Inflorescence Development and Floral Organogenesis in Taraxacum kok-saghyz

 ,
, 
 and
and
Abstract
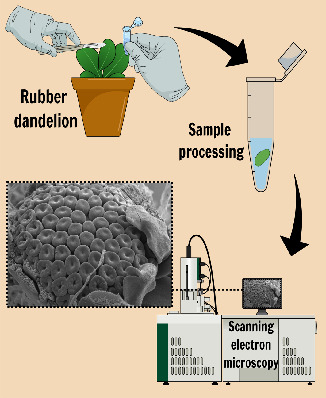
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Morphological Aspects
2.2. Inflorescence Developmental Stages
2.3. Floral Organogenesis
2.4. Timing of Inflorescence and Floral Organogenesis Correlated to Plant Development
2.5. Inflorescence Bud Appearance and Anthesis
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Morphological Evaluation
4.3. Inflorescence Development and Floral Organogenesis
4.4. Timing of Inflorescence Bud Appearance and Anthesis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kilian, N.; Gemeinholzer, B.; Lack, H.W. Cichorieae. In Systematics, Evolution, and Biogeography of Compositae; Funk, V.A., Susanna, A., Stuessy, T.F., Bayer, R.J., Eds.; International Association for Plant Taxonomy: Vienna, Austria, 2009; pp. 343–383. ISBN 9783950175431. [Google Scholar]
- Funk, V.A.; Susanna, A.; Stuessy, T.F.; Robinson, H. Classification of Compositae. In Systematics, Evolution, and Biogeography of Compositae; Funk, V.A., Susanna, A., Stuessy, T.F., Bayer, R.J., Eds.; International Association for Plant Taxonomy: Vienna, Austria, 2009; pp. 171–189. [Google Scholar]
- Musiał, K.; Płachno, B.J.; Świątek, P.; Marciniuk, J. Anatomy of ovary and ovule in dandelions (Taraxacum, Asteraceae). Protoplasma 2013, 250, 715–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musiał, K.; Górka, P.; Kościńska-Pajak, M.; Marciniuk, P. Embryological studies in Taraxacum udum Jordan (sect. Palustria). Botany 2013, 91, 614–620. [Google Scholar] [CrossRef]
- Van Baarlen, P.; van Dijk, P.J.; Hoekstra, R.F.; Jong, J.H.D. Meiotic recombination in sexual diploid and apomictic triploid dandelions (Taraxacum officinale L.). Genome 2000, 43, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Arias, M.; Herrero, J.; Ricobaraza, M.; Hernández, M.; Ritter, E. Evaluation of root biomass, rubber and inulincontents in nine Taraxacum koksaghyz Rodin populations. Ind. Crop. Prod. 2016, 83, 316–321. [Google Scholar] [CrossRef]
- Kirschner, J.; Štěpánek, J.; Černý, T.; De Heer, P.; van Dijk, P.J. Available ex situ germplasm of the potential rubber crop Taraxacum koksaghyz belongs to a poor rubber producer, T. brevicorniculatum (Compositae–Crepidinae). Genet. Resour. Crop. Evol. 2013, 60, 455–471. [Google Scholar] [CrossRef]
- Van Dijk, P.; Kirschner, J.; Štěpánek, J.; Omarovich Baitulin, I.; Černý, T. Taraxacum koksaghyz Rodin definitely is not an example of overcollecting in the past. A reply to S. Volis et al. (2009). J. Appl. Bot. Food Qual. 2010, 83, 217–219. [Google Scholar]
- Krotkov, G. A review of literature on Taraxacum koksaghyz Rod. Bot. Rev. 1945, 11, 417–461. [Google Scholar] [CrossRef]
- Mooibroek, H.; Cornish, K. Alternative sources of natural rubber. Appl. Microbiol. Biotechnol. 2000, 53, 355–365. [Google Scholar] [CrossRef]
- Zhang, N.; Guo, T.; Ma, X.; Liu, J.; Dong, Y.; Zhang, J. Rational rubber extraction and simultaneous determination of rubber content and molecular weight distribution in Taraxacum kok-saghyz Rodin by size-exclusion chromatography. Chromatographia 2019, 82, 1459–1466. [Google Scholar] [CrossRef]
- Bates, G.M.; McNulty, S.K.; Amstutz, N.D.; Pool, V.K.; Cornish, K. Planting density and growth cycle affect actual and potential latex and rubber yields in Taraxacum kok-saghyz. HortScience 2019, 54, 1338–1344. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Cadavid, D.A.; Cornish, K.; Hathwaik, U.; Kozak, R.; McMahan, C.; Michel, F.C. Development of novel processes for the aqueous extraction of natural rubber from Taraxacum kok-saghyz (TK). J. Chem. Technol. Biotechnol. 2019, 94, 2452–2464. [Google Scholar] [CrossRef]
- Ramirez-Cadavid, D.A.; Valles-Ramirez, S.; Cornish, K.; Michel, F.C. Simultaneous quantification of rubber, inulin, and resins in Taraxacum kok-saghyz (TK) roots by sequential solvent extraction. Ind. Crop. Prod. 2018, 122, 647–656. [Google Scholar] [CrossRef]
- Collins-Silva, J.; Nural, A.T.; Skaggs, A.; Scott, D.; Hathwaik, U.; Woolsey, R.; Schegg, K.; McMahan, C.; Whalen, M.; Cornish, K.; et al. Altered levels of the Taraxacum kok-saghyz (Russian dandelion) small rubber particle protein, TkSRPP3, result in qualitative and quantitative changes in rubber metabolism. Phytochemistry 2012, 79, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Van Beilen, J.B.; Poirier, Y. Establishment of new crops for the production of natural rubber. Trends Biotechnol. 2007, 25, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Van Beilen, J.B.; Poirier, Y. Guayule and Russian dandelion as alternative sources of natural rubber. Crit. Rev. Biotechnol. 2007, 27, 217–231. [Google Scholar] [CrossRef]
- Junkong, P.; Cornish, K.; Ikeda, Y. Characteristics of mechanical properties of sulphur cross-linked guayule and dandelion natural rubbers. RSC Adv. 2017, 7, 50739–50752. [Google Scholar] [CrossRef] [Green Version]
- Cherian, S.; Ryu, S.B.; Cornish, K. Natural rubber biosynthesis in plants, the rubber transferase complex, and metabolic engineering progress and prospects. Plant Biotechnol. J. 2019, 17, 2041–2061. [Google Scholar] [CrossRef] [Green Version]
- Keener, H.M.; Shah, A.; Klingman, M.; Wolfe, S.; Pote, D.; Fioritto, R. Progress in direct seeding of an alternative natural rubber plant, Taraxacum kok-saghyz (L.E. Rodin). Agronomy 2018, 8, 182. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Cadavid, D.A.; Cornish, K.; Michel, F.C. Taraxacum kok-saghyz (TK): Compositional analysis of a feedstock for natural rubber and other bioproducts. Ind. Crop. Prod. 2017, 107, 624–640. [Google Scholar] [CrossRef]
- Hodgson-Kratky, K.J.M.; Stoffyn, O.M.; Wolyn, D.J. Recurrent selection for rubber yield in Russian dandelion. J. Am. Soc. Hortic. Sci. 2017, 142, 470–475. [Google Scholar] [CrossRef]
- Cornish, K.; Kopicky, S.L.; Mcnulty, S.K.; Amstutz, N.; Chanon, A.M.; Walker, S.; Kleinhenz, M.D.; Miller, A.R.; Streeter, J.G. Temporal diversity of Taraxacum kok-saghyz plants reveals high rubber yield phenotypes. Biodiversitas J. Biol. Divers. 2016, 17, 847–856. [Google Scholar] [CrossRef]
- Luo, Z.; Iaffaldano, B.J.; Zhuang, X.; Fresnedo-Ramírez, J.; Cornish, K. Variance, Inter-Trait Correlation, Heritability, and Marker-Trait Association of Rubber Yield-Related Characteristics in Taraxacum kok-saghyz. Plant Mol. Biol. Rep. 2018, 36, 576–587. [Google Scholar] [CrossRef]
- Luo, Z.; Iaffaldano, B.J.; Zhuang, X.; Fresnedo-Ramírez, J.; Cornish, K. Analysis of the first Taraxacum kok-saghyz transcriptome reveals potential rubber yield related SNPs. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iaffaldano, B.; Zhang, Y.; Cornish, K. CRISPR/Cas9 genome editing of rubber producing dandelion Taraxacum kok-saghyz using Agrobacterium rhizogenes without selection. Ind. Crop. Prod. 2016, 89, 356–362. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Iaffaldano, B.J.; Cornish, K. Colchicine-induced polyploidy has the potential to improve rubber yield in Taraxacum kok-saghyz. Ind. Crop. Prod. 2018, 112, 75–81. [Google Scholar] [CrossRef]
- McAssey, E.V.; Gudger, E.G.; Zuellig, M.P.; Burke, J.M. Population Genetics of the Rubber-Producing Russian Dandelion (Taraxacum kok-saghyz). PLoS ONE 2016, 11, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Iaffaldano, B.J.; Xie, W.; Blakeslee, J.J.; Cornish, K. Rapid and hormone-free Agrobacterium rhizogenes-mediated transformation in rubber producing dandelions Taraxacum kok-saghyz and T. brevicorniculatum. Ind. Crop. Prod. 2015, 66, 110–118. [Google Scholar] [CrossRef]
- Kreuzberger, M.; Hahn, T.; Zibek, S.; Schiemann, J.; Thiele, K. Seasonal pattern of biomass and rubber and inulin of wild Russian dandelion (Taraxacum koksaghyz L. Rodin) under experimental field conditions. Eur. J. Agron. 2016, 80, 66–77. [Google Scholar] [CrossRef]
- Hodgson-Kratky, K.J.M.; Demers, M.N.K.; Stoffyn, O.M.; Wolyn, D.J. Harvest date, post-harvest vernalization and regrowth temperature affect flower bud induction in Russian dandelion (Taraxacum kok-saghyz). Can. J. Plant Sci. 2015, 95, 1221–1228. [Google Scholar] [CrossRef]
- Hodgson-Kratky, K.J.M.; Wolyn, D.J. Inheritance of flowering habit in Russian dandelion. J. Am. Soc. Hortic. Sci. 2015, 140, 614–619. [Google Scholar] [CrossRef] [Green Version]
- Whaley, G.W.; Bowen, J.S. Russian Dandelion (Kok-Saghyz) an Emergency Source of Natural Rubber; USDA Miscellaneous Publication: Washington, DC, USA, 1947.
- Xiao, H.; Radovich, C.; Welty, N.; Hsu, J.; Li, D.; Meulia, T.; van der Knaap, E. Integration of tomato reproductive developmental landmarks and expression profiles, and the effect of SUN on fruit shape. BMC Plant Biol. 2009, 9, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibañez, M.S.; Mercado, M.I.; Coll Aráoz, M.V.; Zannier, M.L.; Grau, A.; Ponessa, G.I. Flower structure and developmental stages of the capitulum of Smallanthus sonchifolius (Asteraceae): Reproductive implications. J. Plant Res. 2017, 130, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Berti, F.; Fambrini, M.; Turi, M.; Bertini, D.; Pugliesi, C. Mutations of corolla symmetry affect carpel and stamen development in Helianthus. Annu. Can. J. Bot. 2005, 83, 1065–1072. [Google Scholar] [CrossRef]
- Tahtiharju, S.; Rijpkema, A.S.; Vetterli, A.; Albert, V.A.; Teeri, T.H.; Elomaa, P. Evolution and diversification of the CYC/TB1 gene family in Asteraceae-A comparative study in Gerbera (Mutisieae) and Sunflower (Heliantheae). Mol. Biol. Evol. 2012, 29, 1155–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laitinen, R.A.; Broholm, S.; Albert, V.A.; Teeri, T.H.; Elomaa, P. Patterns of MADS-box gene expression mark flower-type development in Gerbera hybrida (Asteraceae). BMC Plant Biol. 2006, 6, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Harris, E.M. Inflorescence and floral ontogeny in Asteraceae: A synthesis of historical and current concepts. Bot. Rev. 1995, 61, 93–278. [Google Scholar] [CrossRef]
- Dadpour, M.R.; Naghiloo, S.; Neycharan, S.F. The development of pistillate and perfect florets in Xeranthemum squarrosum (Asteraceae). Plant Biol. 2012, 14, 234–243. [Google Scholar] [CrossRef]
- Ren, J.B.; Guo, Y.P. Behind the diversity: Ontogenies of radiate, disciform, and discoid capitula of Chrysanthemum and its allies. J. Syst. Evol. 2015, 53, 520–528. [Google Scholar] [CrossRef]
- Yu, D.; Kotilainen, M.; Pöllänen, E.; Mehto, M.; Elomaa, P.; Helariutta, Y.; Albert, V.A.; Teeri, T.H. Organ identity genes and modified patterns of flower development in Gerbera hybrida (Asteraceae). Plant J. 1999, 17, 51–62. [Google Scholar] [CrossRef]
- Teeri, T.H.; Kotilainen, M.; Uimari, A.; Ruokolainen, S.; Peng Ng, Y.; Malm, U.; Pöllänen, E.; Broholm, S.; Laitinen, R.; Elomaa, P.; et al. Floral developmental genetics of Gerbera (Asteraceae). In Advances in Botanical Research-Developmental Genetics of the Flower; Soltis, D.E., LeebensMack, J.H., Soltis, P.S., Callow, J.A., Eds.; Academic Press: Massachusetts, MA, USA, 2006; Volume 44, pp. 323–351. ISBN 0065-2296. [Google Scholar]
- Dadpour, M.R.; Naghiloo, S.; Gohari, G. Inflorescence and floral ontogeny in Osteospermum ecklonis (Asteraceae). Botany 2011, 89, 605–614. [Google Scholar] [CrossRef]
- Oraei, M.; Gohari, G.; Esmaillou, Z.; Naghiloo, S. Comparative ontogeny of perfect and pistillate florets in Senecio vernalis (Asteraceae). Flora-Morphol. Distrib. Funct. Ecol. Plants 2013, 208, 285–292. [Google Scholar] [CrossRef]
- Elomaa, P.; Zhao, Y.; Zhang, T. Flower heads in Asteraceae—Recruitment of conserved developmental regulators to control the flower-like inflorescence architecture. Hortic. Res. 2018, 5, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffrey, C. Evolution of Compositae flowers. In Systematics, Evolution, and Biogeography of Compositae; Funk, V.A., Susanna, A., Stuessy, T.F., Bayer, R.J., Eds.; International Association for Plant Taxonomy: Vienna, Austria, 2009; pp. 131–138. [Google Scholar]
- Endress, P.K. Evolutionary diversification of the flowers in angiosperms. Am. J. Bot. 2011, 98, 370–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coulter, J.M. Development of a dandelion flower. Am. Nat. 1883, 17, 1211–1217. [Google Scholar] [CrossRef]
- Collier, M.H.; Rogstad, S.H. Clonal variation in floral stage timing in the common dandelion Taraxacum officinale (Asteraceae). Am. J. Bot. 2004, 91, 1828–1833. [Google Scholar] [CrossRef]
- Ruzin, S.E. Plant Microtechnique and Microscopy; Oxford University Press: New York, NY, USA, 1999; ISBN 0195089561. [Google Scholar]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schuchovski, C.; Meulia, T.; Sant’Anna-Santos, B.F.; Fresnedo-Ramírez, J. Inflorescence Development and Floral Organogenesis in Taraxacum kok-saghyz. Plants 2020, 9, 1258. https://doi.org/10.3390/plants9101258
Schuchovski C, Meulia T, Sant’Anna-Santos BF, Fresnedo-Ramírez J. Inflorescence Development and Floral Organogenesis in Taraxacum kok-saghyz. Plants. 2020; 9(10):1258. https://doi.org/10.3390/plants9101258
Chicago/Turabian StyleSchuchovski, Carolina, Tea Meulia, Bruno Francisco Sant’Anna-Santos, and Jonathan Fresnedo-Ramírez. 2020. "Inflorescence Development and Floral Organogenesis in Taraxacum kok-saghyz" Plants 9, no. 10: 1258. https://doi.org/10.3390/plants9101258
APA StyleSchuchovski, C., Meulia, T., Sant’Anna-Santos, B. F., & Fresnedo-Ramírez, J. (2020). Inflorescence Development and Floral Organogenesis in Taraxacum kok-saghyz. Plants, 9(10), 1258. https://doi.org/10.3390/plants9101258