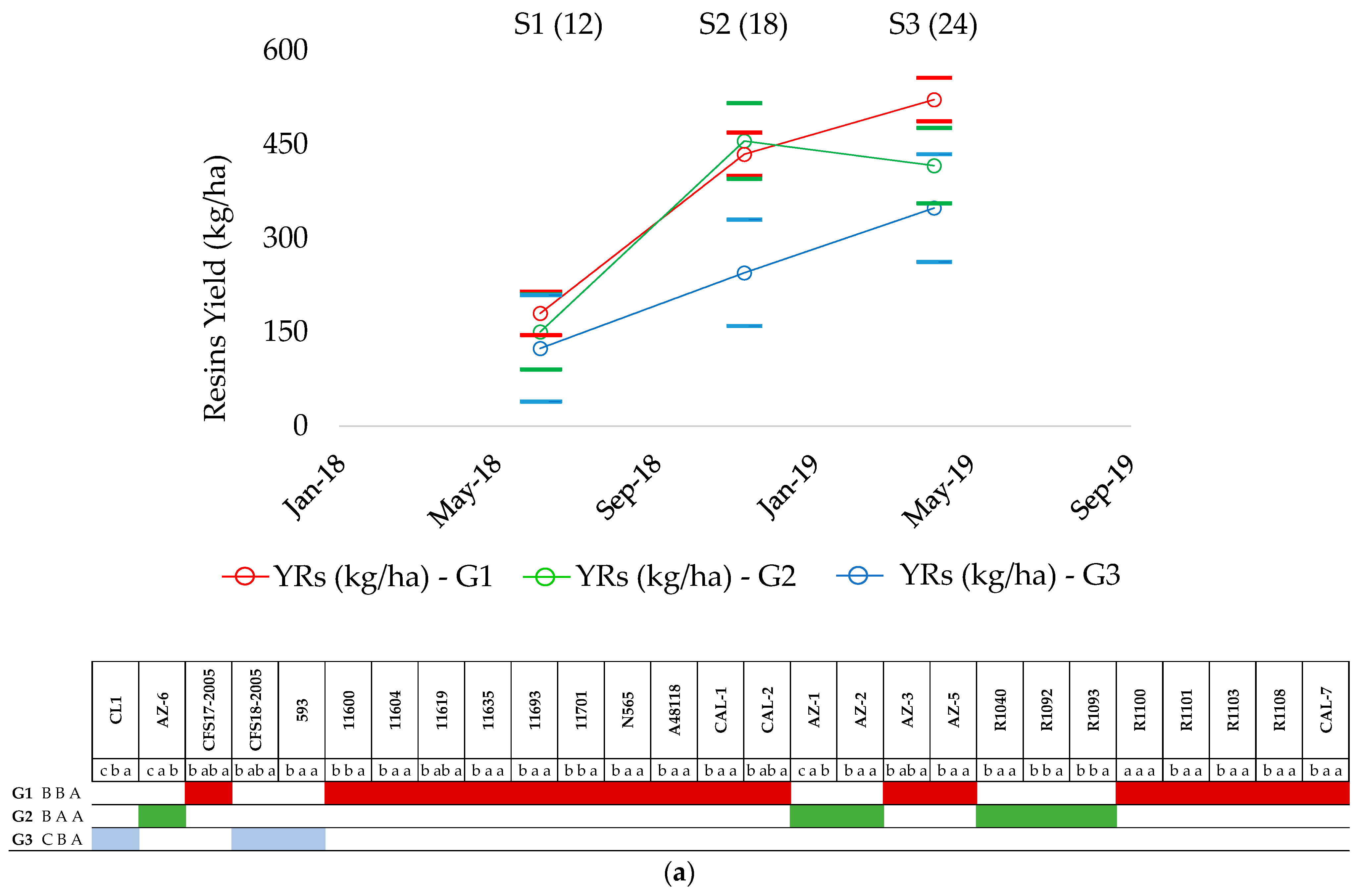
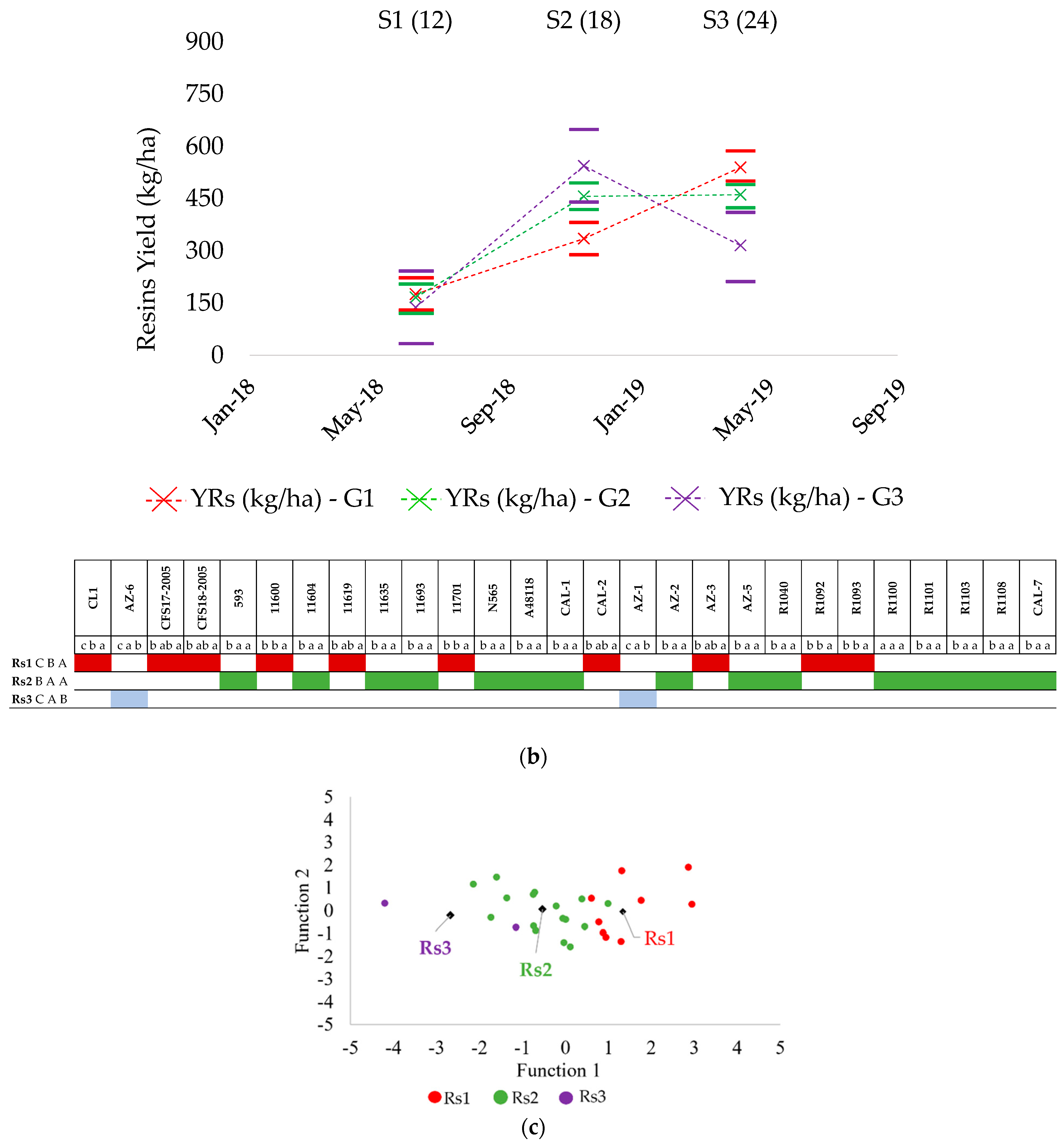
2.2. Resin Production
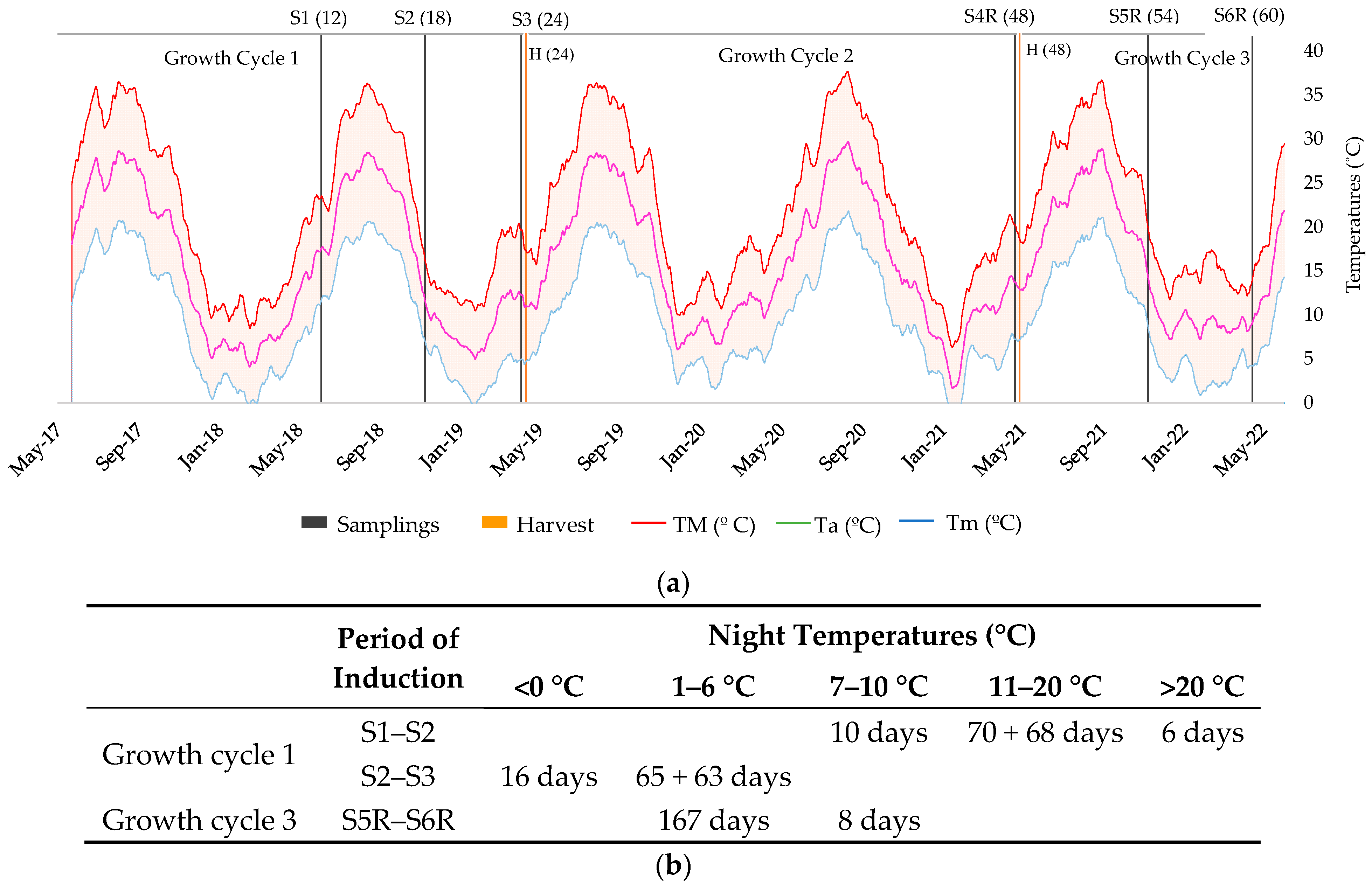
2.3. Patterns of Rubber Production
2.4. Patterns of Resin Production
2.5. Concentration of Rubber and Resin
3. Materials and Methods
3.1. Germplasm, Sampling and Monitoring
3.2. Sampling and Processing
3.3. Resin and Natural Rubber Determination
3.4. Data Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Halima, M.; Abderrahmane, L.; Khéloufi, B. Essai sur le rôle d’une espèce végétale rustique pour un développement durable de la steppe algérienne. Dév. Durable Territ. 2006, 1–15. [Google Scholar] [CrossRef]
- Jara, F.M.; López-Córcoles, H.; Carrión, M.E.; Zalacain, A.; Carmona, M. Phenotypic variability of guayule (Parthenium argentatum Gray) germplasm grown in Castilla-La Mancha (Spain) (I): Morphology and biomass production. Ind. Crop. Prod. 2023, 203, 117126. [Google Scholar] [CrossRef]
- Coffelt, T.A.; Nakayama, F.S.; Ray, D.T.; Cornish, K.; McMahan, C.M. Post-harvest storage effects on guayule latex, rubber, and resin contents and yields. Ind. Crop. Prod. 2009, 29, 326–335. [Google Scholar] [CrossRef]
- George, D.; Gupta, M.; Dissanayake, P. Evaluating New Guayule Varieties for Low-Allergenic Rubber Production; Australian Goverment Rural Industries Research and Development Corporations: Canberra, Australia, 2005. [Google Scholar]
- Clarke, M. Australian Guayule Industry Strategic RD & E Plan Australian Guayule Industry Strategic RD & E Plan; AgriFutures Australia: Wagga Wagga, NSW, Australia, 2022. [Google Scholar]
- Dehghanizadeh, M.; Mendoza Moreno, P.; Sproul, E.; Bayat, H.; Quinn, J.C.; Brewer, C.E. Guayule (Parthenium argentatum) resin: A review of chemistry, extraction techniques, and applications. Ind. Crop. Prod. 2021, 165, 113410. [Google Scholar] [CrossRef]
- Sfeir, N.; Chapuset, T.; Palu, S.; Lançon, F.; Amor, A.; García García, J.; Snoeck, D. Technical and economic feasibility of a guayule commodity chain in Mediterranean Europe. Ind. Crop. Prod. 2014, 59, 55–62. [Google Scholar] [CrossRef]
- Rasutis, D.; Soratana, K.; McMahan, C.; Landis, A.E. A sustainability review of domestic rubber from the guayule plant. Ind. Crop. Prod. 2015, 70, 383–394. [Google Scholar] [CrossRef]
- Abdel-Haleem, H.; Foster, M.; Ray, D.; Coffelt, T. Phenotypic variations, heritability and correlations in dry biomass, rubber and resin production among guayule improved germplasm lines. Ind. Crop. Prod. 2018, 112, 691–697. [Google Scholar] [CrossRef]
- Dissanayake, P.; George, D.L.; Gupta, M.L. Performance of improved guayule lines in Australia. Ind. Crop. Prod. 2004, 20, 331–338. [Google Scholar] [CrossRef]
- Jara, F.M.; Cornish, K.; Carmona, M. Potential applications of guayulins to improve feasibility of guayule cultivation. Agronomy 2019, 9, 804. [Google Scholar] [CrossRef]
- Banigan, T.F.; Verbiscar, A.J.; Weber, C.W. Composition of guayule leaves, seed, and wood. J. Agric. Food Chem. 1982, 30, 427–431. [Google Scholar] [CrossRef]
- Schloman, W.W.; Garrot, D.J.; Ray, D.T.; Bennett, D.J. Seasonal effects on guayule resin composition. J. Agric. Food Chem. 1986, 34, 177–179. [Google Scholar] [CrossRef]
- Bonner, J. Effect of Varying Nutritional Treatments on Growth and Rubber Accumulation in Guayule. Bot. Gaz. 1944, 105, 352–364. [Google Scholar] [CrossRef]
- Goss, R.A.; Benedict, C.R.; Keithly, J.H.; Nessler, C.L.; Stipanovic, R.D. cis-Polyisoprene Synthesis in Guayule Plants (Parthenium argentatum Gray) Exposed to Low, Nonfreezing Temperatures. Plant Physiol. 1984, 74, 534–537. [Google Scholar] [CrossRef] [PubMed]
- Sundar, D.; Reddy, A.R. Interactive influence of temperature and growth light intensity on rubber accumulation and rubber transferase activity in guayule (Parthenium argentatum Gray). J. Plant Physiol. 2001, 158, 1291–1297. [Google Scholar] [CrossRef]
- Appleton, M.R.; Van Staden, J. Influence of temperature and daylength on growth and isoprenoid biosynthesis in guayule under controlled environmental conditions. Bioresour. Technol. 1991, 35, 147–152. [Google Scholar] [CrossRef]
- Appleton, M.R.; van Staden, J. The Relationship between Season, Growth and Isoprenoid Biosynthesis in Parthentum argentatum. J. Plant Physiol. 1989, 134, 524–532. [Google Scholar] [CrossRef]
- Wan, J.; Benedict, C.R.; Foster, M.A. Seasonal variations in rubber biosynthesis, 3-hydroxy-3-methylglutaryl-coenzyme A reductase, and rubber transferase activities in Parthenium argentatum in the Chihuahuan Desert. Plant Physiol. 1993, 103, 535–542. [Google Scholar] [CrossRef]
- Cornish, K.; Backhaus, R.A. Induction of rubber transferase activity in guayule (Parthenium argentatum Gray) by low temperatures. Ind. Crop. Prod. 2003, 17, 83–92. [Google Scholar] [CrossRef]
- Majeau, G.H.; Ray, D.T.; Coffelt, T.A.; Foster, M.A. New guayule (Parthenium argentatum Gray) variety trials. In Proceedings of the Association for the Advancement of Industrial Crops, Annual Meeting Abstracts, Portland, OR, USA, 12–15 October 2003; p. 14. [Google Scholar]
- Foster, M.A.; Coffelt, T.A.; Petty, A.K. Guayule production on the southern high plains. Ind. Crop. Prod. 2011, 34, 1418–1422. [Google Scholar] [CrossRef]
- Ilut, D.C.; Sanchez, P.L.; Coffelt, T.A.; Dyer, J.M.; Jenks, M.A.; Gore, M.A. A century of guayule: Comprehensive genetic characterization of the US national guayule (Parthenium argentatum A. Gray) germplasm collection. Ind. Crop. Prod. 2017, 109, 300–309. [Google Scholar] [CrossRef]
- Dierig, D.A.; Ray, D.T.; Coffelt, T.A.; Nakayama, F.S.; Leake, G.S.; Lorenz, G. Heritability of height, width, resin, rubber, and latex in guayule (Parthenium argentatum). Ind. Crop. Prod. 2001, 13, 229–238. [Google Scholar] [CrossRef]
- Ramos, J.L. El Cultivo del Guayule en España. Mater. Veg. 1954, 1, 363–368. [Google Scholar] [CrossRef]
- Soratana, K.; Rasutis, D.; Azarabadi, H.; Eranki, P.L.; Landis, A.E. Guayule as an alternative source of natural rubber: A comparative life cycle assessment with Hevea and synthetic rubber. J. Clean. Prod. 2017, 159, 271–280. [Google Scholar] [CrossRef]
- Elshikha, D.E.M.; Waller, P.M.; Hunsaker, D.J.; Thorp, K.R.; Wang, G.; Dierig, D.; Cruz, V.M.V.; Attalah, S.; Katterman, M.E.; Williams, C.; et al. Water Use, Growth, and Yield of Ratooned Guayule under Subsurface Drip and Furrow Irrigation in the US Southwest Desert. Water 2023, 15, 3412. [Google Scholar] [CrossRef]
- Elshikha, D.E.M.; Waller, P.M.; Hunsaker, D.J.; Dierig, D.; Wang, G.; Cruz, V.M.V.; Thorp, K.R.; Katterman, M.E.; Bronson, K.F.; Wall, G.W. Growth, water use, and crop coefficients of direct-seeded guayule with furrow and subsurface drip irrigation in Arizona. Ind. Crop. Prod. 2021, 170, 113819. [Google Scholar] [CrossRef]
- Hunsaker, D.J.; Elshikha, D.M. Surface irrigation management for guayule rubber production in the US desert Southwest. Agric. Water Manag. 2017, 185, 43–57. [Google Scholar] [CrossRef]
- Wang, G.S.; Elshikha, D.E.M.; Katterman, M.E.; Sullivan, T.; Dittmar, S.; Cruz, V.M.V.; Hunsaker, D.J.; Waller, P.M.; Ray, D.T.; Dierig, D.A.; et al. Irrigation Effects on Seasonal Growth and Rubber Production of Direct-seeded Guayule. Ind. Crop. Prod. 2022, 177, 114442. [Google Scholar] [CrossRef]
- Coffelt, T.A.; Ray, D.T. Cutting height effects on guayule latex, rubber, and resin yields. Ind. Crop. Prod. 2010, 32, 264–268. [Google Scholar] [CrossRef]
- Reche-Vilches, J.A.; Jara, F.M.; Vicente, D.; López-Córcoles, H.; Zalacain, A.; Carmona, M. Cold resistance of guayule cultivated in Castilla-La Mancha, Spain. In Proceedings of the AAIC 32nd Annual Meeting, Bologna, Italy, 5–8 September 2021; p. 67. [Google Scholar]
- Rozalén, J.; García-Martínez, M.M.; Carrión, M.E.; Carmona, M.; López-Córcoles, H.; Cornish, K.; Zalacain, A. Adapting the accelerated solvent extraction method for resin and rubber determination in guayule using the BÜCHI speed extractor. Molecules 2021, 26, 183. [Google Scholar] [CrossRef]
- Templeton, G.F. A two-step approach for transforming continuous variables to normal: Implications and recommendations for IS research. Commun. Assoc. Inf. Syst. 2011, 28, 41–58. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rb (%) | YRb (kg ha−1) | Rs (%) | YRs (kg ha−1) | |||
|---|---|---|---|---|---|---|
| New plantation | ||||||
| Month 13 | S1 | mean | 4.17 | 60.16 | 11.55 | 167.48 |
| min | 1.84 | 14.25 | 6.81 | 45.64 | ||
| max | 8.52 | 154.82 | 16.25 | 398.27 | ||
| LSD | 0.35 | 4.41 | 0.54 | 10.56 | ||
| Month 18 | S2 | mean | 4.21 | 134.60 | 12.54 | 421.85 |
| min | 1.76 | 3.11 | 6.89 | 11.08 | ||
| max | 8.11 | 500.73 | 19.36 | 1707.50 | ||
| LSD | 0.40 | 13.68 | 0.66 | 40.35 | ||
| Month 24 | S3 | mean | 6.56 | 303.59 | 10.01 | 479.12 |
| min | 1.71 | 2.43 | 5.52 | 6.86 | ||
| max | 11.60 | 1251.14 | 17.14 | 2415.19 | ||
| LSD | 0.54 | 36.06 | 0.52 | 58.41 | ||
| First harvest Month 25 | ||||||
| Regrowth Month 48 | S4R | mean | 5.74 | 341.02 | 11.84 | 720.21 |
| min | 2.54 | 13.75 | 7.18 | 39.44 | ||
| max | 9.10 | 1315.35 | 19.93 | 5079.13 | ||
| LSD | 0.41 | 31.14 | 0.58 | 73.04 | ||
| Second harvest Month 49 | ||||||
| Regrowth Month 56 | S5R | mean | 4.70 | 233.85 | 10.38 | 504.00 |
| min | 2.00 | 6.97 | 6.96 | 13.54 | ||
| max | 6.94 | 1974.77 | 13.21 | 3015.00 | ||
| LSD | 0.31 | 31.87 | 0.35 | 56.11 | ||
| Month 60 | S6R | mean | 3.80 | 174.42 | 7.48 | 348.93 |
| min | 0.87 | 5.19 | 3.85 | 17.57 | ||
| max | 6.36 | 757.21 | 10.24 | 1694.04 | ||
| LSD | 0.27 | 18.24 | 0.34 | 40.26 | ||
| Accessions | Rb (%) | YRb (kg ha−1) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S1 | S2 | S3 | S4R | S5R | S6R | S1 | S2 | S3 | S4R | S5R | S6R | |
| CFS17-2005 | 4.14 | 4.69 | 8.81 | 6.87 | 3.17 | 5.41 | 49.17 | 84.90 | 463.60 | 310.81 | 64.60 | 148.18 |
| CFS18-2005 | 3.54 | 6.15 | 6.73 | 6.53 | 3.84 | 3.10 | 52.54 | 139.44 | 265.50 | 345.52 | 136.10 | 157.05 |
| 593 | 3.76 | 6.49 | 7.56 | 7.27 | 4.17 | 4.88 | 51.64 | 191.65 | 329.04 | 393.68 | 190.18 | 172.16 |
| 11590 (CL1) | 3.15 | 6.82 | 10.28 | 7.21 | 4.07 | 4.42 | 40.55 | 211.54 | 428.81 | 324.02 | 221.96 | 194.86 |
| 11600 | 4.51 | 2.20 | 5.98 | 7.97 | 3.56 | 3.90 | 88.97 | 81.91 | 423.48 | 560.70 | 192.33 | 234.28 |
| 11604 | 3.86 | 3.57 | 6.40 | 8.72 | 4.04 | 4.44 | 40.52 | 116.16 | 310.77 | 412.53 | 284.03 | 273.35 |
| 11619 | 4.35 | 5.42 | 7.95 | 6.85 | 4.11 | 3.67 | 54.52 | 159.59 | 312.05 | 476.69 | 116.96 | 173.91 |
| 11635 | 5.39 | 3.95 | 11.39 | 6.65 | 2.81 | 2.36 | 93.97 | 121.80 | 566.40 | 316.13 | 169.25 | 134.77 |
| 11693 | 7.91 | 2.16 | 5.62 | 7.63 | 4.27 | 2.67 | 107.97 | 87.74 | 294.41 | 480.59 | 107.05 | 117.37 |
| 11701 | 2.51 | 3.18 | 3.44 | 5.55 | 2.51 | 2.47 | 31.39 | 99.05 | 184.38 | 463.21 | 148.50 | 117.52 |
| N565 | 6.15 | 6.44 | 10.08 | 7.26 | 4.43 | 3.96 | 82.06 | 160.81 | 400.09 | 421.66 | 154.92 | 140.77 |
| A48118 | 6.20 | 3.28 | 10.26 | 5.84 | 4.98 | 6.23 | 76.50 | 88.12 | 602.09 | 314.62 | 162.54 | 226.70 |
| CAL-1 | 2.48 | 3.60 | 3.31 | 2.80 | 3.62 | 3.02 | 40.58 | 142.66 | 249.72 | 118.94 | 141.14 | 151.22 |
| CAL-2 | 4.14 | 2.17 | 3.30 | 3.03 | 3.97 | 0.92 | 83.24 | 87.36 | 180.70 | 192.28 | 286.13 | 55.74 |
| CAL-7 | 2.48 | 1.90 | 5.51 | 3.81 | 2.21 | 2.73 | 46.03 | 57.44 | 234.03 | 363.36 | 214.11 | 163.72 |
| AZ-1 | 7.37 | 6.14 | 6.05 | 6.34 | 4.19 | 6.02 | 91.80 | 281.22 | 261.83 | 322.44 | 198.77 | 240.25 |
| AZ-2 | 2.97 | 3.81 | 3.97 | 5.15 | 6.39 | 4.30 | 47.55 | 175.49 | 142.45 | 421.38 | 368.77 | 364.00 |
| AZ-3 | 3.98 | 1.91 | 6.48 | 3.96 | 1.54 | 2.97 | 82.36 | 93.43 | 359.86 | 258.31 | 97.24 | 149.14 |
| AZ-5 | 4.27 | 4.62 | 9.02 | 7.36 | 3.61 | 5.31 | 63.48 | 145.39 | 517.38 | 413.81 | 241.88 | 276.36 |
| AZ-6 | 4.31 | 7.42 | 8.05 | 7.51 | 2.61 | 3.42 | 67.94 | 237.15 | 276.29 | 327.24 | 96.56 | 150.41 |
| R1040 | 4.39 | 7.22 | 8.05 | 6.46 | 5.08 | 4.81 | 77.97 | 387.36 | 484.22 | 326.43 | 413.44 | 295.26 |
| R1092 | 2.99 | 4.62 | 3.03 | 4.36 | 3.86 | 3.41 | 24.89 | 74.50 | 106.33 | 249.44 | 102.29 | 122.36 |
| R1093 | 1.87 | 5.16 | 4.94 | 4.32 | 3.21 | 3.89 | 19.65 | 106.45 | 172.71 | 171.68 | 123.87 | 115.54 |
| R1100 | 3.29 | 2.85 | 6.53 | 3.98 | 2.95 | 3.28 | 49.08 | 83.65 | 186.02 | 188.58 | 132.54 | 119.33 |
| R1101 | 1.96 | 1.78 | 5.00 | 2.64 | 2.70 | 3.57 | 28.12 | 53.94 | 193.68 | 136.87 | 150.90 | 141.30 |
| R1103 | 3.95 | 1.96 | 1.84 | 3.20 | 5.88 | 2.41 | 51.83 | 61.00 | 94.54 | 162.35 | 254.71 | 119.07 |
| R1108 | 6.61 | 4.26 | 6.44 | 5.60 | 3.96 | 5.26 | 72.73 | 84.45 | 178.92 | 218.57 | 116.50 | 166.06 |
| mean | 4.17 | 4.21 | 6.56 | 5.74 | 4.70 | 3.80 | 60.16 | 134.60 | 303.59 | 341.02 | 233.85 | 174.42 |
| LSD | 0.35 | 0.40 | 0.54 | 0.41 | 0.32 | 0.31 | 6.15 | 20.10 | 52.74 | 37.13 | 41.56 | 22.81 |
| Sig. | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | * | *** |
| Accessions | Rs (%) | YRs (kg ha−1) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S1 | S2 | S3 | S4R | S5R | S6R | S1 | S2 | S3 | S4R | S5R | S6R | |
| CFS17-2005 | 9.78 | 10.08 | 7.00 | 9.41 | 7.67 | 6.95 | 116.21 | 182.63 | 368.34 | 425.91 | 148.17 | 190.48 |
| CFS18-2005 | 10.30 | 8.71 | 8.30 | 9.77 | 7.53 | 6.78 | 152.96 | 197.53 | 327.14 | 516.40 | 255.99 | 346.29 |
| 593 | 6.89 | 6.89 | 5.57 | 7.21 | 4.76 | 5.07 | 94.54 | 203.27 | 242.19 | 390.12 | 216.41 | 178.73 |
| 11590 (CL1) | 11.23 | 10.28 | 9.86 | 9.01 | 7.16 | 6.57 | 144.42 | 319.03 | 411.27 | 404.85 | 368.09 | 294.68 |
| 11600 | 11.14 | 14.82 | 16.90 | 12.78 | 10.77 | 5.39 | 220.00 | 552.83 | 1196.34 | 899.19 | 586.21 | 327.06 |
| 11604 | 11.66 | 16.00 | 9.07 | 12.50 | 8.58 | 7.41 | 122.43 | 520.76 | 440.34 | 591.23 | 558.65 | 452.57 |
| 11619 | 14.24 | 14.88 | 11.65 | 11.80 | 8.69 | 8.49 | 178.49 | 438.49 | 457.25 | 820.79 | 257.94 | 408.80 |
| 11635 | 11.08 | 14.69 | 11.41 | 13.74 | 8.00 | 7.31 | 192.94 | 452.78 | 567.22 | 652.73 | 501.17 | 413.97 |
| 11693 | 15.35 | 13.77 | 10.32 | 13.06 | 9.88 | 7.23 | 209.54 | 559.27 | 540.59 | 822.49 | 249.12 | 317.10 |
| 11701 | 14.10 | 11.33 | 13.01 | 13.03 | 7.35 | 7.86 | 176.44 | 352.45 | 697.81 | 1078.39 | 409.86 | 362.04 |
| N565 | 7.11 | 13.07 | 10.99 | 10.04 | 10.34 | 6.10 | 94.89 | 326.18 | 436.04 | 583.64 | 336.54 | 216.95 |
| A48118 | 14.55 | 13.95 | 12.24 | 11.99 | 8.17 | 9.34 | 179.65 | 374.93 | 718.19 | 645.57 | 260.68 | 340.25 |
| CAL-1 | 10.25 | 11.79 | 7.75 | 8.81 | 6.90 | 6.09 | 167.83 | 466.78 | 553.60 | 374.96 | 260.33 | 305.10 |
| CAL-2 | 11.43 | 7.70 | 8.43 | 9.57 | 8.46 | 3.99 | 229.93 | 309.57 | 465.13 | 607.05 | 645.85 | 241.56 |
| CAL-7 | 12.43 | 12.87 | 10.05 | 16.35 | 9.07 | 8.81 | 230.27 | 388.99 | 426.90 | 1559.26 | 745.03 | 528.18 |
| AZ-1 | 11.48 | 16.69 | 8.67 | 12.87 | 8.01 | 9.45 | 143.03 | 765.00 | 375.00 | 654.47 | 367.10 | 376.99 |
| AZ-2 | 9.73 | 12.92 | 10.87 | 14.36 | 8.67 | 9.80 | 155.76 | 598.42 | 389.94 | 1174.73 | 528.04 | 813.00 |
| AZ-3 | 14.17 | 13.00 | 11.23 | 15.31 | 9.89 | 8.85 | 292.97 | 636.78 | 623.30 | 997.42 | 608.58 | 446.86 |
| AZ-5 | 14.57 | 19.25 | 10.92 | 16.23 | 7.83 | 9.06 | 216.71 | 606.04 | 626.53 | 912.53 | 499.39 | 471.52 |
| AZ-6 | 8.70 | 12.50 | 7.52 | 9.51 | 6.67 | 5.88 | 137.17 | 399.66 | 258.11 | 414.25 | 238.13 | 258.49 |
| R1040 | 12.57 | 13.91 | 9.25 | 13.37 | 8.74 | 8.62 | 223.51 | 751.69 | 556.70 | 675.40 | 730.59 | 528.34 |
| R1092 | 9.57 | 9.46 | 12.79 | 12.49 | 7.88 | 8.36 | 79.75 | 152.52 | 449.14 | 715.33 | 224.47 | 299.50 |
| R1093 | 12.04 | 10.32 | 10.77 | 15.61 | 7.15 | 10.08 | 126.78 | 209.28 | 376.08 | 620.71 | 303.85 | 299.21 |
| R1100 | 13.17 | 10.36 | 8.15 | 9.11 | 5.37 | 5.71 | 196.58 | 304.34 | 231.95 | 431.88 | 263.35 | 207.95 |
| R1101 | 7.69 | 11.36 | 10.56 | 10.34 | 10.01 | 8.66 | 110.44 | 344.12 | 409.15 | 536.99 | 571.05 | 342.25 |
| R1103 | 11.42 | 9.84 | 6.55 | 10.86 | 7.08 | 6.90 | 149.70 | 305.69 | 336.73 | 551.28 | 306.04 | 340.17 |
| R1108 | 15.18 | 18.11 | 10.53 | 10.67 | 9.07 | 7.57 | 167.05 | 359.17 | 292.55 | 416.13 | 248.08 | 239.38 |
| mean | 11.55 | 12.54 | 10.01 | 11.84 | 10.38 | 7.48 | 167.48 | 421.85 | 479.12 | 720.21 | 504.00 | 348.93 |
| LSD | 0.54 | 0.68 | 0.52 | 0.60 | 0.44 | 0.38 | 15.08 | 55.86 | 77.93 | 82.03 | 82.64 | 42.85 |
| Sig. | *** | *** | *** | *** | *** | *** | *** | *** | ** | *** | NS | *** |
| Groups | Accessions | Sampling | ||||||
|---|---|---|---|---|---|---|---|---|
| Growth Cycle 1 | 24-Month-Old Plants | 12-Month-Old Plants | ||||||
| S1 | S2 | S3 | S3 | S4R | S1 | S6R | ||
| G1 | CFS17-2005 | c | bc | a | a | a | b | a |
| 11600 | b | b | a | b | a | b | a | |
| 11604 | c | bc | ab | b | a | b | a | |
| 11619 | c | bc | ab | b | a | b | a | |
| 11635 | b | b | a | a | b | a | a | |
| 11693 | c | c | b | b | a | a | a | |
| 11701 | b | b | b | b | a | ab | a | |
| N565 | b | b | a | ab | a | ab | a | |
| A48118 | d | d | a | a | b | b | a | |
| CAL-1 | b | b | a | a | b | b | a | |
| CAL-2 | cd | bcd | ab | a | a | a | ab | |
| AZ-3 | c | c | a | a | a | b | a | |
| AZ-5 | c | c | ab | a | a | b | a | |
| R1100 | c | c | a | a | a | b | a | |
| R1101 | c | c | a | a | b | b | a | |
| R1103 | c | c | c | b | a | b | a | |
| R1108 | b | b | a | ab | a | b | a | |
| CAL-7 | c | bc | a | a | a | b | a | |
| G2 | AZ-6 | c | ab | a | a | a | b | a |
| AZ-1 | c | ab | ab | b | a | b | a | |
| AZ-2 | b | ab | ab | b | a | b | a | |
| R1040 | b | ab | a | a | a | b | a | |
| R1092 | b | ab | ab | b | a | b | a | |
| R1093 | c | b | ab | b | a | b | a | |
| G3 | 11591 (CL1) | d | bc | a | a | ab | b | a |
| CFS18-2005 | c | bc | ab | ab | a | b | a | |
| 593 | d | c | b | b | a | b | a | |
| Accession | Species | Seed Code | Origin | Year | Ploidy |
|---|---|---|---|---|---|
| Traditional guayule pure accessions | |||||
| A48118 | P. argentatum | PI 478662 13i SO | Durango, México | 1948 | 3×, 4×, 4.5× |
| 593 | P. argentatum | PI 478639 13i SO | IRC | 1926 | 4× |
| 11600 | P. argentatum | PI 478641 15i SO | ERP, 4265-I | 1950 | 3×, 4× |
| 11604 | P. argentatum | PI 478642 12i SO | ERP, 4265-I | 1950 | 4× |
| 11619 | P. argentatum | PI 478645 12i SO | ERP, 4265-I | 1950 | 3×, 3.8×, 4× |
| 11635 | P. argentatum | PI 478648 15i SO | ERP, 4265-I | 1950 | 2×, 4× |
| 11693 | P. argentatum | PI 478650 12i SO | ERP, 4265-I | 1950 | 3.8×, 4× |
| 11701 | P. argentatum | PI 478651 12i SO | ERP, 4265-I | 1950 | 4× |
| N-565 | P. argentatum | PI 478655 15i SO | ERP, 4265-I | 1950 | 3×, 4×, 5.7× |
| Modern guayule pure accessions | |||||
| R-1040 | P. argentatum | W6 2192 15i SO | Coahuila, México | 1976 | 3.7×, 4× |
| R-1092 | P. argentatum | W6 2244 12i SO | Durango, México | 1979 | 3×, 4× |
| R-1093 | P. argentatum | W6 2245 15i SO | Durango, México | 1979 | 3×, 3.7× |
| R-1108 | P. argentatum | W6 2260 13i SO | Texas, USA | 1981 | 3×, 4× |
| CAL-7 | P. argentatum | W6 715715i SO | USDA-CA | 1985 | 4× |
| AZ-1 | P. argentatum | PI 599674 13i SO | USDA-AZ | 1997 | 4× |
| AZ-5 | P. argentatum | PI 599678 13i SO | USDA-AZ | 1997 | 4×, 4.4× |
| CFS17-2005 | P. argentatum | PARL 804 15i SO | Texas, USA | 2005 | 4× |
| CFS18-2005 | P. argentatum | PARL 805 15i SO | Texas, USA | 2005 | 3× (3), 4× (3) |
| 11591 (CL-1) | P. argentatum | CL1 | CIRAD | 2017 | 3× |
| AZ-6 | P. argentatum | CL6 | CIRAD | 2017 | 4× |
| Modern hybrid accessions | |||||
| R-1103 | P. argentatum × P. incanum | W6 2255 13i SO | Durango, México | 1977 | 4× |
| R-1100 | P. argentatum × P. incanum | W6 2252 13i SO | Coahuila, México | 1979 | 4× |
| R-1101 | P. argentatum × P. incanum | W6 2253 15i SO | Coahuila, México | 1979 | 4× |
| CAL-1 | P. argentatum × P. tomentosum | PI 478666 13i SO | USDA-CA | 1982 | 3×, 4.5×, 7× |
| CAL-2 | P. argentatum × fructicosum | PI 478667 13i S | USDA-CA | 1982 | 3×, 4× |
| AZ-2 | P. argentatum × unknown | PI 599675 13i SO | USDA-AZ | 1997 | 4× |
| AZ-3 | P. argentatum × unknown | PI 599676 12i SO | USDA-AZ | 1997 | 4×, 6× |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jara, F.M.; García-Martínez, M.d.l.M.; López-Córcoles, H.; Carrión, M.E.; Zalacain, A.; Carmona, M. Evaluating Guayule (Parthenium argentatum A. Gray) Germplasm Grown in Spain: Rubber and Resin along Three Production Cycles. Plants 2024, 13, 1092. https://doi.org/10.3390/plants13081092
Jara FM, García-Martínez MdlM, López-Córcoles H, Carrión ME, Zalacain A, Carmona M. Evaluating Guayule (Parthenium argentatum A. Gray) Germplasm Grown in Spain: Rubber and Resin along Three Production Cycles. Plants. 2024; 13(8):1092. https://doi.org/10.3390/plants13081092
Chicago/Turabian StyleJara, Francisco Miguel, María de las Mercedes García-Martínez, Horacio López-Córcoles, María Engracia Carrión, Amaya Zalacain, and Manuel Carmona. 2024. "Evaluating Guayule (Parthenium argentatum A. Gray) Germplasm Grown in Spain: Rubber and Resin along Three Production Cycles" Plants 13, no. 8: 1092. https://doi.org/10.3390/plants13081092
APA StyleJara, F. M., García-Martínez, M. d. l. M., López-Córcoles, H., Carrión, M. E., Zalacain, A., & Carmona, M. (2024). Evaluating Guayule (Parthenium argentatum A. Gray) Germplasm Grown in Spain: Rubber and Resin along Three Production Cycles. Plants, 13(8), 1092. https://doi.org/10.3390/plants13081092