
An Improved Syringe Agroinfiltration Protocol to Enhance Transformation Efficiency by Combinative Use of 5-Azacytidine, Ascorbate Acid and Tween-20

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
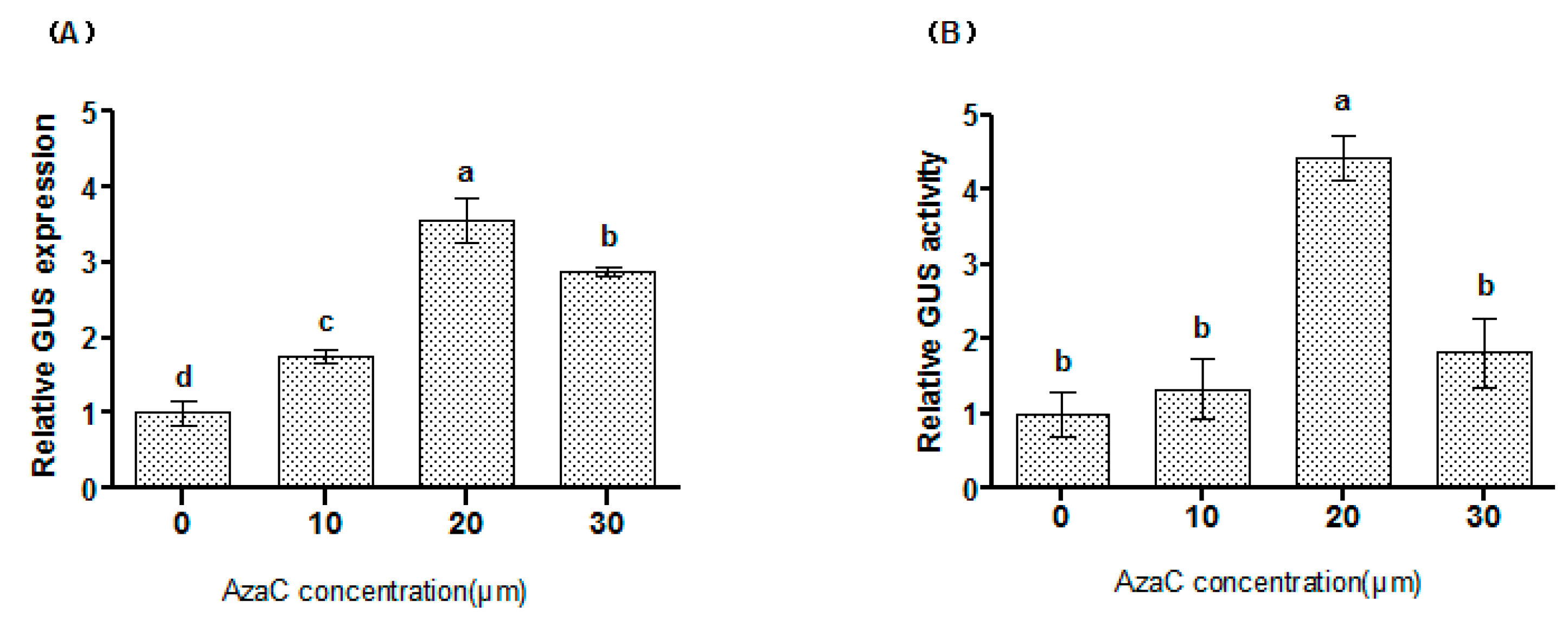
2.1. The Effects of AzaC on Transformation Efficiency
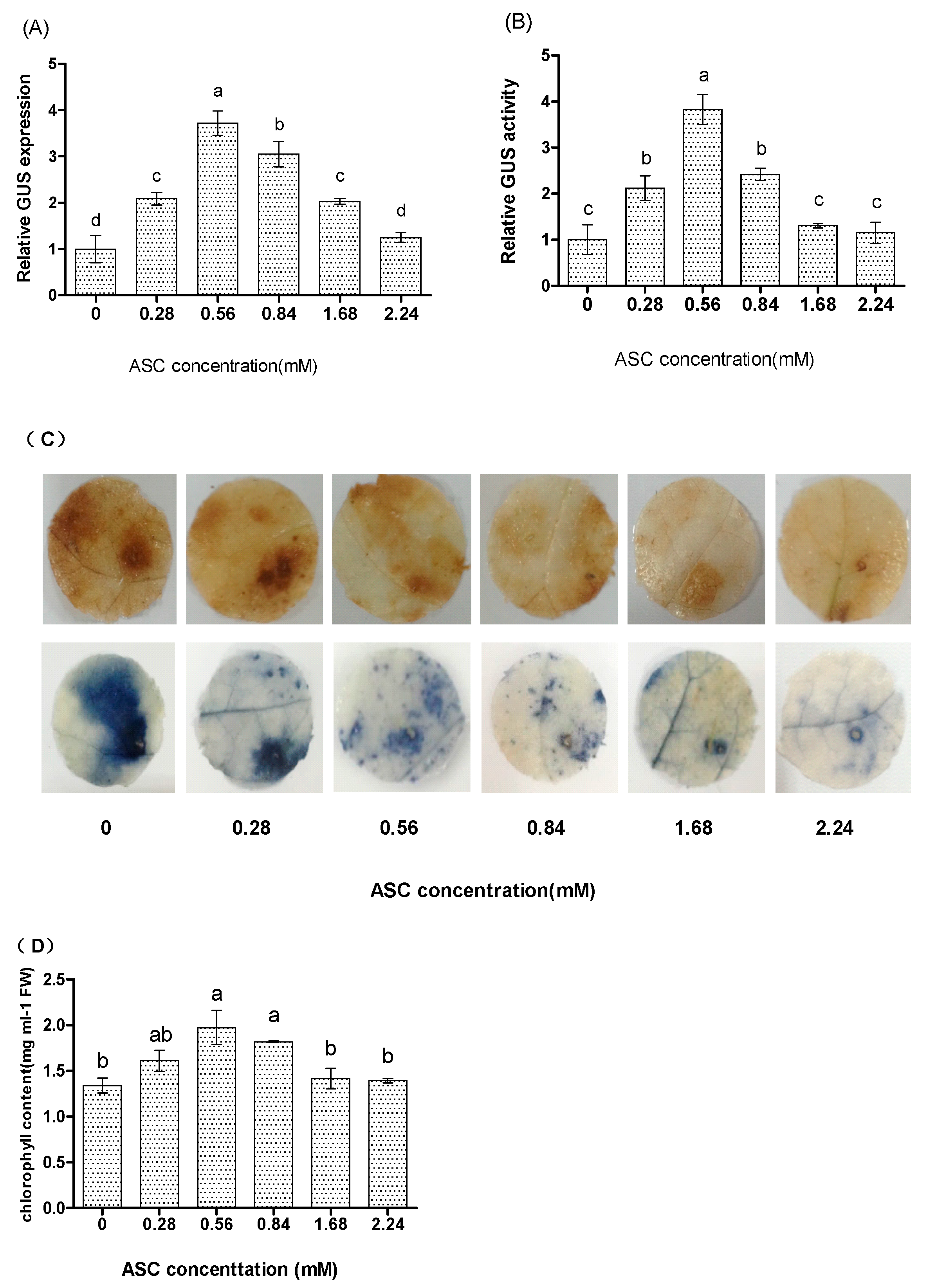
2.2. ASC Significantly Affects the Expression of Transgene
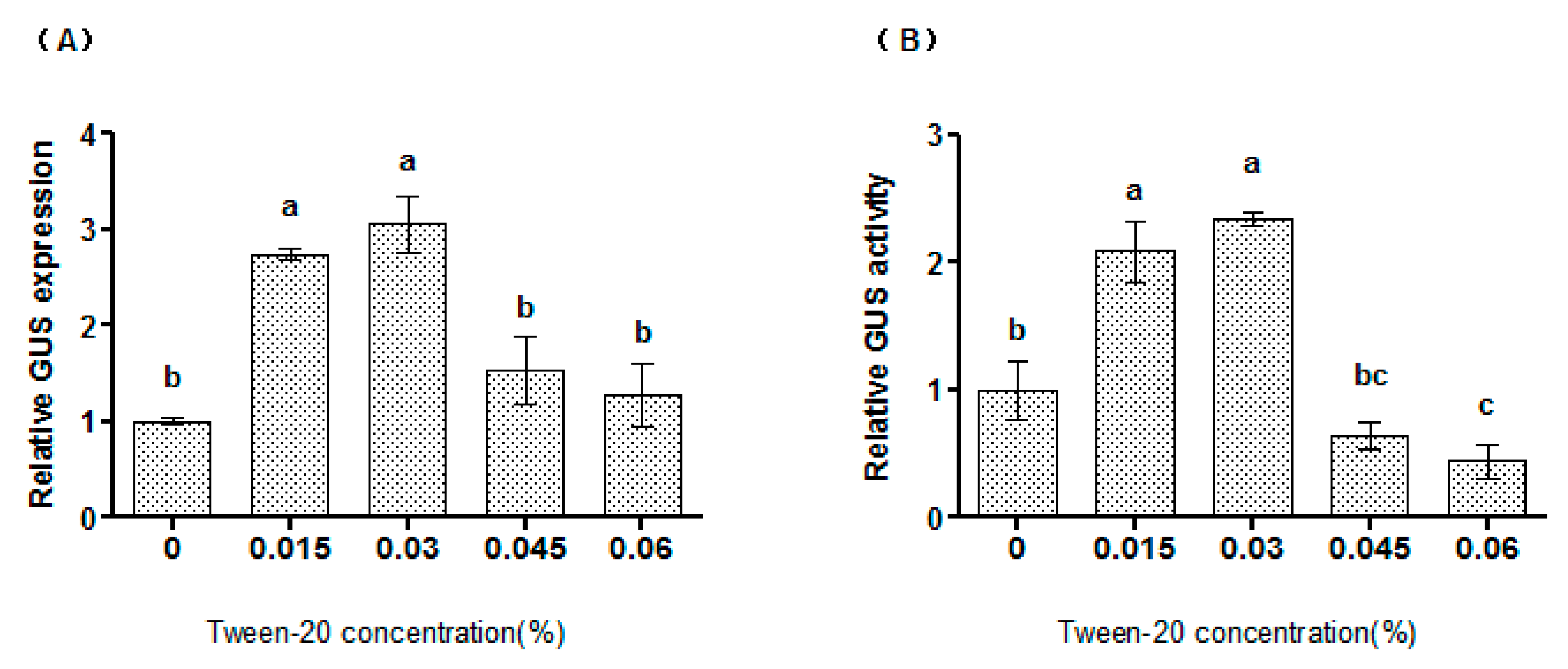
2.3. Tween-20 Could Increase the Transformation Efficiency of Syringe Agroinfiltration
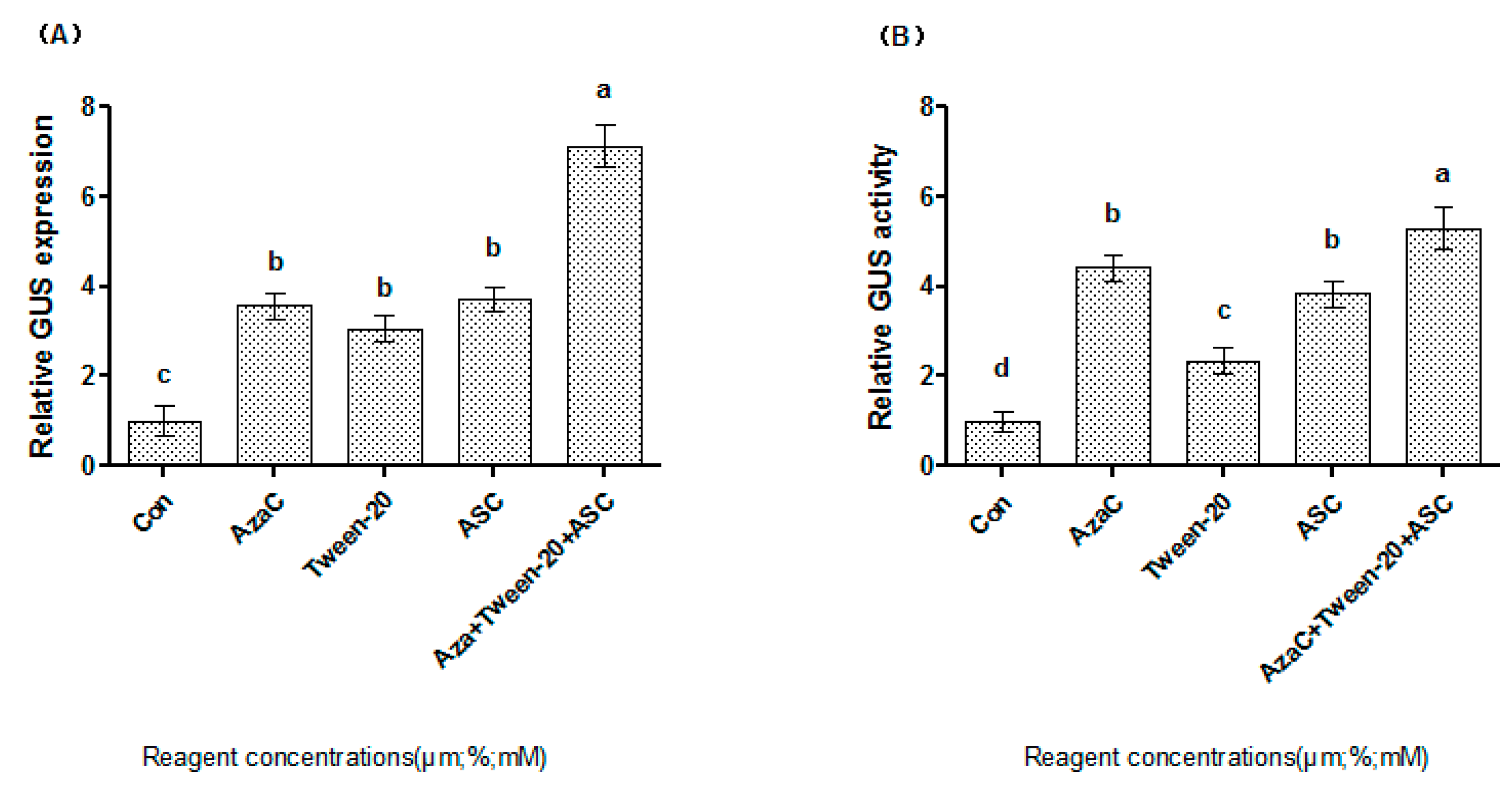
2.4. A Combination of AzaC, ASC and Tween-20 Highly Improves Transformation Efficiency
3. Materials and Methods
3.1. Plant Materials and Growth Conditions
3.2. Infiltration Procedures
3.3. Factors Influencing Syringe Infiltration
3.4. Determination of β-Glucuronidase (GUS) Activity
3.5. Quantitative Reverse Transcription PCR (qRT-PCR)
3.6. DAB and NBT Staining and Chlorophyll Content Assay
3.7. Statistical Analyses
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Guidarelli, M.; Baraldi, E. Transient transformation meets gene function discovery: The strawberry fruit case. Front. Plant Sci. 2015, 6, 444. [Google Scholar] [CrossRef] [PubMed]
- Wroblewski, T.; Tomczak, A.; Michelmore, R. Optimization of Agrobacterium-mediated transient assays of gene expression in lettuce, tomato and Arabidopsis. Plant Biotech. J. 2005, 3, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Santi, L.; Batchelor, L.; Huang, Z.; Hjelm, B.; Kilbourne, J.; Arntzen, C.J.; Chen, Q.; Mason, H.S. An efficient plant viral expression system generating orally immunogenic Norwalk virus-like particles. Vaccine 2008, 26, 1846–1854. [Google Scholar] [CrossRef] [PubMed]
- Vaghchhipawala, Z.; Rojas, C.M.; Senthil-Kumar, M.; Mysore, K.S. Agroinoculation and agroinfiltration: Simple tools for complex gene function analyses. Methods Mol. Biol. 2011, 678, 65–76. [Google Scholar] [PubMed]
- Chen, Q.; Lai, H.; Hurtado, J.; Stahnke, J.; Leuzinger, K.; Dent, M. Agroinfiltration as an Effective and Scalable Strategy of Gene Delivery for Production of Pharmaceutical Proteins. Adv. Tech. Biol. Med. 2013. [Google Scholar] [CrossRef]
- Dugdale, B.; Mortimer, C.L.; Kato, M.; James, T.; Harding, R.M.; Dale, J.L. Design and construction of an in-plant activation cassette for transgene expression and recombinant protein production in plants. Nat. Protoc. 2014, 9, 1010–1027. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, R.; Qi, M. In vivo analysis of plant promoters and transcription factors by agroinfiltration of tobacco leaves. Plant J. 2000, 22, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Johansen, L.K.; Carrington, J.C. Silencing on the spot Induction and suppression of RNA silencing in the Agrobacteriummediated transient expression system. Plant Physiol. 2001, 126, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Takeda, A.; Sugiyama, K.; Nagano, H.; Mori, M.; Kaido, M.; Mise, K.; Tsuda, S.; Okuno, T. Identification of a novel RNA silencing suppressor, NSs protein of Tomato spotted wilt virus. FEBS Lett. 2002, 532, 75–79. [Google Scholar] [CrossRef]
- Fujiuchi, N.; Matsuda, R.; Matoba, N.; Fujiwara, K. Removal of bacterial suspension water occupying the intercellular space of detached leaves after agroinfiltration improves the yield of recombinant hemagglutinin in a N benthamiana transient gene expression system. Biotechnol. Bioeng. 2016, 113, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Bochardt, A.; Hodal, L.; Palmgren, G.; Mattsson, O.; Okkels, F.T. DNA methylation is involved in maintenance of an unusual expression pattern of an introduced gene. Plant Physiol. 1992, 99, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Palmgren, G.; Mattson, O.; Okkels, F.T. Treatment of Agrobacterium or leaf disks with 5-azacytidine increases transgene expression in tobacco. Plant Mol. Biol. 1993, 21, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Wing, V.; Xie, T.; Shi, X.; Wang, Y.P.; Sokolov, V. DNA methylation analysis during the optimization of agrobacterium-mediated transformation of soybean. Genetika 2016, 52, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Z.; Karen, W.H.; Leaf, H. Effects of 5-azacytidine on transformation and gene expression in Nicotiana tabacum. Cell. Dev. Biol. Plant 1991, 27, 77–83. [Google Scholar]
- Christman, J.K. 5-Azacytidine and 5-aza-2′-deoxycytidine as inhibitors of DNA methylation: Mechanistic studies and their implications for cancer therapy. Oncogene 2002, 21, 5483–5495. [Google Scholar] [CrossRef] [PubMed]
- Weber, H.; Ziechmann, C.; Graessmann, A. In vitro DNA methylation inhibits gene expression in transgenic tobacco. EMBO J. 1990, 9, 4409–4415. [Google Scholar] [PubMed]
- Wojtaszek, P. Oxidative burst: An early plant response to pathogen infection. Biochem. J. 1997, 322, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Dan, Y. Biological functions of antioxidants in plant transformation. Cell. Dev. Biol. Plant 2008, 44, 149–161. [Google Scholar] [CrossRef]
- Kuta, D.D.; Tripathi, L. Agrobacterium-mduced hypersensitive necrotic reaction in plant cells: A resistance response against Agrobacterium-mediated DNA transfer. Afr. J. Biotechnol. 2005, 4, 752–757. [Google Scholar]
- Ji, X.; Zheng, L.; Liu, Y.; Nie, X.; Liu, S.; Wang, Y. A Transient Transformation System for the Functional Characterization of Genes Involved in Stress Response. Plant Mol. Biol. Rep. 2014, 3, 732–739. [Google Scholar] [CrossRef]
- Zang, D.; Wang, C.; Ji, X.; Wang, Y. Tamarix hispida zinc finger protein ThZFP1 participates in salt and osmotic stress tolerance by increasing proline content and SOD and POD activities. Plant Sci. 2015, 235, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Tague, B.W.; Mantis, J. In planta Agrobacterium-mediated transformation by vacuum infiltration. Methods Mol. Biol. 2006, 323, 215–223. [Google Scholar] [PubMed]
- Gous, P.W.; Gilbert, R.G.; Fox, G.P. Drought-proofing barley (Hordeum vulgare) and its impact on grain quality: A review. J. Inst. Brew. 2015, 121, 19–27. [Google Scholar] [CrossRef]
- Broghammer, A.; Krusell, L.; Blaise, M.; Sauer, J.; Sullivan, J.T.; Maolanon, N.; Vinther, M.; Lorentzen, A.; Madsen, E.B.; Jensen, K.J.; et al. Legume receptors perceive the rhizobial lipochitin oligosaccharide signal molecules by direct binding. Proc. Natl. Acad. Sci. USA 2012, 109, 13859–13864. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: β-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Fryer, M.J.; Oxborough, K.; Mullineaux, P.M.; Baker, N.R. Imaging of photo-oxidative stress responses in leaves. J. Exp. Bot. 2002, 53, 1249–1254. [Google Scholar] [CrossRef] [PubMed]




© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, H.; Tan, Z.; Wen, X.; Wang, Y. An Improved Syringe Agroinfiltration Protocol to Enhance Transformation Efficiency by Combinative Use of 5-Azacytidine, Ascorbate Acid and Tween-20. Plants 2017, 6, 9. https://doi.org/10.3390/plants6010009
Zhao H, Tan Z, Wen X, Wang Y. An Improved Syringe Agroinfiltration Protocol to Enhance Transformation Efficiency by Combinative Use of 5-Azacytidine, Ascorbate Acid and Tween-20. Plants. 2017; 6(1):9. https://doi.org/10.3390/plants6010009
Chicago/Turabian StyleZhao, Huimin, Zilong Tan, Xuejing Wen, and Yucheng Wang. 2017. "An Improved Syringe Agroinfiltration Protocol to Enhance Transformation Efficiency by Combinative Use of 5-Azacytidine, Ascorbate Acid and Tween-20" Plants 6, no. 1: 9. https://doi.org/10.3390/plants6010009
APA StyleZhao, H., Tan, Z., Wen, X., & Wang, Y. (2017). An Improved Syringe Agroinfiltration Protocol to Enhance Transformation Efficiency by Combinative Use of 5-Azacytidine, Ascorbate Acid and Tween-20. Plants, 6(1), 9. https://doi.org/10.3390/plants6010009