Impact of Elemental Sulfur on the Rhizospheric Bacteria of Durum Wheat Crop Cultivated on a Calcareous Soil



 and
and
Abstract
:1. Introduction
2. Materials and Methods
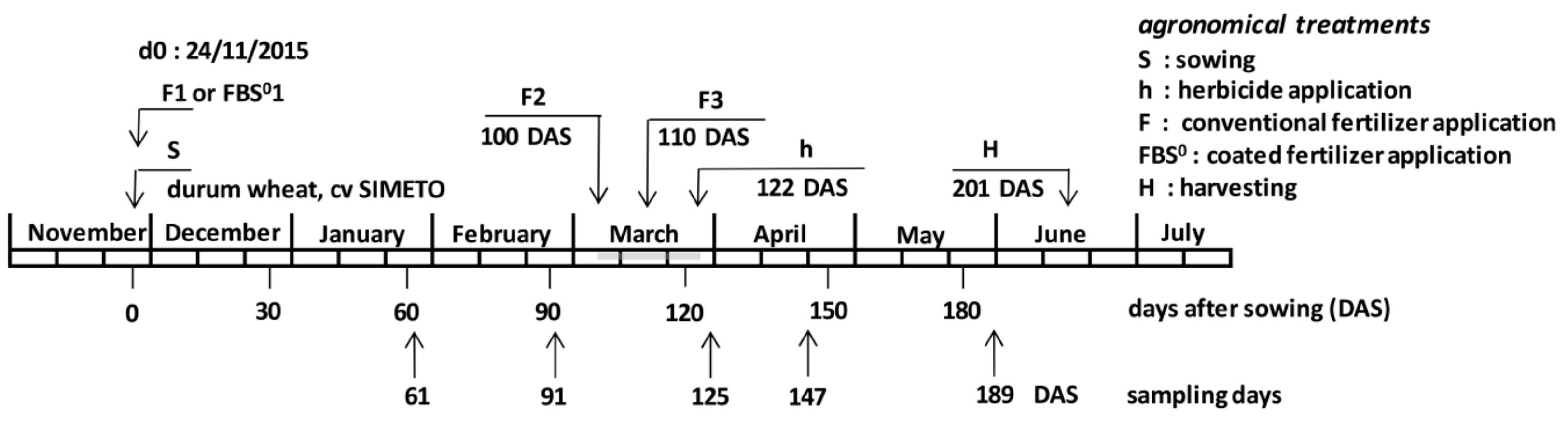
2.1. Experimental Setup
2.2. Determinations of Soil Parameters
2.3. Determinations of Dry Matter, P, Fe, Organic N, and Organic S Concentrations in the Aerial Part of the Plant
2.4. Microorganism Sampling, Isolation, Culture Conditions, and Arylsulfatase Activity
2.5. Assessment of Plant Growth-Promoting (PGP) Traits
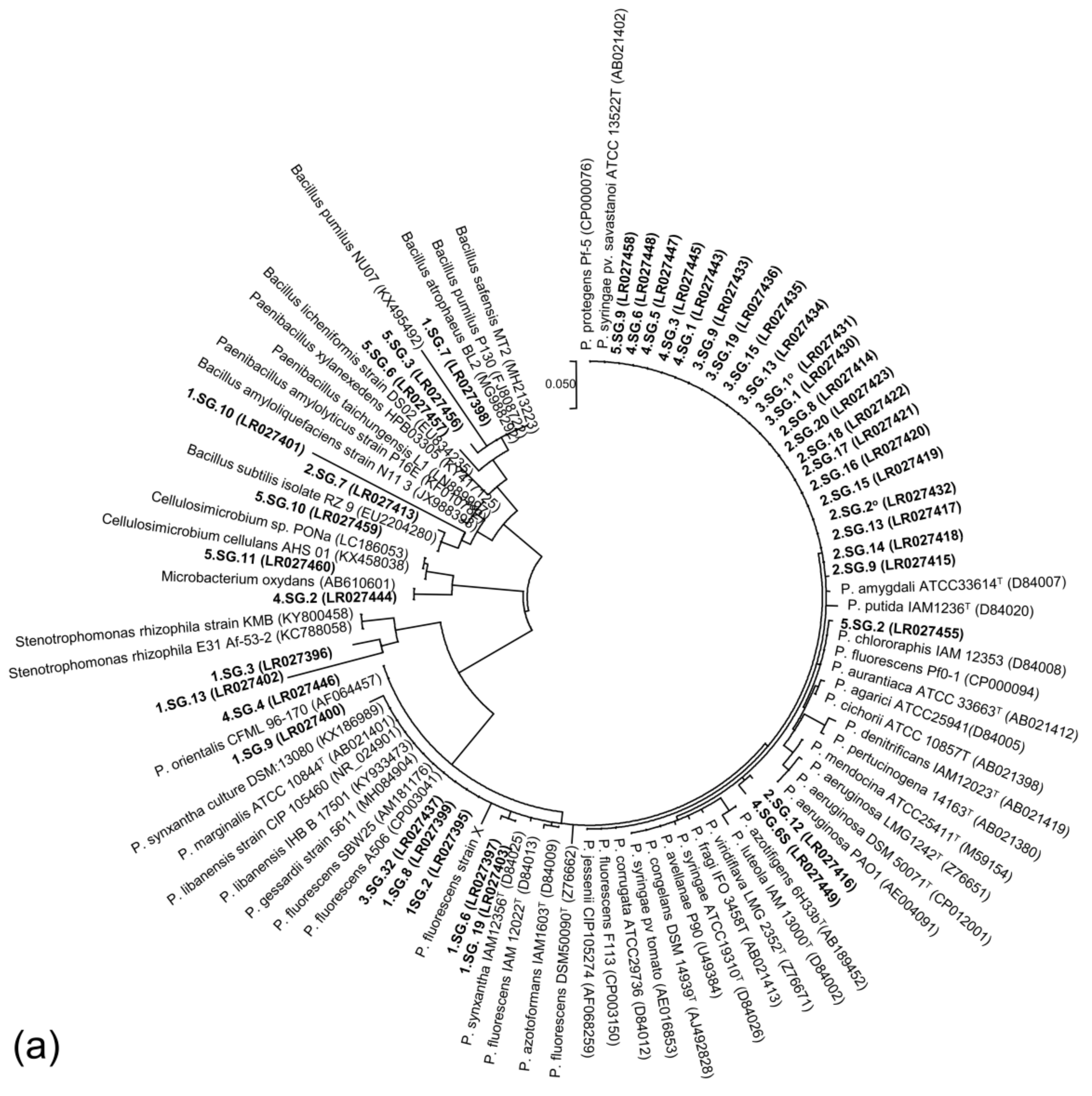
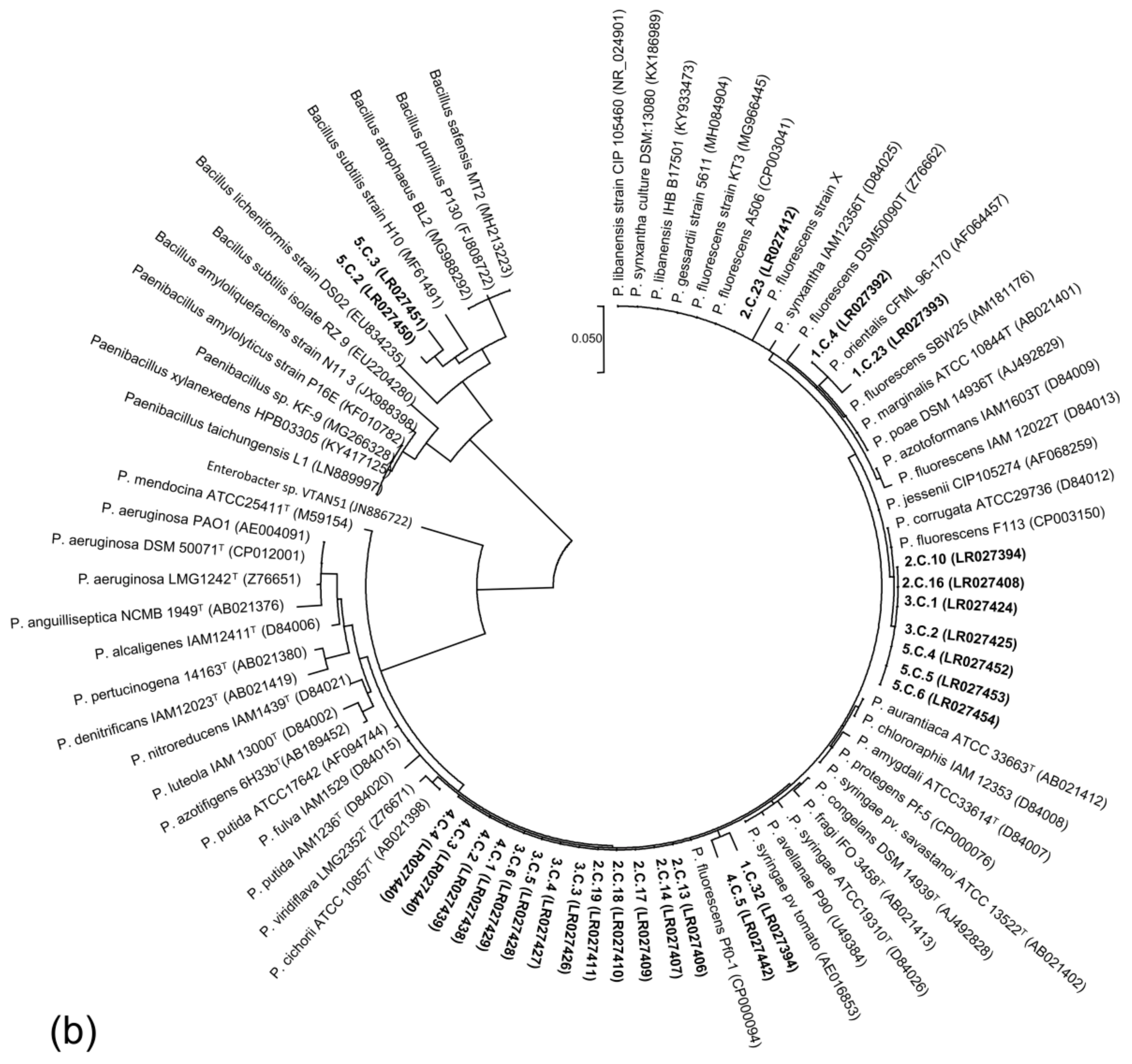
2.6. PCR Amplification 16S rRNA Gene Sequence and Phylogenetic Analysis
2.7. Nucleotide Sequence Accession Numbers
2.8. Statistical Analysis
3. Results
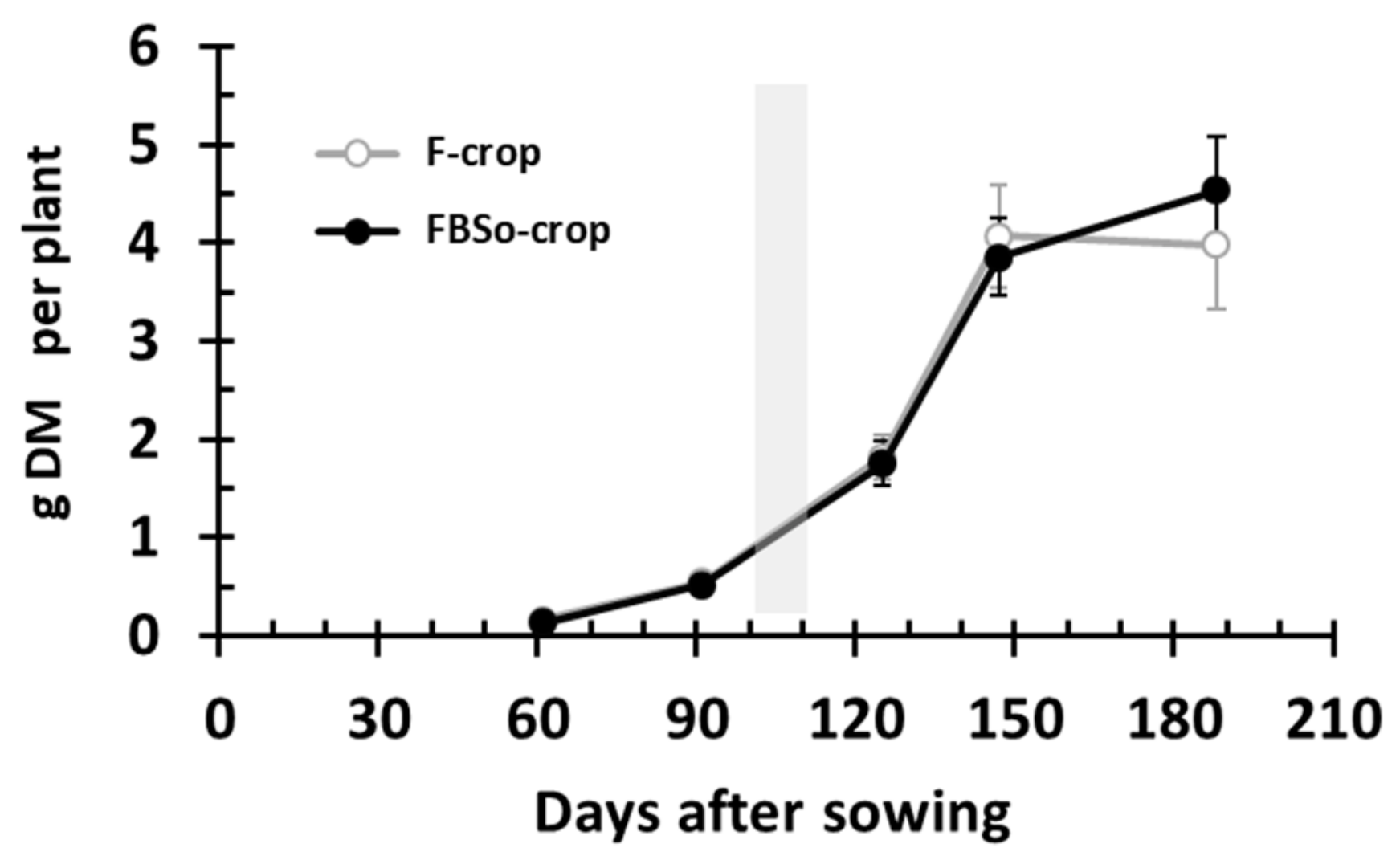
3.1. Dry Mass Accumulation per Plant
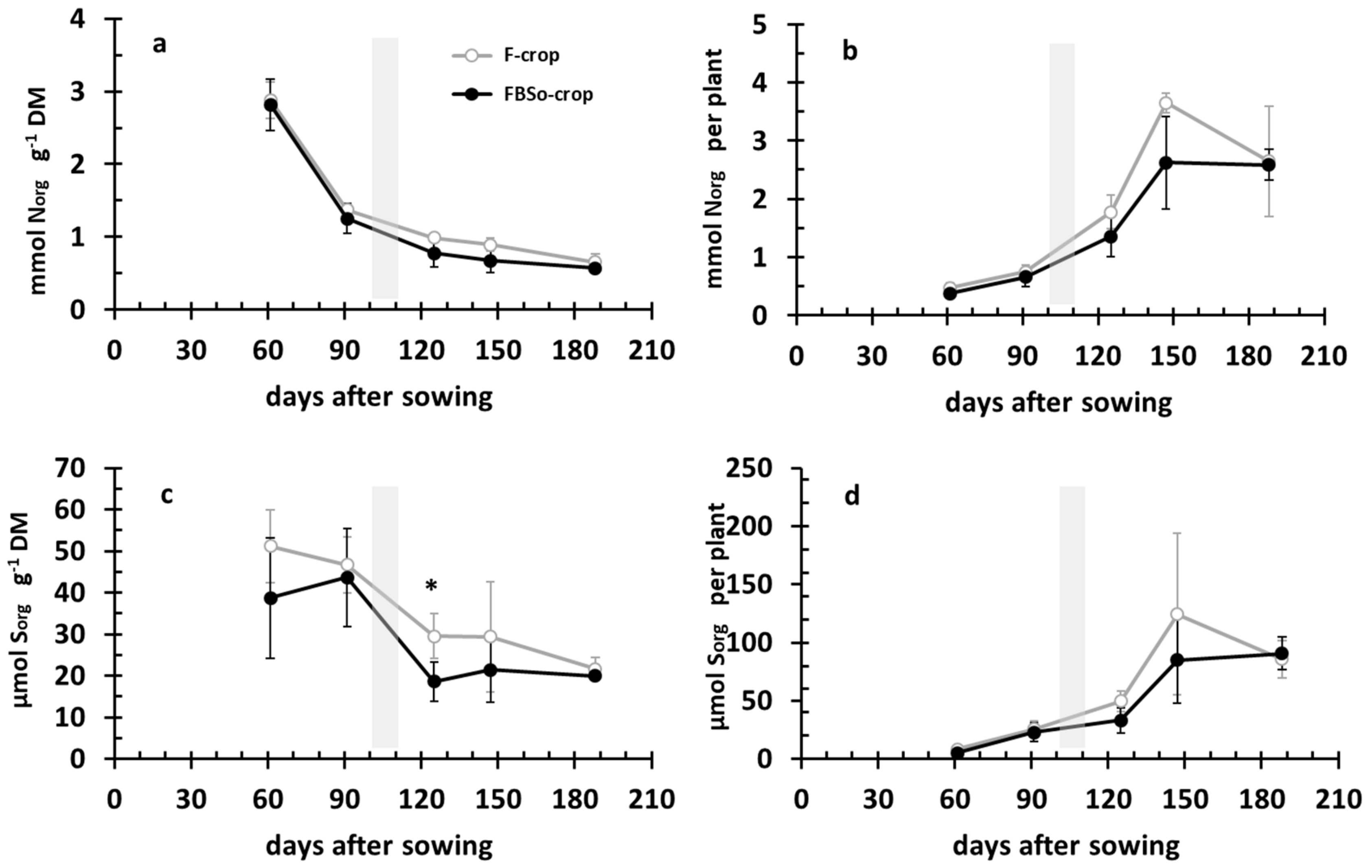
3.2. Developmental Dynamics of Organic Nitrogen versus Organic Sulfur in the Crop’s Aerial Part
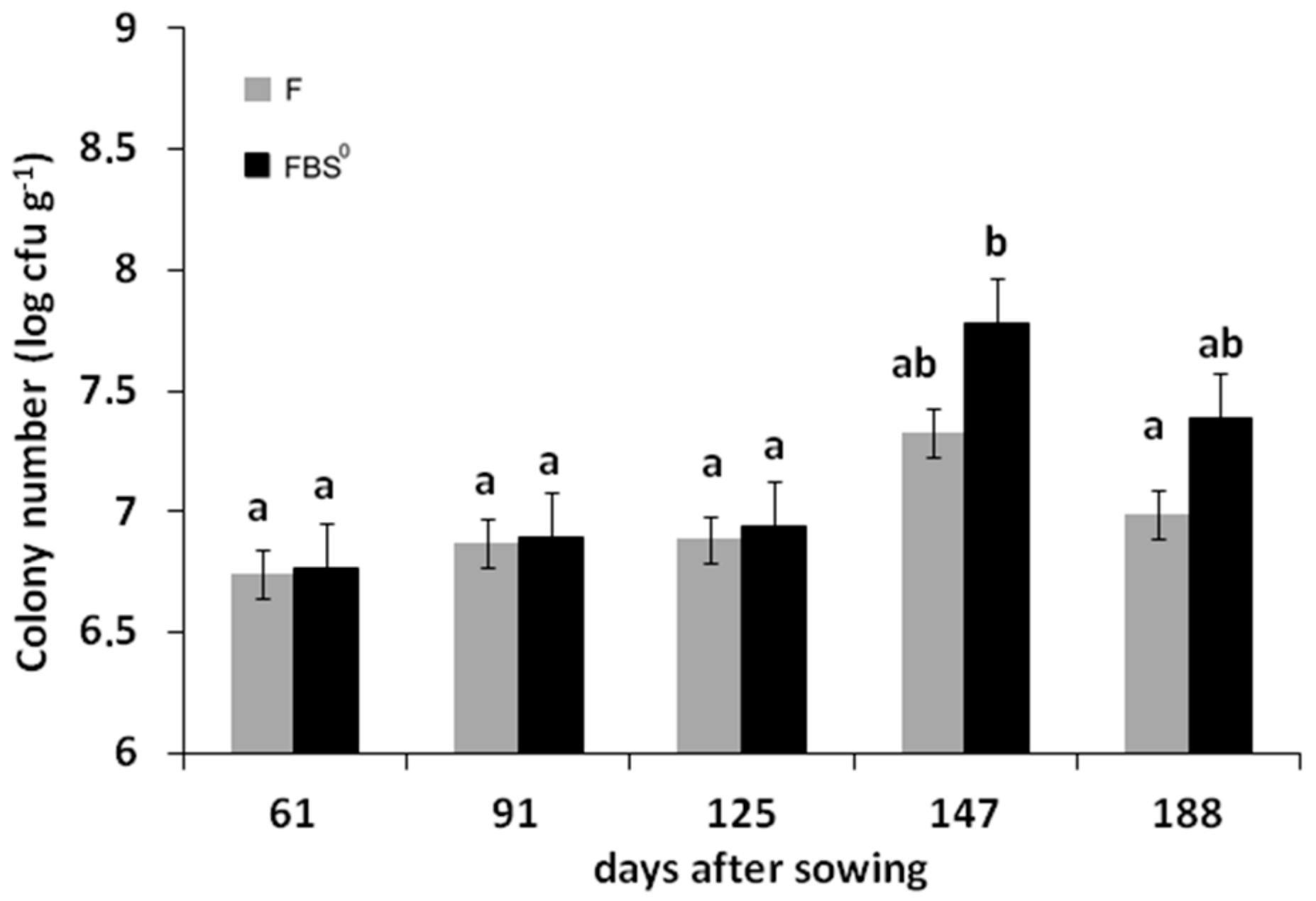
3.3. Quantification of Cultivable ARS-Producing Bacterial Communities in the Rhizosphere of F- and FBS0-Treated Wheat
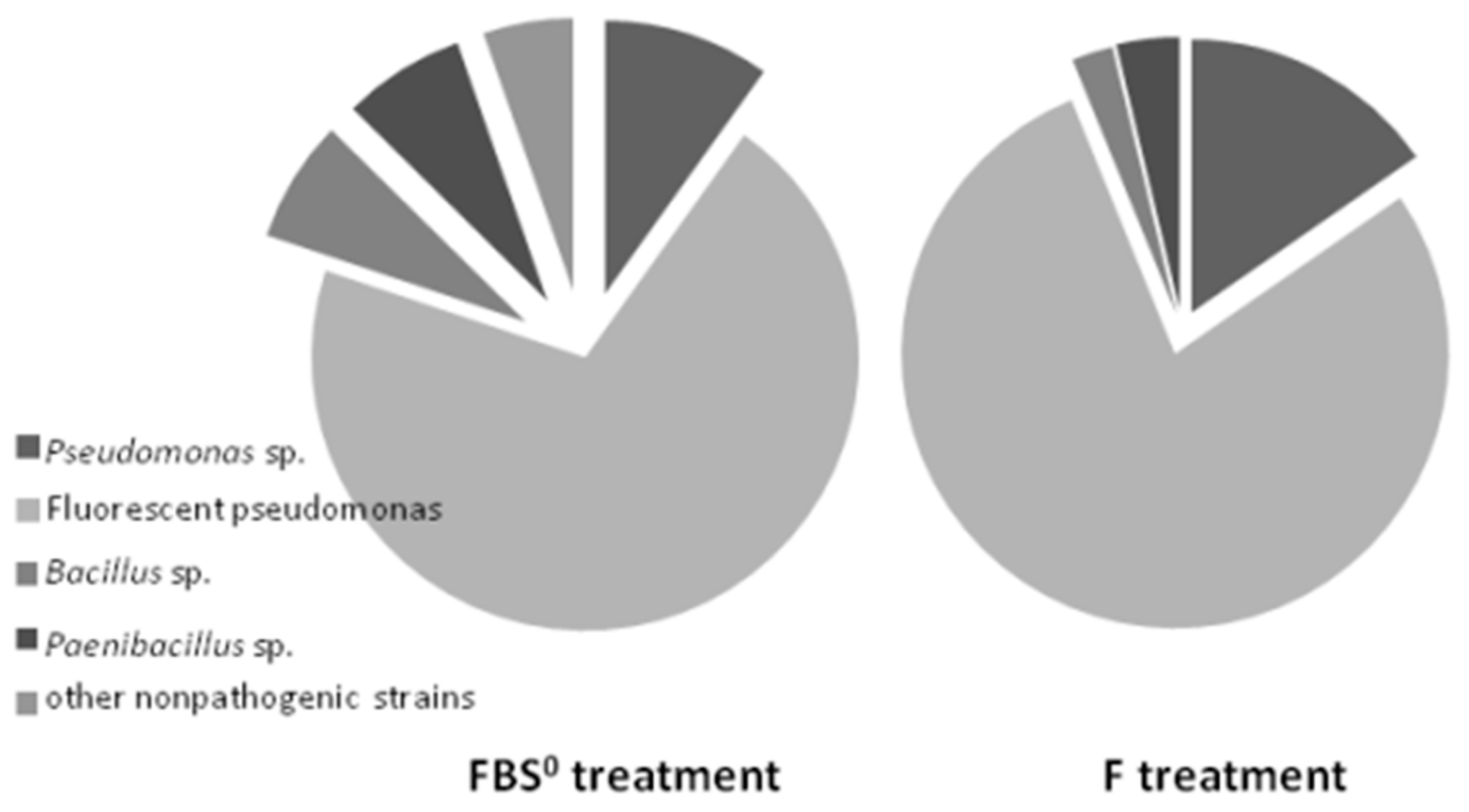
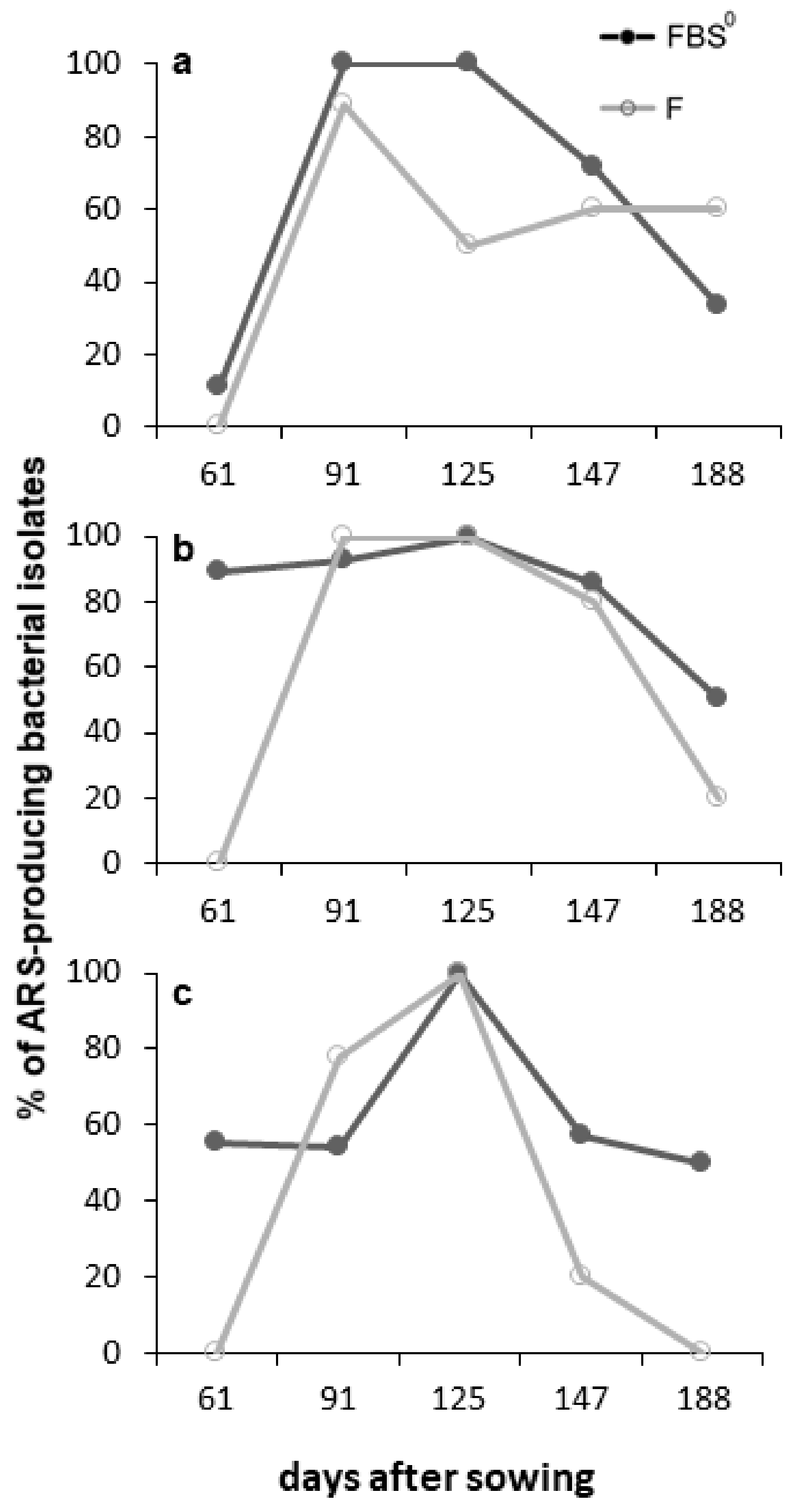
3.4. Diversity of ARS-Producing Bacteria
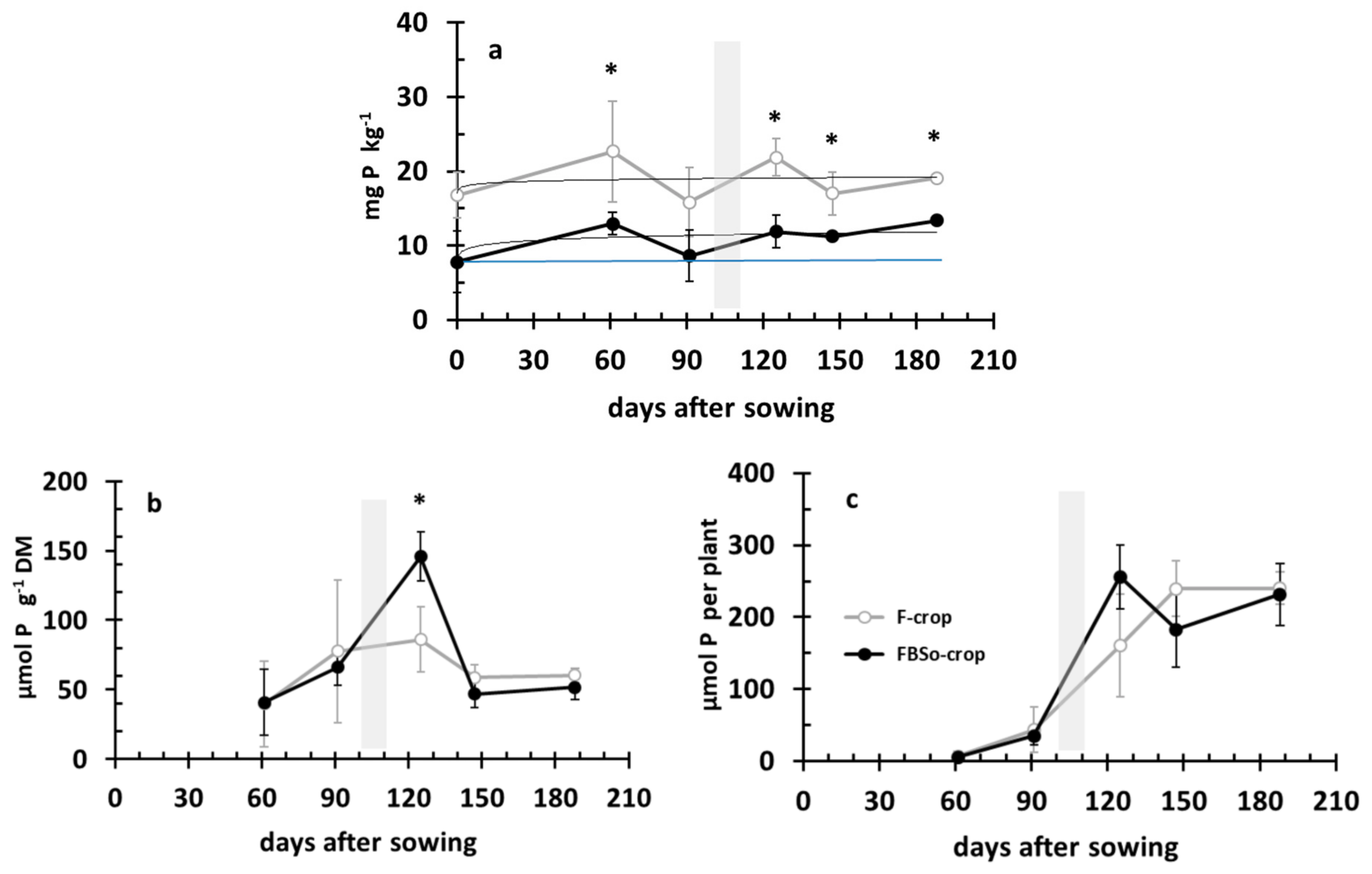
3.5. Phosphorus Dynamics in the Rhizosoil and the Aerial Part of the Plant
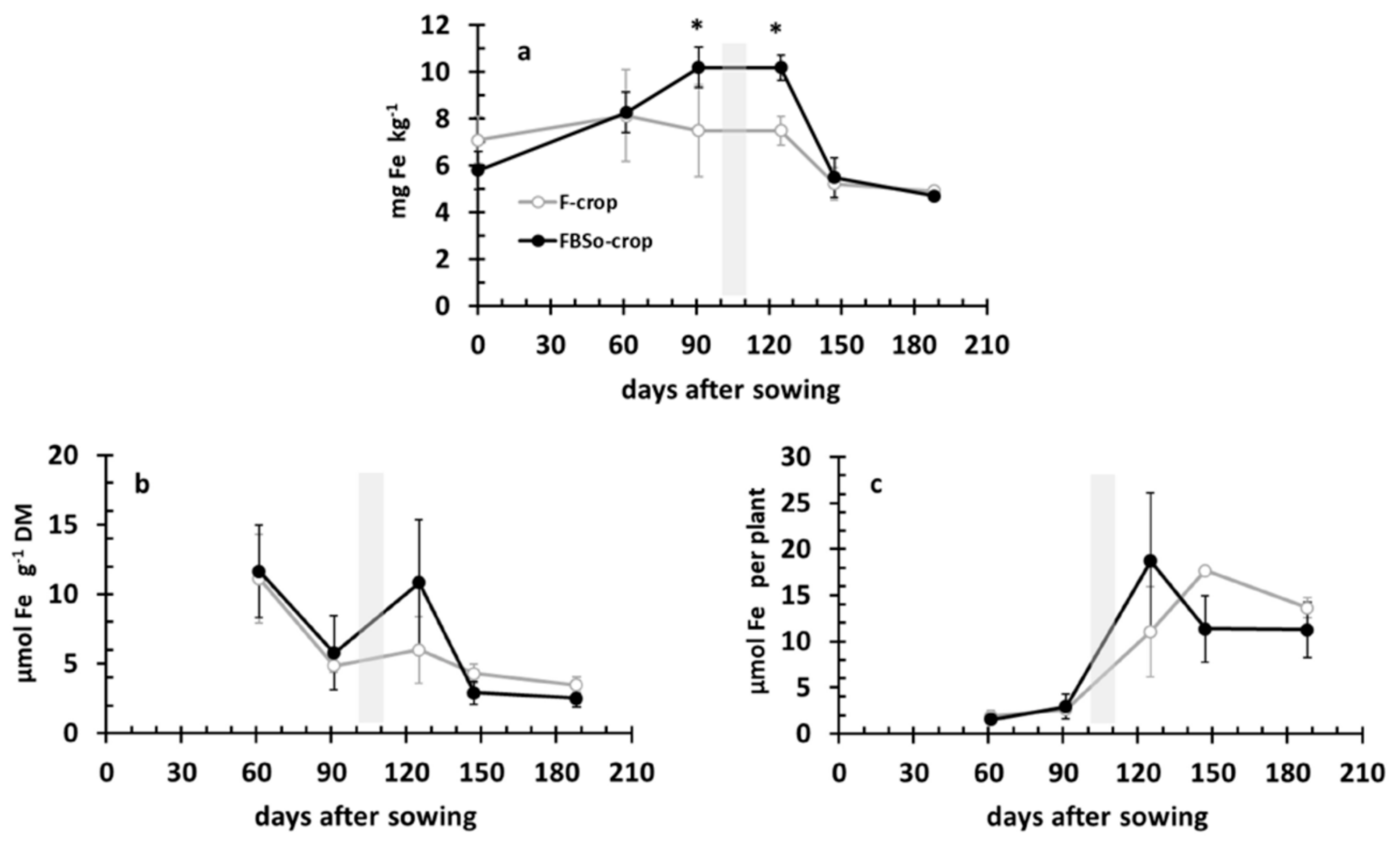
3.6. Iron Dynamics in the Rhizosoil and the Aerial Part of the Plant
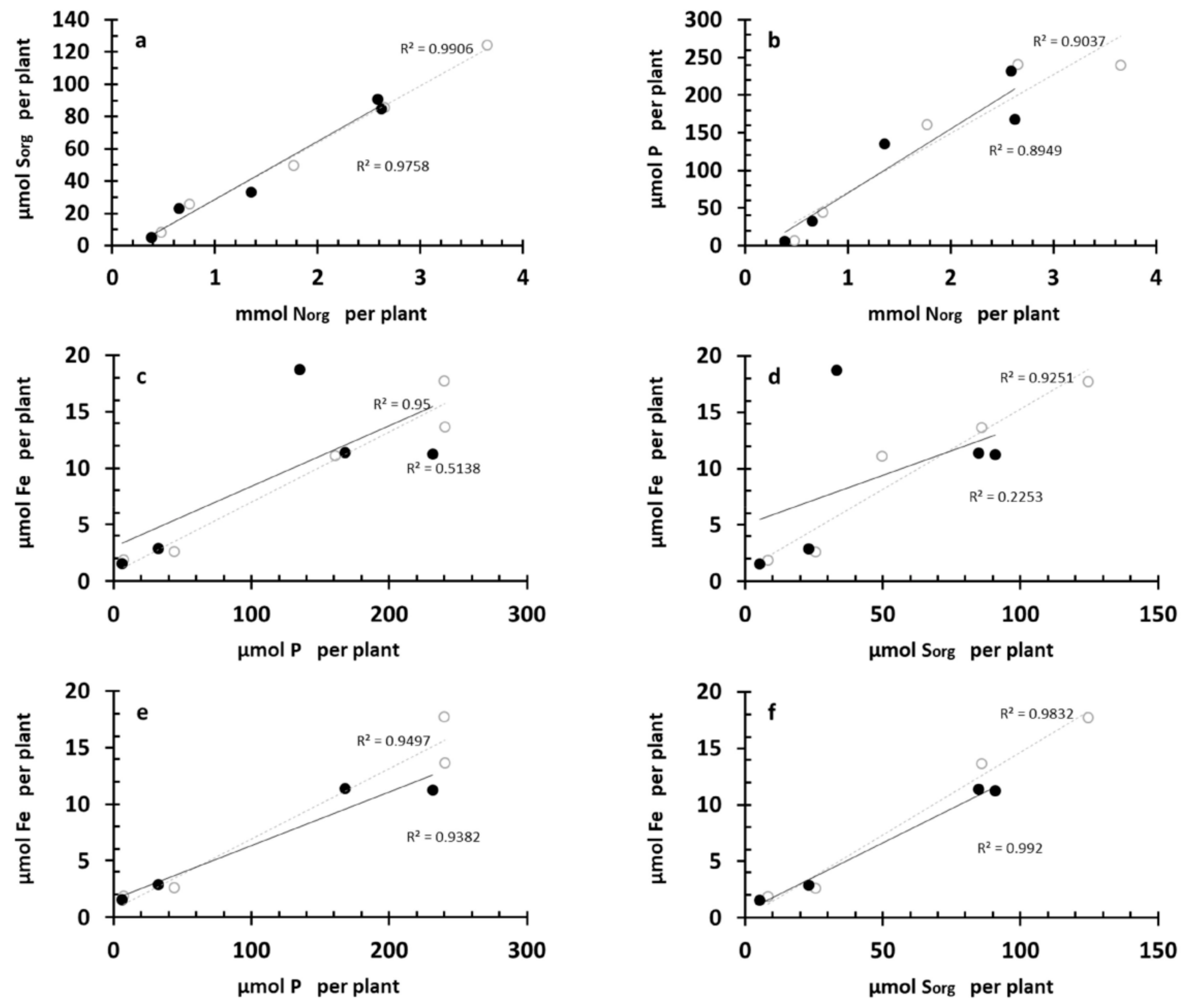
3.7. Correlations between Sorg, Fe, P, and Norg Crops’ Nutritional Dynamics
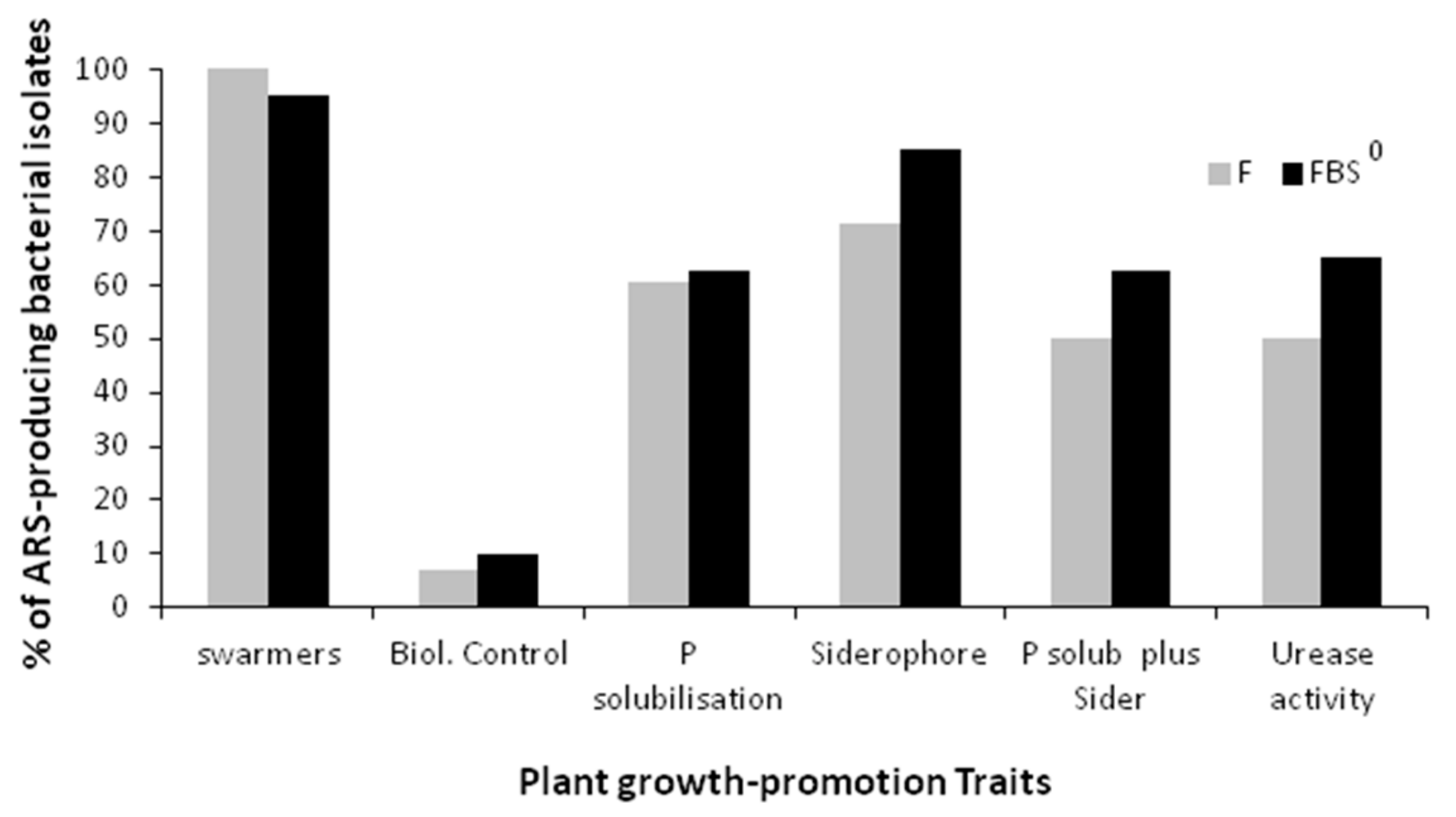
3.8. Phosphate Solubilization, and/or Siderophore-Producing Capacity, and/or Ureolytic Activity of the ARS-Producing Bacterial Isolates
4. Discussion
4.1. The Addition of Elemental Sulfur to the Fertilization Scheme Promoted a Higher Density of ARS-Producing Bacterial Population in the Rhizosphere
4.2. A Large Fraction of the ARS-Producing Bacterial Isolates Possessed Phosphate Solubilization Capacity
4.3. A Large Fraction of the ARS-Producing Bacterial Isolates Possessed Siderophore-Producing Capacity
4.4. A Large Fraction of the ARS-Producing Bacterial Isolates Possessed Ureolytic Activity
4.5. Towards a Sustainable Agronomic Biofortification Practice
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2011. [Google Scholar]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant. Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Ahkamia, A.H.; White, A.R., III; Handakumburaa, P.P.; Janssona, C. Rhizosphere engineering: Enhancing sustainable plant ecosystem productivity. Rhizosphere 2017, 3, 233–243. [Google Scholar] [CrossRef]
- Rana, A.; Joshi, M.; Prassara, R.; Shivay, Y.S.; Nain, L. Biofortification of wheat through inoculation of plant growth promoting rhizobacteria and cyanobacteria. Eur. J. Soil Biol. 2012, 50, 118–126. [Google Scholar] [CrossRef]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant. Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Adnan, M.; Shah, Z.; Fahad, S.; Arif, M.; Alam, M.; Khan, I.A.; Mian, I.A.; Basir, A.; Ullah, H.; Arshad, M.; et al. Phosphate-solubilizing bacteria nullify the antagonistic effect of soil calcification on bioavailability of phosphorus in alkaline soils. Sci. Rep. 2017, 7, 16131. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.K.; Ma, J.J.; Liu, L.J. Effects of phosphate-solubilizing bacteria application on soil phosphorus availability in coal mining subsidence area in Shanxi. J. Plant. Interact. 2017, 12, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Oteino, N.; Lally, R.D.; Kiwanuka, S.; Lloyd, A.; Ryan, D.; Germaine, K.J.; Dowling, D. Plant growth promotion induced by phosphate solubilizing endophytic Pseudomonas isolates. Front. Microbiol. 2015, 6, 745. [Google Scholar] [CrossRef] [PubMed]
- Zaida, A.; Khan, M.S.; Amil, M. Interactive effect of rhizotrophic microorganisms on yield and nutrient uptake of chickpea (Cicer arietinum L.). Eur. J. Agron. 2003, 19, 15–21. [Google Scholar] [CrossRef]
- Hameeda, B.; Harini, G.; Rupela, O.P.; Wani, S.P.; Reddy, G. Growth promotion of maize by phosphate-solubilizing bacteria isolated from compost and macrofauna. Microb. Res. 2008, 163, 234–242. [Google Scholar] [CrossRef]
- García-López, A.M.; Recena, R.; Avilés, M.; Delgado, A. Effect of Bacillus subtilis QST713 and Trichoderma asperellum T34 on P uptake by wheat and how it is modulated by soil properties. J. Soils Sediments 2018, 18, 727–773. [Google Scholar] [CrossRef]
- Kumar, V.; Singh, P.; Jorquera, M.A.; Sangwan, P.; Kumar, P.; Verma, A.K.; Agrawal, S. Isolation of phytase-producing bacteria from Himalayan soils and their effect on growth and phosphorus uptake of Indian mustard (Brassica juncea). World J. Microbiol. Biotechnol. 2013, 29, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Rungin, S.; Indananda, C.; Suttiviriya, P.; Kruasuwan, W.; Jaemsaeng, R.; Thamchaipenet, A. Plant growth enhancing effects by a siderophore-producing endophytic streptomycete isolated from a Thai jasmine rice plant (Oryza sativa L. cv. KDML105). Antonie Van Leeuwenhoek 2012, 102, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Khalid, S.; Asghar, H.N.; Akhtar, M.J.; Aslam, A.; Zahir, Z.A. Biofortification of iron in chickpea by plant growth promoting rhizobacteria. Pak. J. Bot. 2015, 47, 1191–1194. [Google Scholar]
- Liu, D.; Yang, Q.; Ge, K.; Hu, X.; Qi, G.; Du, B.; Liu, K.; Ding, Y. Promotion of iron nutrition and growth on peanut by Paenibacillus illinoisensis and Bacillus sp. strains in calcareous soil. Braz. J. Microbiol. 2017, 48, 656–670. [Google Scholar] [CrossRef] [PubMed]
- De Santiago, A.; García-López, A.M.; Quintero, J.M.; Avilés, M.; Delgado, A. Effect of Trichoderma asperellum strain T34 and glucose addition on iron nutrition in cucumber grown on calcareous soils. Soil Biol. Biochem. 2013, 5, 598–605. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Boyce, A.N. Role of plant growth promoting rhizobacteria in agricultural sustainability—A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef]
- Pii, Y.; Mimmo, T.; Tomasi, N.; Terzano, R.; Cesco, S.; Crecchio, C. Microbial interactions in the rhizosphere: Beneficial influences of plant growth promoting rhizobacteria on nutrient acquisition process. A review. Biol. Fertil. Soils 2015, 51, 403–415. [Google Scholar] [CrossRef]
- Kertesz, M.A.; Fellows, E.; Schmalenberger, A. Rhizobacteria and plant sulfur supply. Adv. Appl. Microbiol. 2007, 62, 235–268. [Google Scholar]
- Gahan, J.; Schmalenberger, A. The role of bacteria and mycorrhiza in plant sulfur supply. Front. Plant. Sci. 2014. [Google Scholar] [CrossRef]
- Fox, A.; Kwapinski, W.; Grifffiths, B.S.; Schmalenberger, A. The role of sulphur- and phosphorus mobilizing bacteria in biochar-induced growth promotion of Lolium perenne. FEMS Microbiol. Ecol. 2014, 90, 78–91. [Google Scholar] [CrossRef]
- Crégut, M.; Piutti, S.; Vong, P.-C.; Slezack-Deschaumes, S.; Crovisier, I.; Benizri, E. Density, structure, and diversity of the cultivable arylsulfatase-producing bacterial community in the rhizosphere of field-grown rape and barley. Soil Biol. Biochem. 2009, 41, 704–710. [Google Scholar] [CrossRef]
- Crégut, M.; Piutti, S.; Slezack-Deschaumes, S.; Benizri, E. Compartmentalization and regulation of arylsulfatase activities in Streptomyces sp., Microbacterium sp. and Rhodococcus sp. soil isolates in response to inorganic sulfate limitation. Microbiol. Res. 2013, 168, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Whalen, J.K.; Warman, P.R. Arylsulfatase activity in soil and soil extracts using natural and artificial substrates. Biol. Fertil. Soils 1996, 22, 373–378. [Google Scholar] [CrossRef]
- Meldau, D.G.; Meldau, S.; Hoang, L.H.; Underberg, S.; Wunsche, H.; Baldwin, I.T. Dimethyl disulfide produced by the naturally associated bacterium Bacillus sp B55 promotes Nicotia attenuata growth by enhancing sulfur nutrition. Plant. Cell 2013, 25, 2731–2747. [Google Scholar] [PubMed]
- Bouranis, D.L.; Chorianopoulou, S.N.; Margetis, M.; Saridis, G.I.; Sigalas, P.P. Effect of Elemental Sulfur as Fertilizer Ingredient on the Mobilization of Iron from the Iron Pools of a Calcareous Soil Cultivated with Durum Wheat and the Crop’s Iron and Sulfur Nutrition. Agriculture 2018, 8, 20. [Google Scholar] [CrossRef]
- Bouranis, D.L.; Gasparatos, D.; Zechmann, B.; Bouranis, L.D.; Chorianopoulou, S.N. The Effect of Granular Commercial Fertilizers Containing Elemental Sulfur on Wheat Yield under Mediterranean Conditions. Plants 2019, 8, 2. [Google Scholar] [CrossRef]
- Jones, J.B., Jr. Soil Analysis Handbook of Reference Methods; CRC Press: Boca Raton, FL, USA, 1999; ISBN 9780849302053. [Google Scholar]
- Sörbo, B. Sulfate: Turbidimetric and nephelometric methods. In Methods in Enzymology: Sulfur and Sulfur Amino Acids; Jakoby, W.B., Griffith, O.W., Eds.; Academic Press, Inc.: New York, NY, USA, 1987. [Google Scholar]
- Miller, R.O. Extractable chloride, nitrate, orthophosphate, potassium, and sulfate sulfur in plant tissue: 2% acetic acid extraction. In Handbook of Reference Methods for Plant Analysis; Kalra, Y.P., Ed.; CRC Press LLC: Boca Raton, FL, USA, 1998; pp. 115–118. [Google Scholar]
- Chorianopoulou, S.N.; Nikologiannis, S.; Gasparatos, D.; Bouranis, D.L. Relationships between iron, sulfur, nitrogen and phosphorus in lawns grown on a calcareous soil irrigated by slightly saline water. Fresenius Environ. Bull. 2017, 26, 1240–1246. [Google Scholar]
- Pikovskaya, R. Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Mikrobiologiya 1948, 17, 362–370. [Google Scholar]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Venieraki, A.; Tsalgatidou, P.C.; Georgakopoulos, D.G.; Dimou, M.; Katinakis, P. Swarming motility in plant-associated bacteria. Hellenic Plant. Protect. J. 2016, 9, 16–27. [Google Scholar] [CrossRef] [Green Version]
- Weisburg, W.S.; Barns, S.M.; Pelletier, D.A.; Lane, D.I. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evolut. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Astolfi, S.; Cesco, S.; Zuchi, S.; Neumann, G.; Roemheld, V. Sulphur starvation reduces phytosiderophores release by Fe-deficient barley plants. Soil Sci. Plant. Nutr. 2006, 52, 80–85. [Google Scholar] [CrossRef]
- Astolfi, S.; Zuchi, S.; Cesco, S.; Sanita di Toppi, L.; Pirazzi, D.; Badiani, M.; Varanini, Z.; Pinton, R. Fe deficiency induces sulphate uptake and modulates redistribution of reduced sulphur pool in barley plants. Funct. Plant. Biol. 2006, 33, 1055–1061. [Google Scholar] [CrossRef]
- Chorianopoulou, S.N.; Saridis, Y.I.; Dimou, M.; Katinakis, P.; Bouranis, D.L. Arbuscular mycorrhizal symbiosis alters the expression patterns of three key iron homeostasis genes, ZmNAS1, ZmNAS3, and ZmYS1, in S deprived maize plants. Front. Plant. Sci. 2015, 6, 257. [Google Scholar] [CrossRef] [PubMed]
- Saridis, G.I.; Chorianopoulou, S.N.; Katinakis, P.; Bouranis, D.L. Evidence for regulation of the iron uptake pathway by sulfate supply in S-deprived maize plants. In Sulfur Metabolism in Higher Plants—Fundamental, Environmental and Agricultural Aspects, Proceedings of the International Plant Sulfur Workshop, Goslar, Germany, 1–4 September 2015; De Kok, L.J., Hawkesford, M.J., Haneklaus, S.H., Schnug, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 175–180. [Google Scholar]
- Saridis, G.; Chorianopoulou, S.N.; Ventouris, Y.E.; Sigalas, P.P.; Bouranis, D.L. An Exploration of the Roles of Ferric Iron Chelation-Strategy Components in the Leaves and Roots of Maize Plants. Plants 2019, 8, 133. [Google Scholar] [CrossRef] [PubMed]
- Astolfi, S.; Zuchi, S.; Neumann, G.; Cesco, S.; Sanita di Toppi, L.; Pinton, R. Response of barley plants to Fe deficiency and Cd contamination as affected by S starvation. J. Exp. Bot. 2012, 63, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Astolfi, S.; Zuchi, S.; Passera, C.; Cesco, S. Does the sulfur assimilation pathway play a role in the response to Fe deficiency in maize (Zea mays L.) plants? J. Plant. Nutr. 2003, 26, 2111–2121. [Google Scholar] [CrossRef]
- Calvo, P.; Watts, D.B.; Kloepper, J.W.; Torbert, H.A. Effect of microbial-based inoculants on nutrient concentrations and early root morphology of corn (Zea mays). J. Plant. Nutr. Soil Sci. 2016, 180, 56–70. [Google Scholar] [CrossRef]
- Piutti, S.; Slezack-Deschaumes, S.; Niknahad-Gharmakher, H.; Vong, P.C.; Recous, S.; Benizri, E. Relationships between the density and activity of microbial communities possessing arylsulfatase activity and soil sulfate dynamics during the decomposition of plant residues in soil. Eur. J. Soil Biol. 2015, 70, 88–96. [Google Scholar] [CrossRef]
- Tang, H.; Xiao, X.; Sun, J.; Guo, L.; Wang, K.; Li, E.; Wenguang, T. Soil enzyme activities and soil microbe population as influenced by long-term fertilizer management during the barley growth in Hunan Province, China. Afr. J. Microbiol. Res. 2016, 10, 1720–1727. [Google Scholar] [Green Version]
- Rossini, F.; Provenzano, M.E.; Sestili, F.; Ruggeri, R. Synergistic effect of sulfur and nitrogen in the organic and mineral fertilization of durum wheat: Grain yield and quality traits in the Mediterranean environment. Agronomy 2018, 8, 189. [Google Scholar] [CrossRef]
- Vong, P.-C.; Nguyen, C.; Guckert, A. Fertilizer sulphur uptake and transformations in soil as affected by plant species and soil type. Eur. J. Agron. 2007, 27, 35–43. [Google Scholar] [CrossRef]
- Hu, Z.Y.; Zhao, F.J.; McGrath, S.P. Sulphur fractionation in calcareous soils and bioavailability to plants. Plant. Soil 2005, 268, 103–109. [Google Scholar] [CrossRef]
- Chien, S.H.; Gearhart, M.M.; Villagarcia, S. Comparison of ammonium sulfate with other nitrogen and sulfur fertilizers in increasing crop production and minimizing environmental impact: A review. Soil Sci. 2011, 176, 327–335. [Google Scholar] [CrossRef]
- Recena, R.; Díaz, I.; Delgado, A. Estimation of total plant available phosphorus in representative soils from Mediterranean areas. Geoderma 2017, 297, 10–18. [Google Scholar] [CrossRef]
- Houben, D.; Michel, E.; Nobile, C.; Lambers, H.; Kandeler, E.; Faucon, M.-P. Response of phosphorus dynamics to sewage sludge application in an agroecosystem in northern France. Appl. Soil Ecol. 2019, 137, 178–186. [Google Scholar] [CrossRef]
- Ipek, M.; Pirlak, L.; Esitken, A.; Figen, M.; Dönmez, M.; Turan, M.; Sahin, F. Plant growth promoting rhizobacteria (PGPR) increase yield, growth and nutrition of Strawberry under high-calcareous soil conditions. J. Plant. Nutr. 2014, 37, 990–1001. [Google Scholar] [CrossRef]
- Iqbal, S.; Khan, M.Y.; Asghar, H.N.; Akhtar, M.J. Combined use of phosphate solubilizing bacteria and poultry manure to enhance the growth and yield of mung bean in calcareous soil. Soil Environ. 2014, 35, 146. [Google Scholar]
- Ibarra-Galeana, J.A.; Castro-Martínez, C.; Fierro-Coronado, R.A.; Armenta-Bojórquez, A.D.; Maldonado-Mendoza, I.E. Characterization of phosphate-solubilizing bacteria exhibiting the potential for growth promotion and phosphorus nutrition improvement in maize (Zea mays L.) in calcareous soils of Sinaloa, Mexico. Ann. Microbiol. 2017, 67, 801–807. [Google Scholar] [CrossRef]
- Sharma, A.; Shankhdhar, D.; Shankhdhar, S.C. Enhancing grain iron content of rice by the applicationof plant growth promoting rhizobacteria. Plant. Soil Environ. 2013, 59, 89–94. [Google Scholar] [CrossRef]
- Abbasi, M.K.; Sharif, S.; Kazmi, M.; Sultan, T.; Aslam, M. Isolation of plant growth promoting rhizobacteria from wheat rhizosphere and their effect on improving growth, yield and nutrient uptake of plants. Plant. Biosyst. 2011, 145, 159–168. [Google Scholar] [CrossRef]
- Tscherko, D.; Kandeler, E.; Bardossy, A. Fuzzy classification of microbial biomass and enzyme activities in grassland soils. Soil Biol. Biochem. 2007, 39, 1799–1808. [Google Scholar] [CrossRef]
- Zhao, F.J.; Hawkesford, M.J.; McGrath, S.P. Sulphur assimilation and effects on yield and quality of wheat. J. Cereal. Sci. 1999, 30, 1–17. [Google Scholar] [CrossRef]
- Ercoli, L.; Lulli, L.; Arduini, I.; Mariotti, M.; Masoni, A. Durum wheat grain yield and quality as affected by S rate under Mediterranean conditions. Eur. J. Agron. 2011, 35, 63–70. [Google Scholar] [CrossRef]
- Thomloudi, E.-E.; Tsalgatidou, P.C.; Douka, D.; Spantidos, T.-N.; Dimou, M.; Venieraki, A.; Katinakis, P. Multistrain versus single-strain plant growth promoting microbial inoculants—The compatibility issue. Hellenic Plant. Protect. J. 2019, 12, 61–77. [Google Scholar] [CrossRef]
- Li, Y.; Yu, S.; Strong, J.; Wang, H. Are the biogeochemical cycles of carbon, nitrogen, sulfur, and phosphorus driven by the “FeIII–FeII redox wheel” in dynamic redox environments? J. Soils Sediments 2012, 12, 683–693. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crop | Clay | Silt | Sand | Class | pH | EC | CaCO3 | SOM |
|---|---|---|---|---|---|---|---|---|
| % | % | % | μS cm−1 | % | % | |||
| F | 38 | 34 | 28 | Silty Clay | 7.86 | 685 | 31.5 | 2.55 |
| FBS0 | 42 | 30 | 28 | Clay | 8.01 | 613 | 24.5 | 1.79 |
| Δx/x (%) | 1.9 | −10.5 | −22.2 | −29.8 | ||||
| Olsen P | NO3-N | Kexc | Fe-DTPA | Mn-DTPA | Cu-DTPA | Zn-DTPA | ||
| mg kg−1 | mg kg−1 | mg kg−1 | mg kg−1 | mg kg−1 | mg kg−1 | mg kg−1 | ||
| F | 16.8 | 28.48 | 410 | 7.08 | 8.97 | 1.76 | 1.26 | |
| FBS0 | 7.8 | 24.57 | 270 | 5.78 | 6.25 | 1.43 | 0.75 | |
| Δx/x (%) | −53.6 | −13.7 | −34.1 | −18.4 | −30.3 | −18.8 | −40.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouranis, D.L.; Venieraki, A.; Chorianopoulou, S.N.; Katinakis, P. Impact of Elemental Sulfur on the Rhizospheric Bacteria of Durum Wheat Crop Cultivated on a Calcareous Soil. Plants 2019, 8, 379. https://doi.org/10.3390/plants8100379
Bouranis DL, Venieraki A, Chorianopoulou SN, Katinakis P. Impact of Elemental Sulfur on the Rhizospheric Bacteria of Durum Wheat Crop Cultivated on a Calcareous Soil. Plants. 2019; 8(10):379. https://doi.org/10.3390/plants8100379
Chicago/Turabian StyleBouranis, Dimitris L., Anastasia Venieraki, Styliani N. Chorianopoulou, and Panagiotis Katinakis. 2019. "Impact of Elemental Sulfur on the Rhizospheric Bacteria of Durum Wheat Crop Cultivated on a Calcareous Soil" Plants 8, no. 10: 379. https://doi.org/10.3390/plants8100379
APA StyleBouranis, D. L., Venieraki, A., Chorianopoulou, S. N., & Katinakis, P. (2019). Impact of Elemental Sulfur on the Rhizospheric Bacteria of Durum Wheat Crop Cultivated on a Calcareous Soil. Plants, 8(10), 379. https://doi.org/10.3390/plants8100379