
Halotolerant Endophytic Bacteria Regulate Growth and Field Performance of Two Durum Wheat Genotypes with Contrasting Salinity Tolerance Potential

 , and
, and 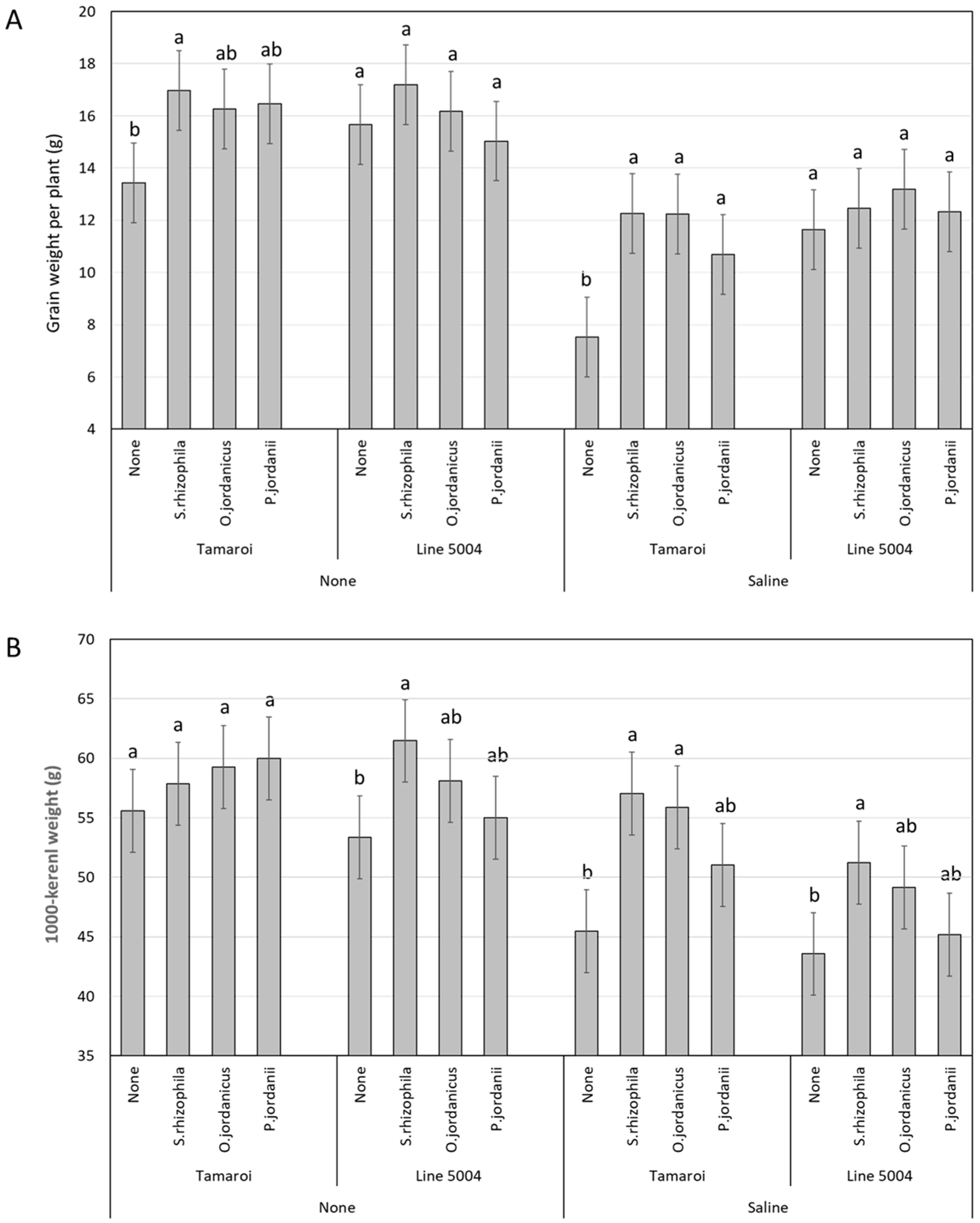{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
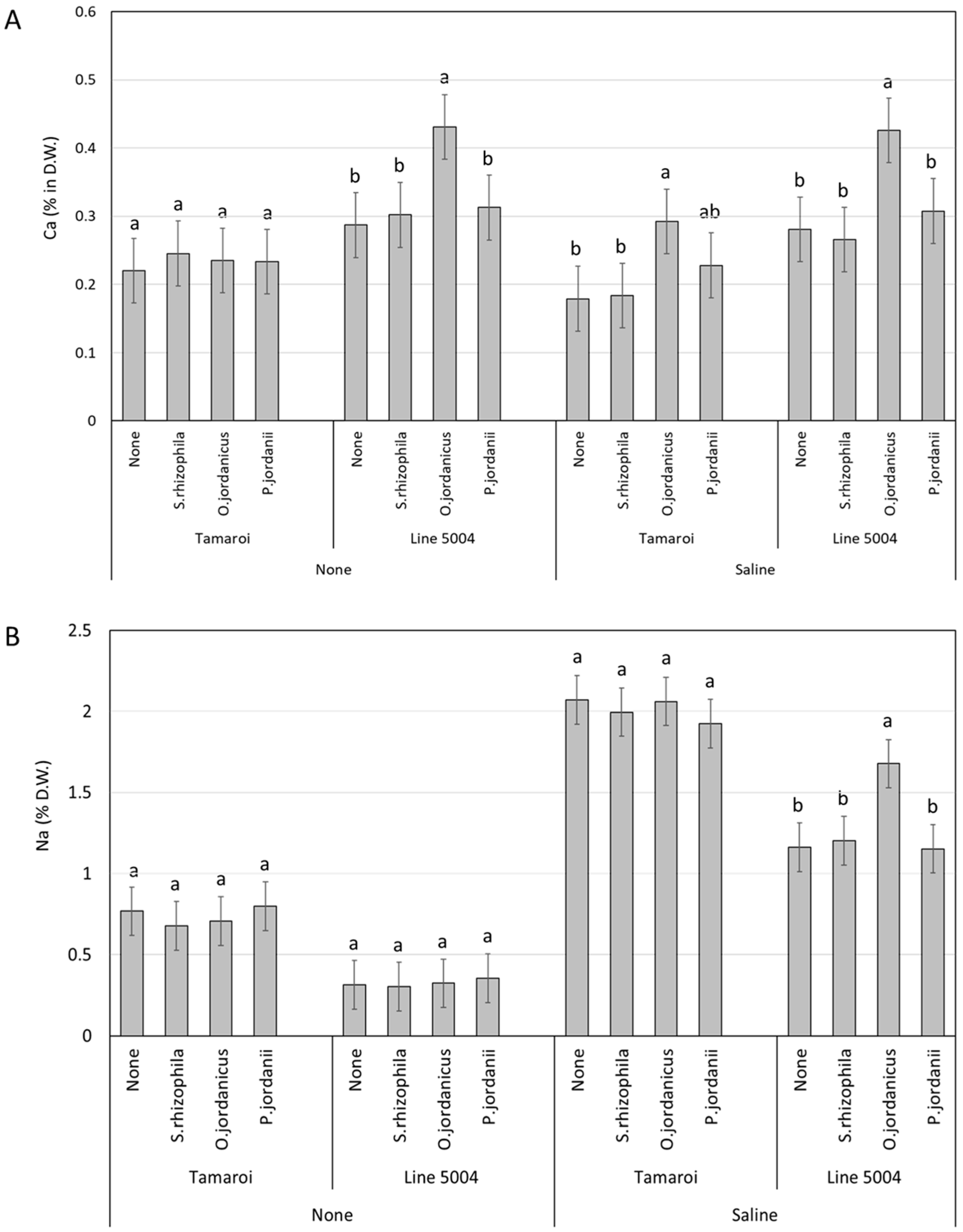
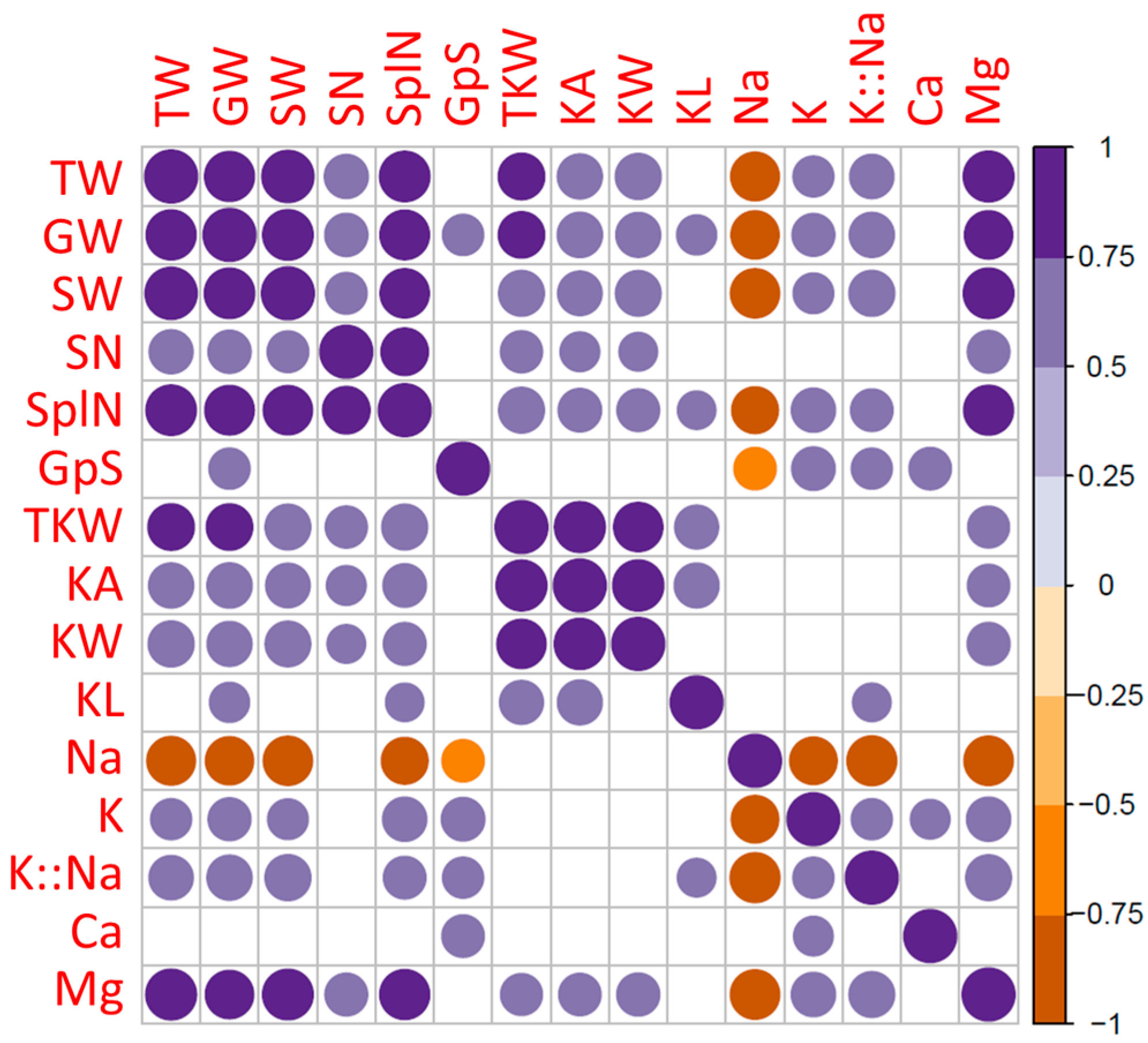
2. Results
2.1. Greenhouse Experiment
2.2. Field Experiment
3. Discussion
4. Materials and Methods
4.1. Bacteria Strains
4.2. Plant Material and Bacterial Inoculation
4.3. Greenhouse Experiment
4.4. Measurements and Data Analysis of Greenhouse Experiment
4.5. Field Experiment
4.6. Measurements and Data Analysis of Field Experiment
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rozema, J.; Flowers, T. Crops for a Salinized World. Science 2008, 322, 1478–1480. [Google Scholar] [CrossRef] [PubMed]
- Zörb, C.; Geilfus, C.M.; Dietz, K.J. Salinity and Crop Yield. Plant Biol. J. 2019, 21, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Etikala, B.; Adimalla, N.; Madhav, S.; Somagouni, S.G.; Keshava Kiran Kumar, P.L. Salinity problems in groundwater and management strategies in arid and semi-arid regions. In Groundwater Geochemistry: Pollution and Remediation; Madhav, S., Singhmethods, P., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2021; pp. 42–56. [Google Scholar] [CrossRef]
- Bazihizina, N.; Barrett-Lennard, E.G.; Colmer, T.D. Plant Growth and Physiology under Heterogeneous Salinity. Plant Soil 2012, 354, 1–19. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef]
- Munns, R.; Gilliham, M. Salinity Tolerance of Crops—What Is the Cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Saddiq, M.S.; Afzal, I.; Basra, S.M.A.; Iqbal, S.; Ashraf, M. Sodium Exclusion Affects Seed Yield and Physiological Traits of Wheat Genotypes Grown Under Salt Stress. J. Soil Sci. Plant Nutr. 2020, 20, 1442–1456. [Google Scholar] [CrossRef]
- Alom, R.; Hasan, M.A.; Islam, M.R.; Wang, Q.F. Germination Characters and Early Seedling Growth of Wheat (Triticum aestivum L.) Genotypes under Salt Stress Conditions. J. Crop Sci. Biotechnol. 2016, 19, 383–392. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Rahman, A.; Anee, T.I.; Alam, M.U.; Bhuiyan, T.F.; Oku, H.; Fujita, M. Approaches to Enhance Salt Stress Tolerance in Wheat. In Wheat Improvement, Management and Utilization; Wanyera, R., Owuoche, J., Eds.; InTech: Cambridge, ON, USA, 2017; pp. 151–187. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Mehrvar, M.R.; Sanjani, S.; Amini, A.; Nikkhah-Chamanabad, H.; Asadi, A. Effects of Salinity Stress on Seedling Biomass, Physiochemical Properties, and Grain Yield in Different Breeding Wheat Genotypes. Acta Physiol. Plant. 2021, 43, 98. [Google Scholar] [CrossRef]
- Saddiq, M.S.; Iqbal, S.; Hafeez, M.B.; Ibrahim, A.M.H.; Raza, A.; Fatima, E.M.; Baloch, H.; Jahanzaib; Woodrow, P.; Ciarmiello, L.F. Effect of Salinity Stress on Physiological Changes in Winter and Spring Wheat. Agronomy 2021, 11, 1193. [Google Scholar] [CrossRef]
- Uzair, M.; Ali, M.; Fiaz, S.; Attia, K.; Khan, N.; Al-Doss, A.A.; Ramzan Khan, M.; Ali, Z. The Characterization of Wheat Genotypes for Salinity Tolerance Using Morpho-Physiological Indices under Hydroponic Conditions. Saudi J. Biol. Sci. 2022, 29, 103299. [Google Scholar] [CrossRef]
- Borrelli, G.; Ficco, D.; Giuzio, L.; Pompa, M.; Cattivelli, L.; Flagella, Z. Durum Wheat Salt Tolerance in Relation to Physiological, Yield and Quality Characters. Cereal Res. Commun. 2011, 39, 525–534. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Läuchli, A. Approaches to Increasing the Salt Tolerance of Wheat and Other Cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Shabala, L.; Azzarello, E.; Huang, Y.; Pandolfi, C.; Su, N.; Wu, Q.; Cai, S.; Bazihizina, N.; Wang, L.; et al. Na+ Extrusion from the Cytosol and Tissue-Specific Na+ Sequestration in Roots Confer Differential Salt Stress Tolerance between Durum and Bread Wheat. J. Exp. Bot. 2018, 69, 3987–4001. [Google Scholar] [CrossRef] [PubMed]
- James, R.A.; Blake, C.; Byrt, C.S.; Munns, R. Major Genes for Na+ Exclusion, Nax1 and Nax2 (Wheat HKT1;4 and HKT1;5), Decrease Na+ Accumulation in Bread Wheat Leaves under Saline and Waterlogged Conditions. J. Exp. Bot. 2011, 62, 2939–2947. [Google Scholar] [CrossRef]
- James, R.A.; Blake, C.; Zwart, A.B.; Hare, R.A.; Rathjen, A.J.; Munns, R. Impact of Ancestral Wheat Sodium Exclusion Genes Nax1 and Nax2 on Grain Yield of Durum Wheat on Saline Soils. Funct. Plant Biol. 2012, 39, 609. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; James, R.A.; Xu, B.; Athman, A.; Conn, S.J.; Jordans, C.; Byrt, C.S.; Hare, R.A.; Tyerman, S.D.; Tester, M.; et al. Wheat Grain Yield on Saline Soils Is Improved by an Ancestral Na+ Transporter Gene. Nat. Biotechnol. 2012, 30, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Mujeeb-Kazi, A.; Munns, R.; Rasheed, A.; Ogbonnaya, F.C.; Ali, N.; Hollington, P.; Dundas, I.; Saeed, N.; Wang, R.; Rengasamy, P.; et al. Breeding Strategies for Structuring Salinity Tolerance in Wheat. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2019; Volume 155, pp. 121–187. [Google Scholar] [CrossRef]
- Majeed, A.; Muhammad, Z. Salinity: A Major Agricultural Problem—Causes, Impacts on Crop Productivity and Management Strategies. In Plant Abiotic Stress Tolerance; Hasanuzzaman, M., Hakeem, K.R., Nahar, K., Alharby, H.F., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 83–99. [Google Scholar]
- Huang, M.; Zhang, Z.; Zhai, Y.; Lu, P.; Zhu, C. Effect of Straw Biochar on Soil Properties and Wheat Production under Saline Water Irrigation. Agronomy 2019, 9, 457. [Google Scholar] [CrossRef]
- Mousavi, H.; Bruning, B.; Van Straten, G.; Almås, Å.R.; Lethin, J.; Naik, N.; Hassan, S.; Olsson, O.; Aronsson, H. Effects of Increasing Salinity by Drip Irrigation on Total Grain Weight Show High Yield Potential of Putative Salt-Tolerant Mutagenized Wheat Lines. Sustainability 2022, 14, 5061. [Google Scholar] [CrossRef]
- Etesami, H.; Maheshwari, D.K. Use of Plant Growth Promoting Rhizobacteria (PGPRs) with Multiple Plant Growth Promoting Traits in Stress Agriculture: Action Mechanisms and Future Prospects. Ecotoxicol. Environ. Saf. 2018, 156, 225–246. [Google Scholar] [CrossRef]
- Adeleke, B.S.; Babalola, O.O.; Glick, B.R. Plant Growth-Promoting Root-Colonizing Bacterial Endophytes. Rhizosphere 2021, 20, 100433. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, P.; Pahal, V.; Kumar, J.; Pandey, S.S. Endophytic Phytohormone Production and Utilization of Functional Traits in Plant Growth Promotion. In Plant Microbiome for Plant Productivity and Sustainable Agriculture; Chhabra, S., Prasad, R., Maddela, N.R., Tuteja, N., Eds.; Microorganisms for Sustainability; Springer Nature: Singapore, 2023; Volume 37, pp. 365–385. [Google Scholar] [CrossRef]
- Win, K.T.; Tanaka, F.; Okazaki, K.; Ohwaki, Y. The ACC Deaminase Expressing Endophyte Pseudomonas spp. Enhances NaCl Stress Tolerance by Reducing Stress-Related Ethylene Production, Resulting in Improved Growth, Photosynthetic Performance, and Ionic Balance in Tomato Plants. Plant Physiol. Biochem. 2018, 127, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Afridi, M.S.; Mahmood, T.; Salam, A.; Mukhtar, T.; Mehmood, S.; Ali, J.; Khatoon, Z.; Bibi, M.; Javed, M.T.; Sultan, T.; et al. Induction of Tolerance to Salinity in Wheat Genotypes by Plant Growth Promoting Endophytes: Involvement of ACC Deaminase and Antioxidant Enzymes. Plant Physiol. Biochem. 2019, 139, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Alhindi, T.; Albdaiwi, R. Draft Genome Sequence of Oceanobacillus jordanicus Strain GSFE11, a Halotolerant Plant Growth-Promoting Bacterial Endophyte Isolated from the Jordan Valley. Evol. Bioinform. 2022, 18, 117693432110711. [Google Scholar] [CrossRef]
- Albdaiwi, R.N.; Khyami-Horani, H.; Ayad, J.Y.; Alananbeh, K.M.; Al-Sayaydeh, R. Isolation and Characterization of Halotolerant Plant Growth Promoting Rhizobacteria from Durum Wheat (Triticum turgidum subsp. durum) Cultivated in Saline Areas of the Dead Sea Region. Front. Microbiol. 2019, 10, 1639. [Google Scholar] [CrossRef] [PubMed]
- Al-Sayaydeh, R.; Shtaya, M.J.; Qubbaj, T.; Al-Rifaee, M.K.; Alabdallah, M.A.; Migdadi, O.; Gammoh, I.A.; Al-Abdallat, A.M. Performance and Stability Analysis of Selected Durum Wheat Genotypes Differing in Their Kernel Characteristics. Plants 2023, 12, 2664. [Google Scholar] [CrossRef] [PubMed]
- Pastuszak, J.; Dziurka, M.; Hornyák, M.; Szczerba, A.; Kopeć, P.; Płażek, A. Physiological and Biochemical Parameters of Salinity Resistance of Three Durum Wheat Genotypes. Int. J. Mol. Sci. 2022, 23, 8397. [Google Scholar] [CrossRef] [PubMed]
- Kerbab, S.; Silini, A.; Chenari Bouket, A.; Cherif-Silini, H.; Eshelli, M.; El Houda Rabhi, N.; Belbahri, L. Mitigation of NaCl Stress in Wheat by Rhizosphere Engineering Using Salt Habitat Adapted PGPR Halotolerant Bacteria. Appl. Sci. 2021, 11, 1034. [Google Scholar] [CrossRef]
- Zahir, Z.A.; Ghani, U.; Naveed, M.; Nadeem, S.M.; Asghar, H.N. Comparative Effectiveness of Pseudomonas and Serratia sp. Containing ACC-Deaminase for Improving Growth and Yield of Wheat (Triticum aestivum L.) under Salt-Stressed Conditions. Arch. Microbiol. 2009, 191, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Soltani, J.; Samavati, R.; Jalili, B.; Bagheri, H.; Hamzei, J. Halotolerant Endophytic Bacteria from Desert-Adapted Halophyte Plants Alleviate Salinity Stress in Germinating Seeds of the Common Wheat Triticum aestivum L. Cereal Res. Commun. 2024, 52, 165–175. [Google Scholar] [CrossRef]
- Mahmoud, O.M.B.; Slimene, I.B.; Zribi, O.T.; Abdelly, C.; Djébali, N. Response to Salt Stress Is Modulated by Growth-Promoting Rhizobacteria Inoculation in Two Contrasting Barley Cultivars. Acta Physiol. Plant 2017, 39, 120. [Google Scholar] [CrossRef]
- Amini Hajiabadi, A.; Mosleh Arani, A.; Etesami, H. Salt-Tolerant Genotypes and Halotolerant Rhizobacteria: A Potential Synergistic Alliance to Endure High Salinity Conditions in Wheat. Environ. Exp. Bot. 2022, 202, 105033. [Google Scholar] [CrossRef]
- Razzaghi Komaresofla, B.; Alikhani, H.A.; Etesami, H.; Khoshkholgh-Sima, N.A. Improved Growth and Salinity Tolerance of the Halophyte Salicornia sp. by Co–Inoculation with Endophytic and Rhizosphere Bacteria. Appl. Soil Ecol. 2019, 138, 160–170. [Google Scholar] [CrossRef]
- Abd_Allah, E.F.; Alqarawi, A.A.; Hashem, A.; Radhakrishnan, R.; Al-Huqail, A.A.; Al-Otibi, F.O.N.; Malik, J.A.; Alharbi, R.I.; Egamberdieva, D. Endophytic Bacterium Bacillus subtilis (BERA 71) Improves Salt Tolerance in Chickpea Plants by Regulating the Plant Defense Mechanisms. J. Plant Interact. 2018, 13, 37–44. [Google Scholar] [CrossRef]
- Davenport, R.J.; Reid, R.J.; Smith, F.A. Sodium-Calcium Interactions in Two Wheat Species Differing in Salinity Tolerance. Physiol. Plant. 1997, 99, 323–327. [Google Scholar] [CrossRef]
- Seifikalhor, M.; Aliniaeifard, S.; Shomali, A.; Azad, N.; Hassani, B.; Lastochkina, O.; Li, T. Calcium Signaling and Salt Tolerance are Diversely Entwined in Plants. Plant Signal. Behav. 2019, 14, 1665455. [Google Scholar] [CrossRef] [PubMed]
- Al-Whaibi, M.H.; Siddiqui, M.H.; Basalah, M.O. Salicylic Acid and Calcium-Induced Protection of Wheat against Salinity. Protoplasma 2012, 249, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Egamberdieva, D.; Lugtenberg, B.; Hagemann, M. Symbiotic Plant–Microbe Interactions: Stress Protection, Plant Growth Promotion, and Biocontrol by Stenotrophomonas. In Symbioses and Stress; Seckbach, J., Grube, M., Eds.; Cellular Origin, Life in Extreme Habitats and Astrobiology; Springer: Dordrecht, The Netherlands, 2010; Volume 17, pp. 445–460. [Google Scholar] [CrossRef]
- James, R.A.; Davenport, R.J.; Munns, R. Physiological Characterization of Two Genes for Na+ Exclusion in Durum Wheat, Nax1 and Nax2. Plant Physiol. 2006, 142, 1537–1547. [Google Scholar] [CrossRef] [PubMed]
- Albdaiwi, R.N.; Al-Hawadi, J.S.; Al-Rawashdeh, Z.B.; Al-Habahbeh, K.A.; Ayad, J.Y.; Al-Sayaydeh, R.S. Effect of Treated Wastewater Irrigation on the Accumulation and Transfer of Heavy Metals in Lemon Trees Cultivated in Arid Environment. Horticulturae 2022, 8, 514. [Google Scholar] [CrossRef]
- Nawaz, A.; Shahbaz, M.; Asadullah; Imran, A.; Marghoob, M.U.; Imtiaz, M.; Mubeen, F. Potential of salt tolerant PGPR in growth and yield augmentation of wheat (Triticum aestivum L.) under saline conditions. Front. Microbiol. 2020, 11, 2019. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albdaiwi, R.; Al-Sayaydeh, R.; Al-Rifaee, M.K.; Alhindi, T.; Ashraf, M.; Al-Abdallat, A.M. Halotolerant Endophytic Bacteria Regulate Growth and Field Performance of Two Durum Wheat Genotypes with Contrasting Salinity Tolerance Potential. Plants 2024, 13, 1179. https://doi.org/10.3390/plants13091179
Albdaiwi R, Al-Sayaydeh R, Al-Rifaee MK, Alhindi T, Ashraf M, Al-Abdallat AM. Halotolerant Endophytic Bacteria Regulate Growth and Field Performance of Two Durum Wheat Genotypes with Contrasting Salinity Tolerance Potential. Plants. 2024; 13(9):1179. https://doi.org/10.3390/plants13091179
Chicago/Turabian StyleAlbdaiwi, Randa, Rabea Al-Sayaydeh, Mohammad K. Al-Rifaee, Tareq Alhindi, Muhammad Ashraf, and Ayed M. Al-Abdallat. 2024. "Halotolerant Endophytic Bacteria Regulate Growth and Field Performance of Two Durum Wheat Genotypes with Contrasting Salinity Tolerance Potential" Plants 13, no. 9: 1179. https://doi.org/10.3390/plants13091179
APA StyleAlbdaiwi, R., Al-Sayaydeh, R., Al-Rifaee, M. K., Alhindi, T., Ashraf, M., & Al-Abdallat, A. M. (2024). Halotolerant Endophytic Bacteria Regulate Growth and Field Performance of Two Durum Wheat Genotypes with Contrasting Salinity Tolerance Potential. Plants, 13(9), 1179. https://doi.org/10.3390/plants13091179