Early Season Drought Largely Reduces Grain Yield in Wheat Cultivars with Smaller Root Systems




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growing Conditions
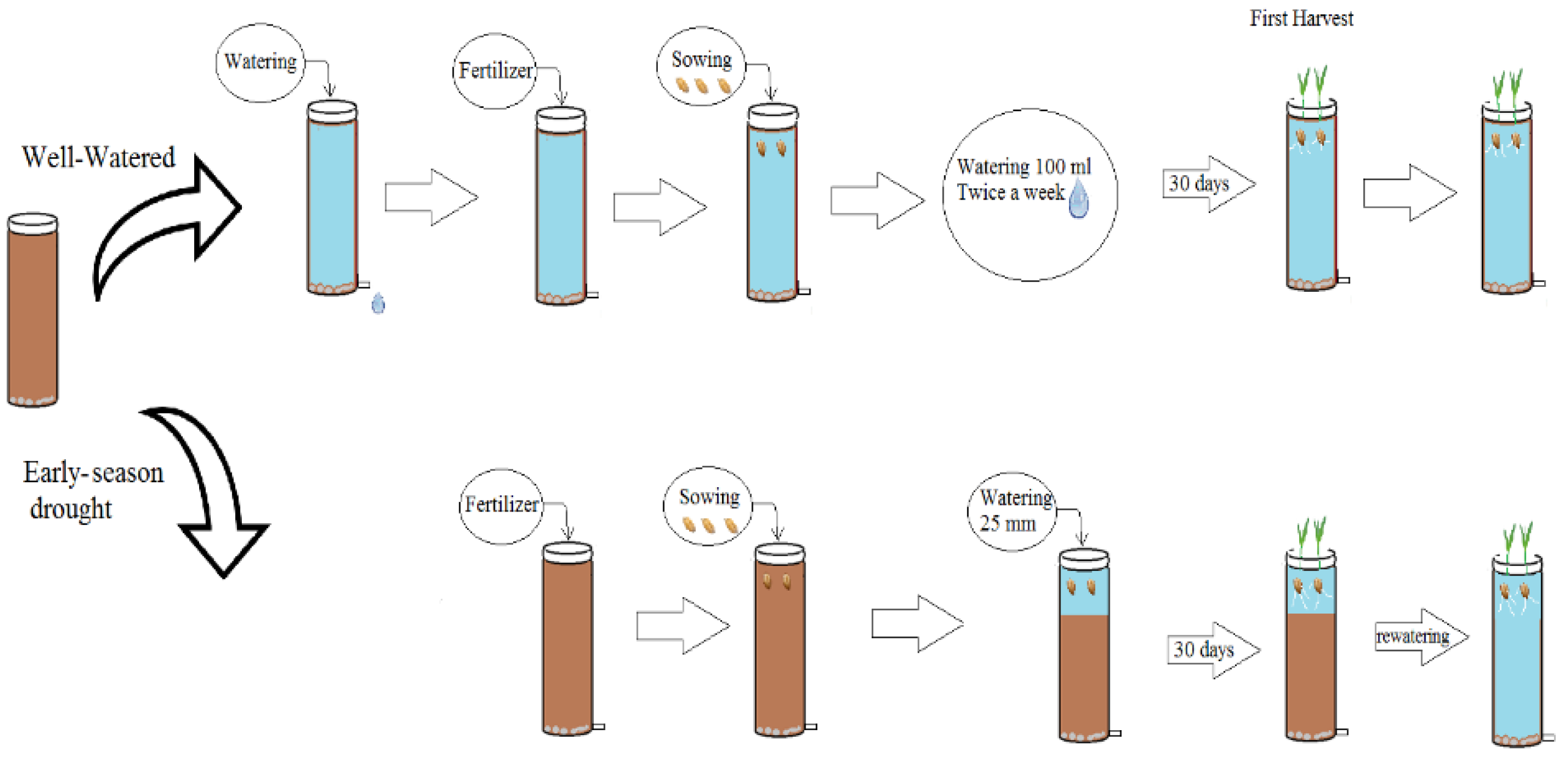
2.2. Experimental Design and Treatments
2.3. Measurements
2.4. Statistical Analysis
3. Results
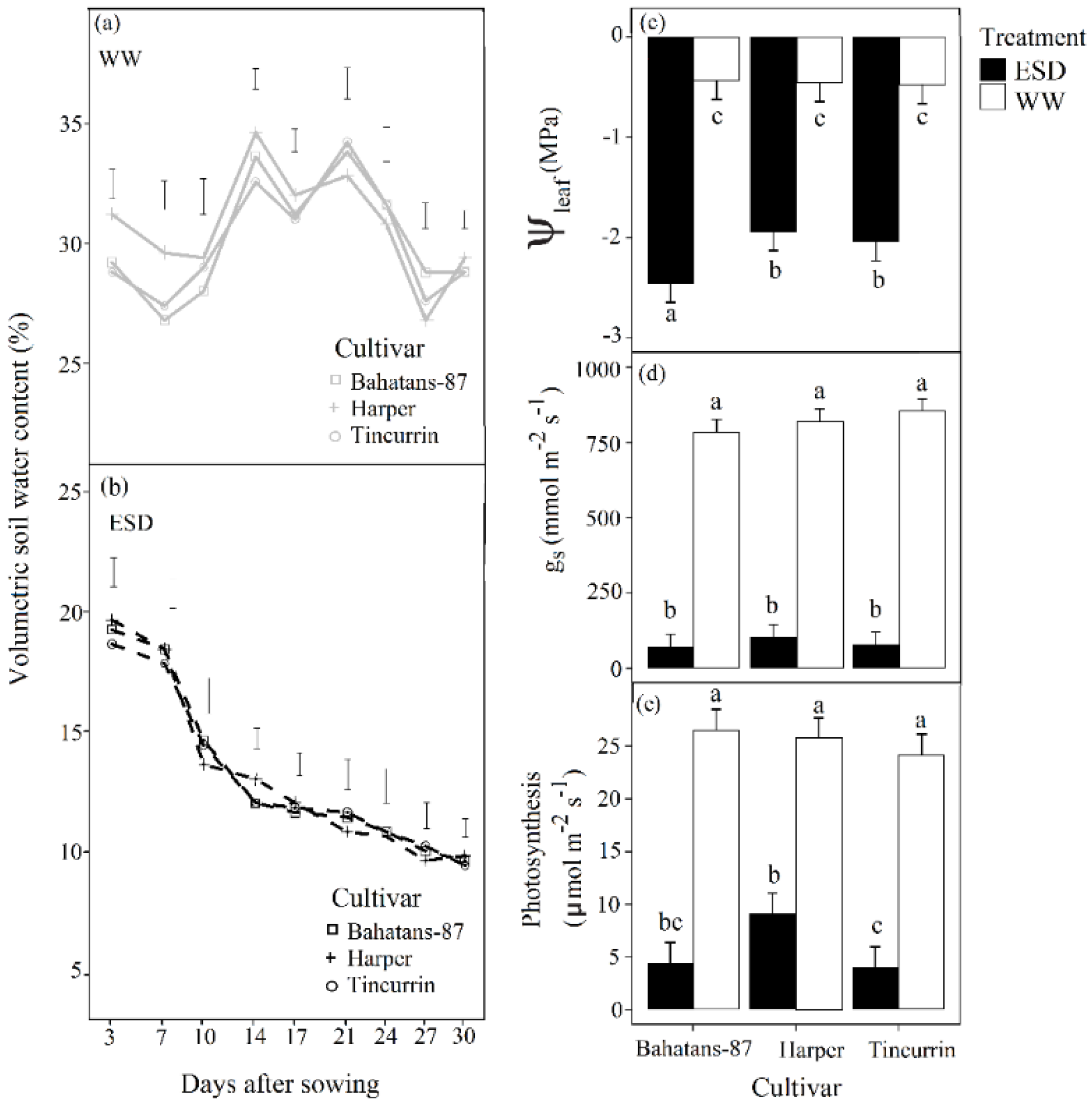
3.1. Soil Water Content, Water Potential, Stomatal Conductance and Leaf Photosynthesis
3.2. Phenology
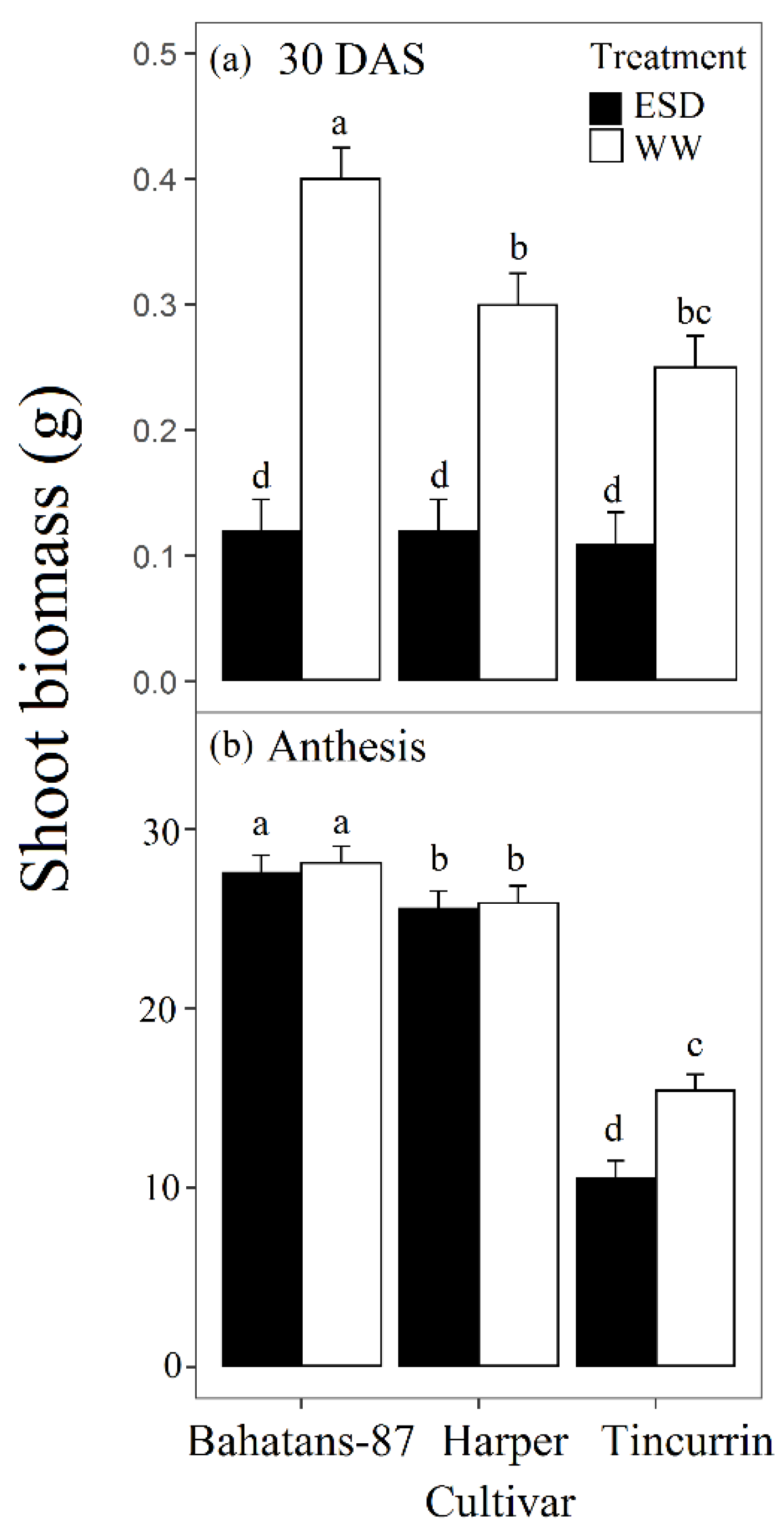
3.3. Shoot Traits
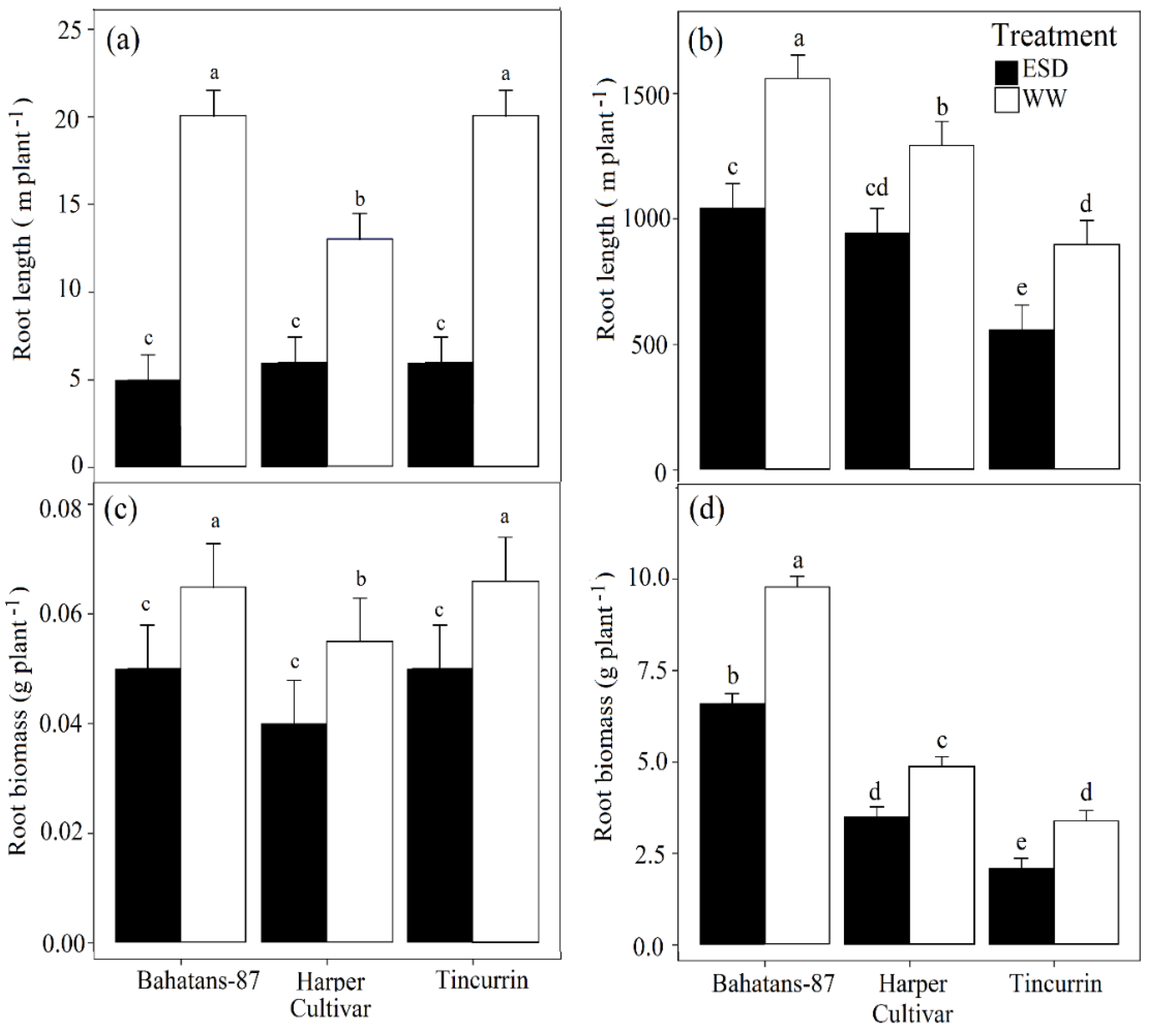
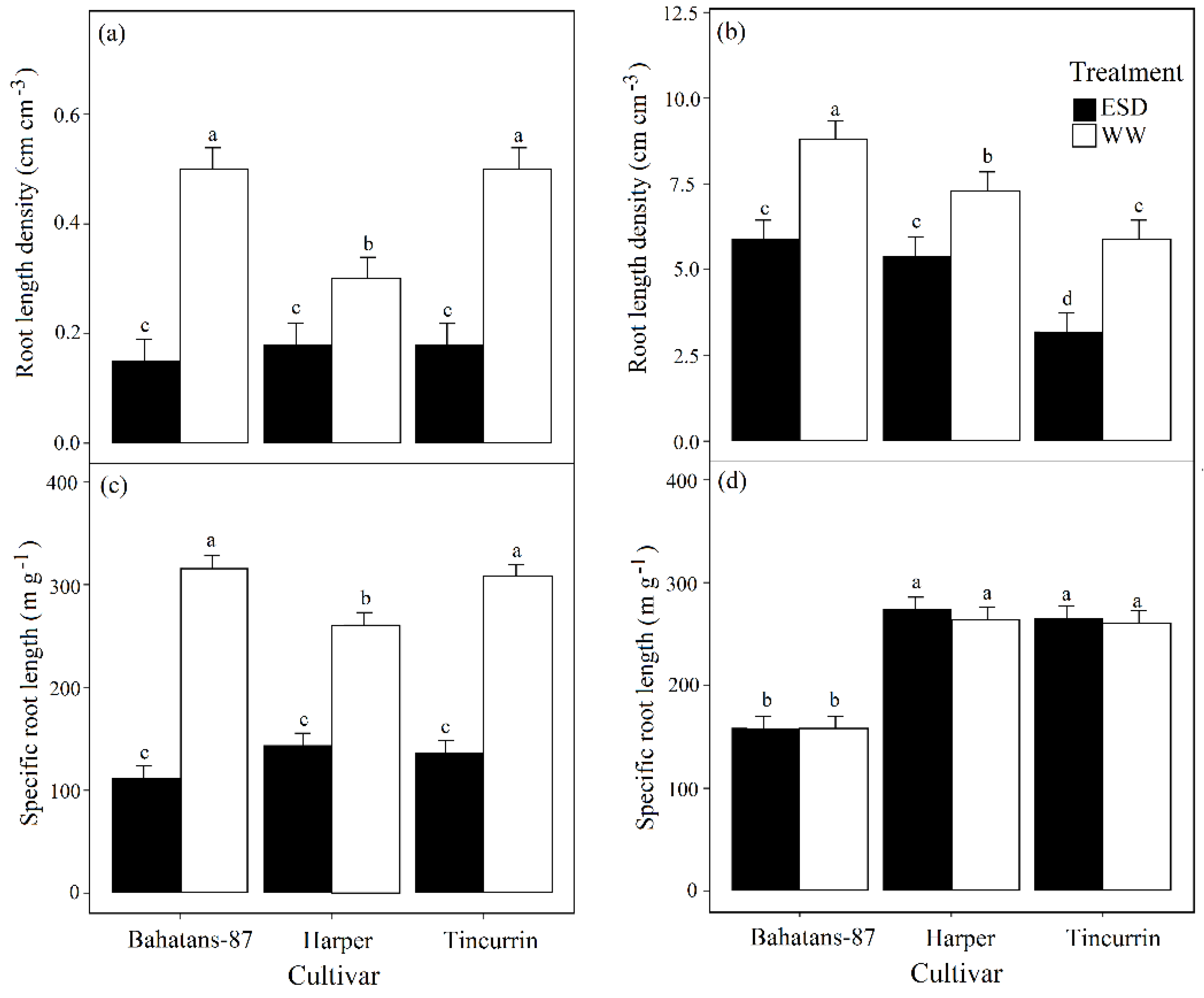
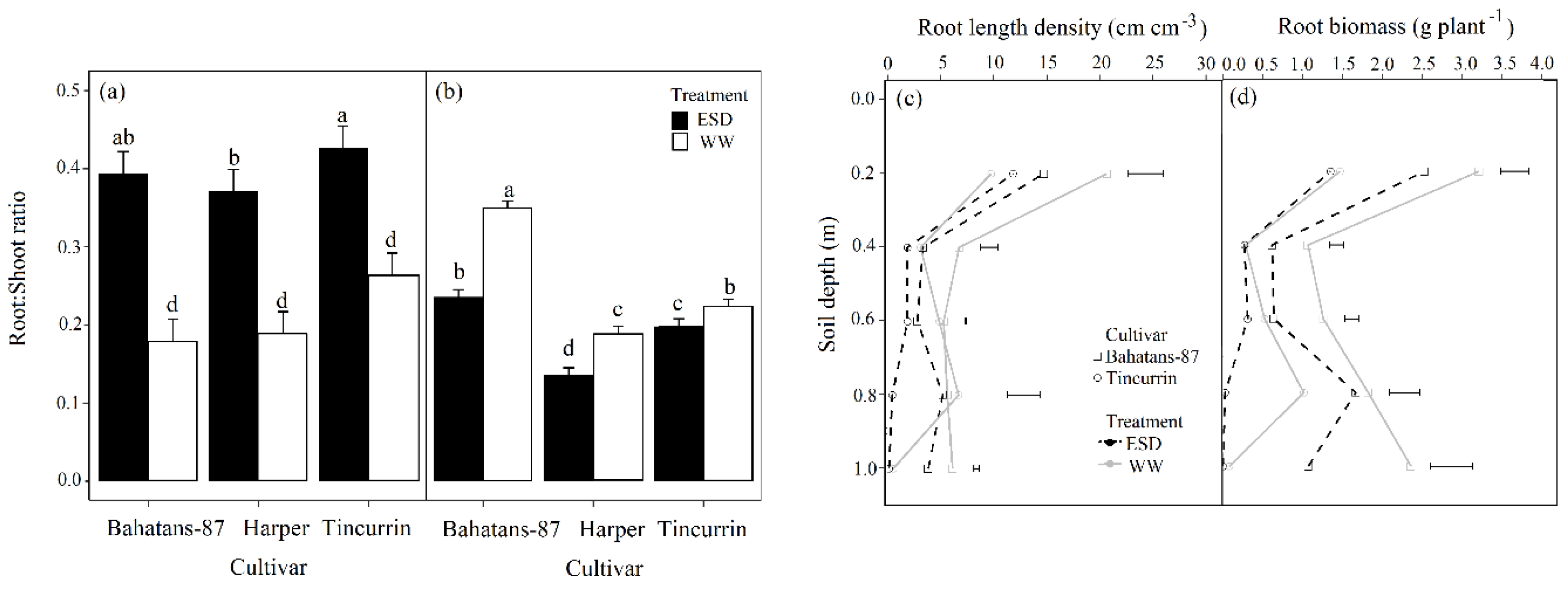
3.4. Root Traits
3.5. Root Systems Distribution in the Soil Profile
3.6. Grain Yield and Yield Components
4. Discussion
4.1. Water Stress Characterisation of the Early Season Drought Treatment
4.2. Early Season Drought Delayed Phenology in Wheat Cultivars with Different-Sized Root Systems
4.3. Small Root System had Significant Reduction in Leaf Area and Biomass at Anthesis
4.4. Early Season Drought Reduced the Root: Shoot Ratio at Anthesis
4.5. Early Season Drought Largely Reduced Grain Yield in a Cultivar with a Smaller Root System
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Asseng, S.; Pannell, D.J. Adapting dryland agriculture to climate change: Farming implications and research and development needs in Western Australia. Clim. Chang. 2013, 118, 167–181. [Google Scholar] [CrossRef]
- Fletcher, A.L.; Robertson, M.J.; Abrecht, D.G.; Sharma, D.L.; Holzworth, D.P. Dry sowing increases farm level wheat yields but not production risks in a mediterranean environment. Agric. Syst. 2015, 136, 114–124. [Google Scholar] [CrossRef]
- Chenu, K.; Deihimfard, R.; Chapman, S.C. Large-scale characterization of drought pattern: A continent-wide modelling approach applied to the Australian wheatbelt—Spatial and temporal trends. New Phytol. 2013, 198, 801–820. [Google Scholar] [CrossRef] [PubMed]
- Sprigg, H.; Belford, R.; Milroy, S.; Bennett, S.J.; Bowran, D. Adaptations for growing wheat in the drying climate of Western Australia. Crop Pasture Sci. 2014, 65, 627–644. [Google Scholar] [CrossRef]
- Cammarano, D.; Basso, B.; Stefanova, L.; Grace, P. Adapting wheat sowing dates to projected climate change in the Australian subtropics: Analysis of crop water use and yield. Crop Pasture Sci. 2012, 63, 974–986. [Google Scholar] [CrossRef]
- Adda, A.; Sahnoune, M.; Kaid-Harch, M.; Merah, O. Impact of water deficit intensity on durum wheat seminal roots. C. R. Biol. 2005, 328, 918–927. [Google Scholar] [CrossRef] [PubMed]
- French, B.; Palta, J.A. Early vigour avoids drought stress. GRDC Gr. Cover 2014, 112, 71–74. [Google Scholar]
- Armstrong, L.J.; Abrecht, D.J.; Anderson, W.K.; Belford, R.K. The effect of non-lethal water deficits during establishment on the growth of wheat crops. In Proceedings of the 8th Australian Agronomy Conference, Toowoomba, QLD, Australia, 30 January–2 February 1996; pp. 80–83. [Google Scholar]
- El Hafid, R.; Smith, D.H.; Karrou, M.; Samir, K. Physiological responses of spring durum wheat cultivars to early-season drought in a mediterranean environment. Ann. Bot. 1998, 81, 363–370. [Google Scholar] [CrossRef]
- El Hafid, R.; Smith, D.H.; Karrou, M.; Samir, K. Morphological attributes associated with early-season drought tolerance in spring durum wheat in a mediterranean environment. Euphytica 1998, 101, 273–282. [Google Scholar] [CrossRef]
- Han, H.; Tian, Z.; Fan, Y.; Cui, Y.; Cai, J.; Jiang, D.; Cao, W.; Dai, T. Water-deficit treatment followed by re-watering stimulates seminal root growth associated with hormone balance and photosynthesis in wheat (Triticum aestivum L.) seedlings. Plant Growth Regul. 2015, 77, 201–210. [Google Scholar] [CrossRef]
- Hamblin, A.; Tennant, D. Root length density and water uptake in cereals and grain legumes: How well are they correlated? Aust. J. Agric. Res. 1987, 38, 513–527. [Google Scholar] [CrossRef]
- Palta, J.A.; Chen, X.; Milroy, S.P.; Rebetzke, G.J.; Dreccer, M.F.; Watt, M. Large root systems: Are they useful in adapting wheat to dry environments? Funct. Plant Biol. 2011, 38, 347–354. [Google Scholar] [CrossRef]
- Středa, T.; Dostál, V.; Horáková, V.; Chloupek, O. Effective use of water by wheat varieties with different root system sizes in rain-fed experiments in central Europe. Agric. Water Manag. 2012, 104, 203–209. [Google Scholar] [CrossRef]
- Heřmanská, A.; Středa, T.; Chloupek, O. Improved wheat grain yield by a new method of root selection. Agron. Sustain. Dev. 2014, 35, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Abdolshahi, R.; Nazari, M.; Safarian, A.; Sadathossini, T.S.; Salarpour, M.; Amiri, H. Integrated selection criteria for drought tolerance in wheat (Triticum aestivum L.) breeding programs using discriminant analysis. Field Crops Res. 2015, 174, 20–29. [Google Scholar] [CrossRef]
- Aziz, M.M.; Palta, J.A.; Siddique, K.H.M.; Sadras, V.O. Five decades of selection for yield reduced root length density and increased nitrogen uptake per unit root length in Australian wheat varieties. Plant Soil 2016, 413, 181–192. [Google Scholar] [CrossRef]
- Cossani, C.M.; Sadras, V.O. Water-nitrogen co-limitation in grain crops. Adv. Agron. 2018, 150, 231–274. [Google Scholar]
- Isbell, R.F. A Classification System for Australian Soils (Third Approximation); Technical Report 2/1993; CSIRO: Canberra, Australia, 1993.
- Figueroa-Bustos, V.; Palta, J.; Chen, Y.; Siddique, K.H.M. Characterization of root and shoot traits in wheat cultivars with putative differences in root system size. Agronomy 2018, 8, 109. [Google Scholar] [CrossRef]
- Flower, K.C.; Cordingley, N.; Ward, P.R.; Weeks, C. Nitrogen, weed management and economics with cover crops in conservation agriculture in a mediterranean climate. Field Crops Res. 2012, 132, 63–75. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Austin, R.B. Prospects for genetically increasing the photosynthetic capacity of crops. In Perspectives in Biochemical and Genetic Regulation of Photosynthesis; Zelitch, I., Ed.; Alan R. Liss Inc.: New York, NY, USA, 1990. [Google Scholar]
- Turner, N.C. Measurement of plant water status by the pressure chamber technique. Irrig. Sci. 1988, 9, 289–308. [Google Scholar] [CrossRef]
- Palta, J.A.; Fillery, I.R. Nitrogen accumulation and remobilisation in wheat of 15n-urea applied to a duplex soil at seeding. Aust. J. Exp. Agric. 1992, 32, 995. [Google Scholar] [CrossRef]
- Chen, Y.; Ghanem, M.; Siddique, K.H.M. Characterising root trait variability in chickpea (Cicer arietinum L.) germplasm. J. Exp. Bot. 2016, 68, 1987–1999. [Google Scholar]
- Liao, M.; Palta, J.A.; Fillery, I.R.P. Root characteristics of vigorous wheat improve early nitrogen uptake. Aust. J. Agric. Res. 2006, 57, 1097–1107. [Google Scholar] [CrossRef]
- Henson, I.E.; Jensen, C.R.; Turner, N.C. Leaf gas exchange and water relations of lupins and wheat. I. Shoot responses to soil water deficits. Aust. J. Plant Physiol. 1989, 16, 401–413. [Google Scholar] [CrossRef]
- Jensen, C.R.; Henson, I.E.; Turner, N.C. Leaf gas exchange and water relations of lupins and wheat. II. Root and shoot water relations of lupin during drought-induced stomatal closure. Aust. J. Plant Physiol. 1989, 16, 415–428. [Google Scholar] [CrossRef]
- Angus, J.F.; Moncur, M.W. Water stress and phenology in wheat. Aust. J. Agric. Res. 1977, 28, 177–181. [Google Scholar] [CrossRef]
- Motzo, R.; Attene, G.; Deidda, M. Genotypic variation in durum wheat root systems at different stages of development in a mediterranean environment. Euphytica 1993, 66, 197–206. [Google Scholar] [CrossRef]
- Gregory, P.J.; Atwell, B.J. The fate of carbon in pulse-labelled crops of barley and wheat. Plant Soil 1991, 136, 205–213. [Google Scholar] [CrossRef]
- Palta, J.A.; Gregory, P.J. Drought affects the fluxes of carbon to roots and soil in 13c pulse-labelled plants of wheat. Soil Biol. Biochem. 1997, 29, 1395–1403. [Google Scholar] [CrossRef]
- Siddique, K.H.M.; Belford, R.K.; Tennant, D. Root: Shoot ratios of old and modern, tall and semi-dwarf wheats in a Mediterranean environment. Plant Soil 1990, 121, 89–98. [Google Scholar] [CrossRef]
- Kobata, T.; Palta, J.A.; Turner, N.C. Rate of development of postanthesis water deficits and grain filling of spring wheat. Crop Sci. 1992, 32, 1238–1242. [Google Scholar] [CrossRef]
- Palta, J.; Kobata, T.; Turner, N.C.; Fillery, I.R. Remobilization of carbon and nitrogen in wheat as influenced by postanthesis water deficits. Crop Sci. 1994, 34, 118–124. [Google Scholar] [CrossRef]
- Allard, V.; Martre, P.; Le Gouis, J. Genetic variability in biomass allocation to roots in wheat is mainly related to crop tillering dynamics and nitrogen status. Eur. J. Agron. 2013, 46, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Hamblin, A.; Tennant, D.; Perry, M.W. The cost of stress: Dry matter partitioning changes with seasonal supply of water and nitrogen to dryland wheat. Plant Soil 1990, 122, 47–58. [Google Scholar] [CrossRef]
- Asseng, S.; Ritchie, J.T.; Smucker, A.J.M.; Robertson, M.J. Root growth and water uptake during water deficit and recovering in wheat. Plant Soil 1998, 201, 265–273. [Google Scholar] [CrossRef]
- El Hafid, R.; Smith, D.H.; Karrou, M.; Samir, K. Root and shoot growth, water use and water use efficiency of spring durum wheat under early-season drought. Agronomie 1998, 18, 181–195. [Google Scholar] [CrossRef] [Green Version]
- van Ginkel, M.; Calhoun, D.S.; Gebeyehu, G.; Miranda, A.; Tian-you, C.; Pargas Lara, R.; Trethowan, R.M.; Sayre, K.; Crossa, J.; Rajaram, S. Plant traits related to yield of wheat in early, late, or continuous drought conditions. Euphytica 1998, 100, 109–121. [Google Scholar] [CrossRef]
- Asseng, S.; van Herwaarden, A.F. Analysis of the benefits to wheat yield from assimilates stored prior to grain filling in a range of environments. Plant Soil 2003, 256, 217–229. [Google Scholar] [CrossRef]
- Golabadi, M.; Arzani, A.; Mirmohammadi Maibody, S.A.M.; Sayed Tabatabaei, B.E.; Mohammadi, S.A. Identification of microsatellite markers linked with yield components under drought stress at terminal growth stages in durum wheat. Euphytica 2010, 177, 207–221. [Google Scholar] [CrossRef]
- Senapati, N.; Stratonovitch, P.; Paul, M.J.; Semenov, M.A. Drought tolerance during reproductive development is important for increasing wheat yield potential under climate change in europe. J. Exp. Bot. 2018, 70, 2549–2560. [Google Scholar] [CrossRef]
- Siddique, K.H.M.; Tennant, D.; Perry, M.W.; Belford, R.K. Water use and water use efficiency of old and modern wheat cultivars in a mediterranean-type environment. Aust. J. Exp. Agric. 1990, 41, 431–447. [Google Scholar] [CrossRef]
- Lopes, M.S.; Reynolds, M.P.; Manes, Y.; Singh, R.P.; Crossa, J.; Braun, H.J. Genetic yield gains and changes in associated traits of cimmyt spring bread wheat in a “historic” set representing 30 years of breeding. Crop Sci. 2012, 52, 1123–1131. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Kurishbayev, A.; Jatayev, S.; Shvidchenko, V.; Zotova, L.; Koekemoer, F.; de Groot, S.; Soole, K.; Langridge, P. Early flowering as a drought escape mechanism in plants: How can it aid wheat production? Front. Plant. Sci. 2017, 8, 1–8. [Google Scholar] [CrossRef]
- Siddique, K.H.M.; Belford, R.K.; Perry, M.W.; Tennant, D. Growth, development and light interception of old and modern wheat cultivars in a mediterranean-type environment. Aust. J. Exp. Agric. 1989, 40, 473–487. [Google Scholar]
- French, B.; Abrecht, D.G. Wheat Genotypes Differ in Response to Early Water Deficit; 2013 WA Crop Updates; GRDC: Canberra, Australia, 2013. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Booting | Anthesis | Physiological Maturity | Duration of the Grain Filling | ||||
|---|---|---|---|---|---|---|---|---|
| (DAS) | (DAS) | (DAS) | Days | |||||
| WW | ESD | WW | ESD | WW | ESD | WW | ESD | |
| Bahatans-87 | 93c | 100b | 102c | 109b | 139c | 150b | 37c | 41c |
| Harper | 100b | 110a | 107b | 120a | 170a | 174a | 63a | 54b |
| Tincurrin | 74e | 81d | 87e | 94d | 137c | 147b | 50b | 53b |
| LSD (C) LSD (T) LSD (CxT) | 2.3 1.9 3.3** | 2.9 2.4 4.1** | 6.1*** 5** NS | 4.1 3.4 5.8*** | ||||
| Cultivar | Leaf Area (cm2plant−1) | Specific Leaf Area (cm2 g −1) | Tiller Number (plant−1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 30 DAS | Anthesis | 30 DAS | Anthesis | 30 DAS | Anthesis | |||||||
| WW | ESD | WW | ESD | WW | ESD | WW | ESD | WW | ESD | WW | ESD | |
| Bahatans-87 | 101a | 20c | 1946b | 1648c | 270b | 159d | 229c | 215bc | 3a | 0d | 17a | 14b |
| Harper | 75b | 24c | 2604a | 2444a | 272b | 212c | 245b | 225b | 2b | 0d | 18a | 16b |
| Tincurrin | 77b | 24c | 1548c | 1194d | 307a | 219c | 305a | 287a | 1c | 0d | 8c | 7c |
| LSD (C) | 8.4 | 179.7 | 17.6 | 13.2 | 0.2 | 1.2*** | ||||||
| LSD (T) | 6.9 | 146.7 | 14.4 | 10.7 | 0.2 | 1.0** | ||||||
| LSD (CxT) | 11.9** | 254.1* | 24.9* | 18.6*** | 0.3*** | NS | ||||||
| Cultivar | Grain Yield | Spikes | Grains | 1000 Grain Weight | Harvest Index | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| (g plant−1) | (plant−1) | (spike−1) | (g) | |||||||
| WW | ESD | WW | ESD | WW | ESD | WW | ESD | WW | ESD | |
| Bahatans-87 | 31a | 28b | 17b | 16b | 42c | 40c | 44a | 45a | 0.38c | 0.35d |
| Harper | 26c | 23d | 21a | 16b | 44bc | 40c | 32c | 33c | 0.35d | 0.34d |
| Tincurrin | 32a | 27bc | 14c | 14c | 55a | 46b | 41b | 40b | 0.52a | 0.46b |
| LSD (C) | 2.4*** | 1.3 | 3.7 | 1.6** | 0.03*** | |||||
| LSD (T) | 1.9*** | 1.1 | 3.0 | NS | 0.03* | |||||
| LSD (CxT) | NS | 1.8** | 5.2* | NS | NS | |||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figueroa-Bustos, V.; Palta, J.A.; Chen, Y.; Siddique, K.H.M. Early Season Drought Largely Reduces Grain Yield in Wheat Cultivars with Smaller Root Systems. Plants 2019, 8, 305. https://doi.org/10.3390/plants8090305
Figueroa-Bustos V, Palta JA, Chen Y, Siddique KHM. Early Season Drought Largely Reduces Grain Yield in Wheat Cultivars with Smaller Root Systems. Plants. 2019; 8(9):305. https://doi.org/10.3390/plants8090305
Chicago/Turabian StyleFigueroa-Bustos, Victoria, Jairo A. Palta, Yinglong Chen, and Kadambot H.M. Siddique. 2019. "Early Season Drought Largely Reduces Grain Yield in Wheat Cultivars with Smaller Root Systems" Plants 8, no. 9: 305. https://doi.org/10.3390/plants8090305
APA StyleFigueroa-Bustos, V., Palta, J. A., Chen, Y., & Siddique, K. H. M. (2019). Early Season Drought Largely Reduces Grain Yield in Wheat Cultivars with Smaller Root Systems. Plants, 8(9), 305. https://doi.org/10.3390/plants8090305