
Ongoing Evolution in the Genus Crocus: Diversity of Flowering Strategies on the Way to Hysteranthy

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Flowering of Spanish Crocuses in Their Natural Environment
2.2. Phenology of Spanish Crocuses Grown Ex Situ
2.3. Temperature Requirements for Flower Initiation and Differentiation during Corm Storage
2.4. Effect of Growth Temperatures on Flowering Time
2.5. Statistical Analysis
3. Results and Discussion
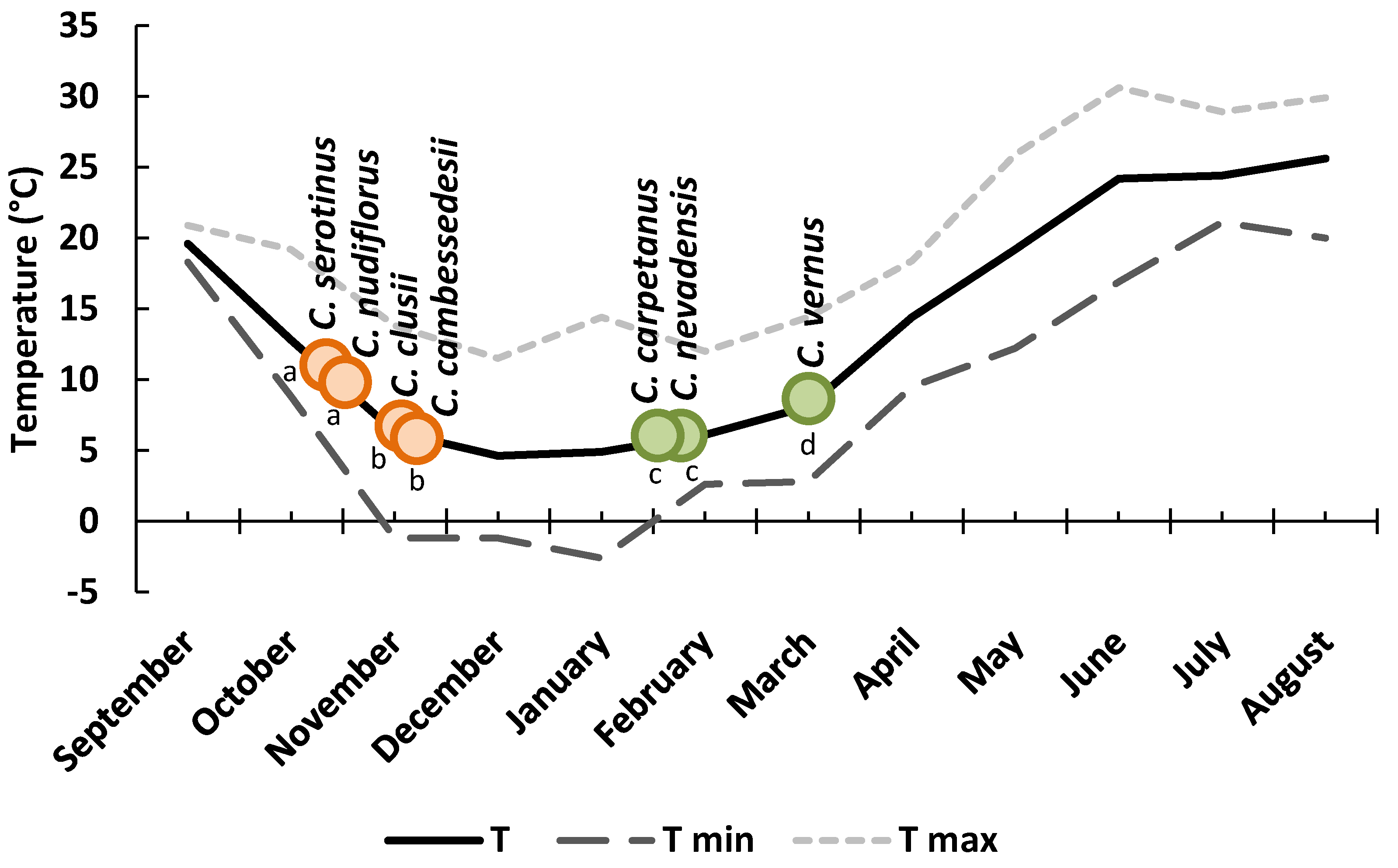
3.1. Phenology and Flowering Patterns in Natural Habitats
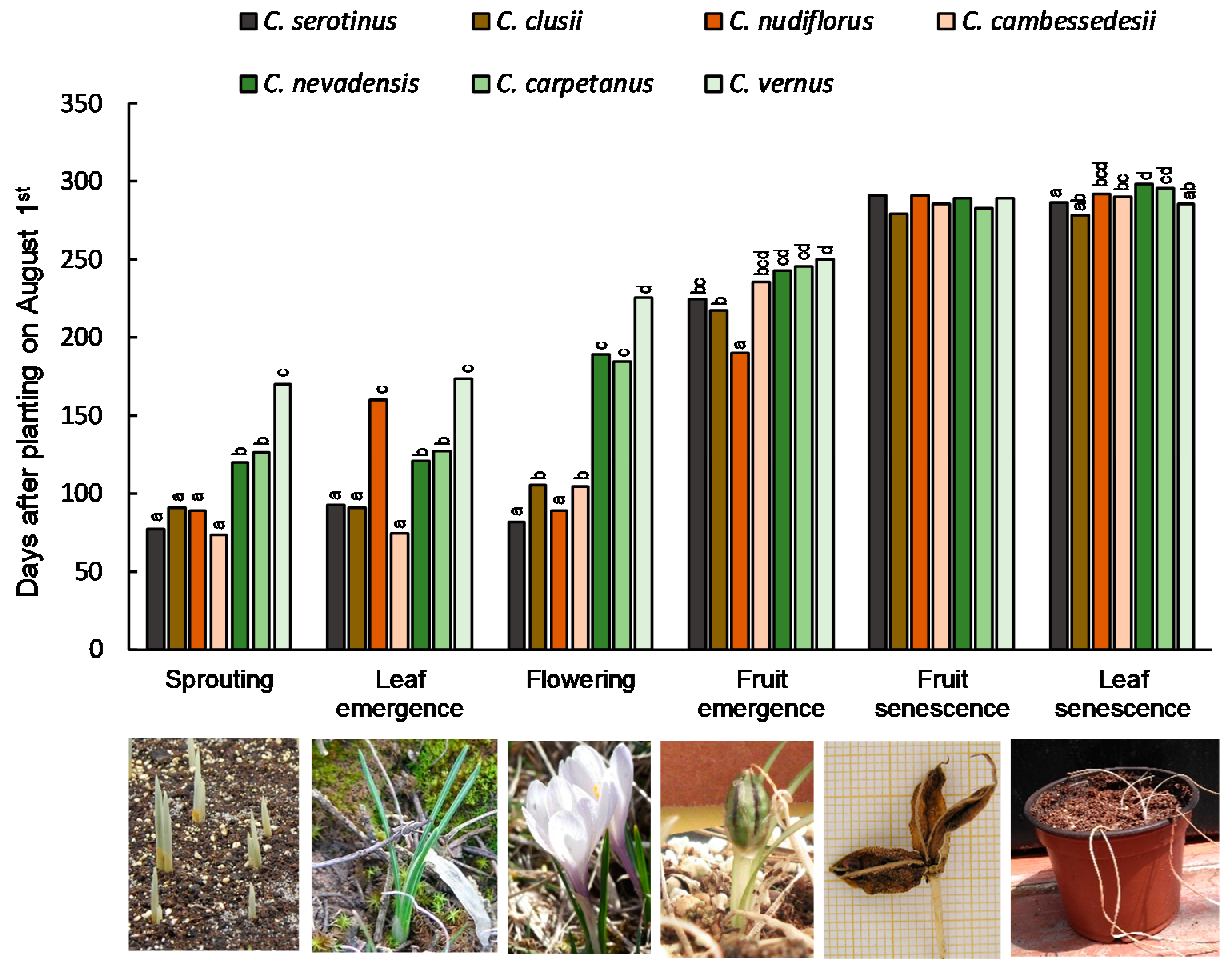
3.2. Phenology and Flowering in Common Garden Experiments Ex Situ
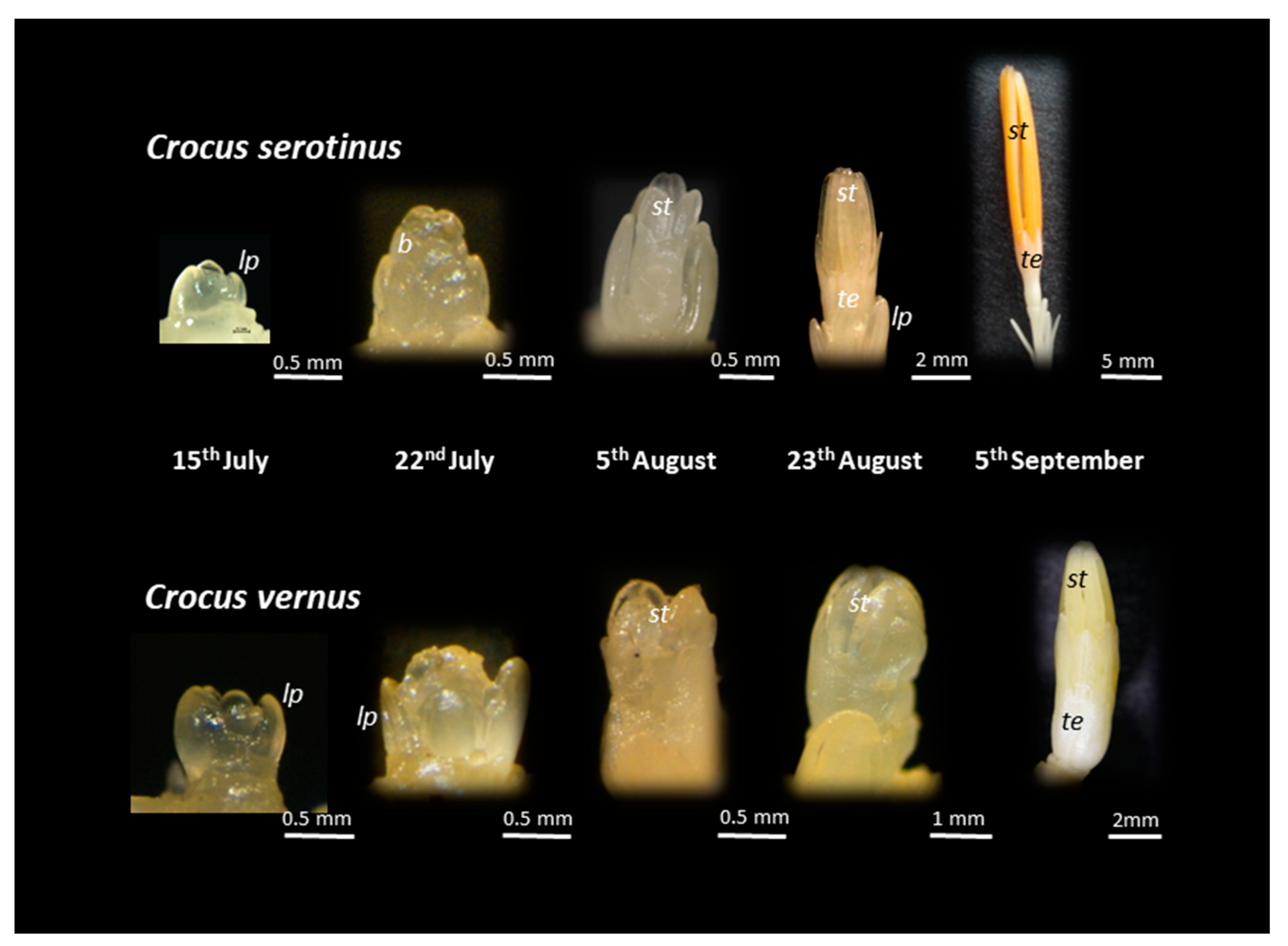
3.3. Temperature Requirements for Flower Initiation
3.4. Effect of Growth Temperatures on Flowering
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kamenetsky, R. Biodiversity of Geophytes. Phytogeography, Morphology and Survival Strategies. In Ornamental Geophytes. From Basic Science to Sustainable Production; Kamenetsky, K., Okubo, H., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 57–75. [Google Scholar]
- Guo, L.; Luedeling, E.; Dai, J.-H.; Xu, J.C. Differences in heat requirements of flower and leaf buds make hysteranthous trees bloom before leaf unfolding. Plant Divers. Resour. 2014, 36, 245–253. [Google Scholar]
- Dafni, A.; Cohen, D.; Noy-Meir, I. Life-cycle variation in geophytes. Ann. Mo. Bot. Gard. 1981, 68, 652–660. [Google Scholar] [CrossRef]
- Dafni, A.; Shmida, A.; Avishai, M. Leafless autumnal flowering geophytes in the Mediterranean region; Phytogeographical, ecological and evolutionary aspects. Plant Syst. Evol. 1981, 137, 181–193. [Google Scholar] [CrossRef]
- Shmida, A. The Tragacantic Alpine Vegetation of Mt. Hermon. Ph.D. Thesis, The Hebrew University of Jerusalem, Jerusalem, Israel, 1977. [Google Scholar]
- McNeil, J. Urginea Staeinh. Scilla. In Flora Europea; Tutin, T.G., Heywood, V.H., Burgues, N.A., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1980; Volume 5, pp. 40–43. [Google Scholar]
- Negbi, M. Theophastus on geophytes. Bot. J. Linn. Soc. 1989, 100, 15–43. [Google Scholar] [CrossRef]
- Rukšāns, J. Crocuses: A Complete Guide to the Genus, 1st ed.; Timber Press Inc.: Portland, OR, USA, 2010; pp. 53–196. [Google Scholar]
- Mathew, B. The Crocus. A Revision of the Genus Crocus (Iridaceae); Timber Press Inc.: Portland, OR, USA, 1982; pp. 27–117. [Google Scholar]
- Pérez, E.; Pastor, J. Contribución al estudio cariológico de la familia Iridaceae en Andalucía occidental. Lagascalia 1994, 17, 257–272. [Google Scholar]
- Sanz, G.M.; Redó, V.J.A. Nuevos datos sobre la flora de la provincia de Cuenca VI. Flora Montibérica 1998, 9, 28–36. [Google Scholar]
- Benschop, M. Crocus. In The Physiology of Flower Bulbs; De Hertog, A., Le Nard, M., Eds.; Elsevier Science Publishers: Amsterdam, The Netherlands, 1993; pp. 257–283. [Google Scholar]
- Chryssanthi, D.G.; Lamari, F.N.; Iatrou, G.; Pylara, A.; Karamanos, N.K.; Cordopatis, P. Inhibition of breast cancer cell proliferation by style constituents of different Crocus species. Anticancer Res. 2007, 27, 357–362. [Google Scholar]
- Molina, R.V.; Valero, M.; Navarro, Y.; García-Luis, A.; Guardiola, J.L. The effect of time of corm lifting and duration of incubation at inductive temperature on flowering in the saffron plant (Crocus sativus L.). Sci. Hortic. 2004, 103, 79–91. [Google Scholar] [CrossRef]
- Molina, R.V.; Valero, M.; Navarro, Y.; Garcia-Luis, A.; Guardiola, J.L. Low temperature storage of corms extends the flowering season of saffron (Crocus sativus L.). J. Hortic. Sci. Biotechnol. 2005, 80, 319–326. [Google Scholar] [CrossRef]
- Molina, R.V.; Valero, M.; Navarro, Y.; Guardiola, J.L.; García-Luis, A. Temperature effects on flower formation in saffron (Crocus sativus L.). Sci. Hortic. 2005, 103, 361–379. [Google Scholar] [CrossRef]
- De Los Mozos-Pascual, M.; Santana-Méridas, O.; Rodríguez-Conde, M.F.; Pastor-Férriz, T.; Fernández, J.A. A preliminary characterization of saffron germplasm from the CROCUSBANK collection. ISHS. Acta Hortic. 2010, 850, 35–40. [Google Scholar] [CrossRef]
- Pastor-Férriz, T. Caracterización Fenológica, Morfológica, Fisiológica, Fitoquímica y Molecular de las Especies Silvestres Españolas del Género Crocus. Ph.D. Thesis, Universitat Politècnica de València, Valencia, Spain, 2015. [Google Scholar]
- Fernández, J.A.; Santana, O.; Guardiola, J.L.; Molina, R.V.; Heslop-Harrison, P.; Borbely, G.; Branca, F.; Argento, S.; Maloupa, E.; Talou, T.; et al. The World Saffron and Crocus collection: Strategies for establishment, management, characterisation and utilisation. Genet. Resour. Crop. Evol. 2011, 58, 125–137. [Google Scholar] [CrossRef] [Green Version]
- Engels, J.M.M.; Visser, L. A Guide to Effective Management of Germplasm Collections. IPGRI Handbooks for Genebanks No 6; IPGRI: Rome, Italy, 2003. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circular. Calif. Agric. Exp. Stn. 1950, 347, 1–32. [Google Scholar]
- Guillén, A.; Castroviejo, S.; Rico, E.; Crespo, M.B.; Quintanar, A.; Herrero, A.; Aedo, C. Crocus L. Flora Ibérica. In Real Jardín Botánico; CSIC: Madrid, Spain, 2014; Volume 20, pp. 454–464. [Google Scholar]
- Benito, A.J.L. Base de Datos de la Flora y Vegetación del Parque Nacional de Ordesa y Monte Perdido. (Pirineo aragonés), 2nd ed.; JOLUBE Consultor y Editor Ambiental: Huesca, Spain, 2010; p. 386. [Google Scholar]
- Hervás, J.L.; Fernández, C. Algunas monocotiledóneas del Alto Guadalquivir (sur de la península Ibérica) hasta 1.999. Blancoana 2000, 17, 71–76. [Google Scholar]
- Shmida, A.; Dafni, A. Blooming strategies, flower size and advertisement in the “Lily Group” geophytes of Israel. Herbertia 1989, 45, 111–123. [Google Scholar]
- Dafni, A. Autumnal and winter adaptations under Mediterranean conditions. Bocconea 1996, 5, 171–181. [Google Scholar]
- Kamenetsky, R. Patterns of dormancy and florogenesis in herbaceous perennial plants: Environmental and internal regulation. Crop Sci. 2009, 49, 2400–2404. [Google Scholar] [CrossRef]
- Badri, M.A.; Minchin, P.E.H.; Lapointe, L. Effects of temperature on the growth of spring ephemerals: Crocus vernus. Physiol. Plant. 2007, 130, 67–76. [Google Scholar] [CrossRef]
- Balaguer, L.; Martínez-Ferri, E.; Valladares, F.; Pérez-Corona, M.E.; Baquedano, F.J.; Castillo, F.J.; Manrique, E. Population divergence in the plasticity of the response of Quercus coccifera to the light environment. Funct. Ecol. 2001, 15, 124–135. [Google Scholar] [CrossRef] [Green Version]
- Valladares, F.; Gianoli, E.; Gómez, J.M. Ecological limits to plant phenotypic plasticity. New Phytol. 2007, 176, 749–763. [Google Scholar] [CrossRef]
- De los Mozos, M. Personal Communication; Departamento de Gestión y Conservación de Recursos Fitogenéticos; Centro de Investigación Agroforestal de Albaladejito: Cuenca, Spain, 2008. [Google Scholar]
- Hartsema, A.M. Influence of temperature on flower formation and flowering of bulbous and tuberous plants. Allium cepa, Onion. In Encyclopedia of Plant Physiology; Ruhland, W.W., Ed.; Springer: Berlin, Germany, 1961; Volume 16, pp. 123–167. [Google Scholar]
- Kamenetsky, R.; Dole, J. Herbaceous peony (Paeonia): Genetics, physiology and cut flower production. Floric. Ornam. Biotechnol. 2012, 6, 62–77. [Google Scholar]
- Roh, M.S.; Joung, Y.H. Inflorescence development in an Ornithogalum dubium hybrid as influenced by bulb temperature treatments. J. Hort. Sci. Biotech. 2004, 79, 576–581. [Google Scholar] [CrossRef]
- Noy-Porat, T.; Flaishman, M.A.; Eshel, A.; Sandler-Ziv, D.; Kamenetsky, R. Florogenesis of the Mediterranean geophyte Narcissus tazetta and temperature requirements for flower initiation and differentiation. Sci. Hortic. 2009, 120, 138–142. [Google Scholar] [CrossRef]
- De Hertogh, A.A.; Le Nard, M. Physiological and biochemical aspects of flower bulbs. In The Physiology of Flower Bulbs; De Hertogh, A.A., Le Nard, M., Eds.; Elsevier Science Publishers: Amsterdam, The Netherlands, 1993; pp. 53–70. [Google Scholar]
- Noy-Porat, T.; Cohen, D.; Mathew, D.; Eshel, A.; Kamenetsky, R.; Flaishman, M.A. Turned on by heat: Differential expression of LFY- FT and like genes in Narcissus tazetta during floral transition. J. Exp. Bot. 2013, 64, 3273–3284. [Google Scholar] [CrossRef] [Green Version]
- Li, X.F.; Wu, W.T.; Zhang, X.P.; Qiu, Y.; Zhang, W.; Li, R.; Xu, J.; Sun, Y.; Wang, Y.; Xu, L. Narcissus tazetta SVP-like gene NSVP1 affects flower development in Arabidopsis. J. Plant Physiol. 2015, 173, 89–96. [Google Scholar] [CrossRef]
- Leeggangers, H.A.C.F.; Nijveen, H.; Bigas, J.N.; Hilhorst, H.W.M.; Immink, R.G.H. Molecular Regulation of Temperature-Dependent Floral Induction in Tulipa gesneriana. Plant Physiol. 2017, 173, 1904–1919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Liu, Y.; Tang, X.; Rao, H.; Ren, C.; Chen, J.; Wu, Q.; Jiang, Y.; Geng, F.; Pei, J. Transcriptome profiling of the flowering transition in saffron (Crocus sativus L.). Sci. Rep. 2020, 10, 9680. [Google Scholar] [CrossRef]
- Lundmark, M.; Hurry, V.; Lapointe, L. Low temperature maximizes growth of Crocus vernus (L.) Hill via changes in carbon partitioning and corm development. J. Exp. Bot. 2009, 60, 2203–2213. [Google Scholar] [CrossRef]
- Rees, A.R. Ornamental Bulbs, Corms and Tubers; CAB International: Wallingford, UK, 1992; p. 220. [Google Scholar]
- De Hertogh, A.A. Holland Bulb Forcer´s Guide, 4th ed.; International Flower Bulb Center: Hillegom, The Netherlands, 1989. [Google Scholar]
- Elzinga, J.A.; Atlan, A.; Biere, A.; Gigord, L.; Weis, A.E.; Bernasconi, G. Time after time: Flowering phenology and biotic interactions. Trends Ecol. Evol. 2007, 22, 432–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinn, J.A.; Wetherington, J.D. Genetic variability and phenotypic plasticity in flowering phenology in populations of two grasses. J. Torrey Bot. Soc. 2002, 129, 96–106. [Google Scholar] [CrossRef]
- Franks, S.J.; Sim, S.; Weis, A.E. Rapid evolution of flowering time by an anual plant in response to a climate fluctuation. Proc. Natl. Acad. Sci. USA 2007, 104, 1278–1284. [Google Scholar] [CrossRef] [Green Version]
- Kerndorff1, H.; Pasche, E.; Harpke, D. The Genus Crocus (Liliiflorae, Iridaceae): Lifecycle, Morphology, Phenotypic Characteristics, and Taxonomical Relevant Parameters. STAPFIA 2015, 103, 27–65. [Google Scholar]
- Ivashchenko, A.; Belialov, O. Tulips and Other Bulbs Plants of Kazakhstan; Dve Stolizy: Almaty, Kazakhstan, 2005; p. 191. [Google Scholar]
- Petersen, G.; Seberg, O.; Thorsøe, S.; Jørgensen, T.; Mathew, B. A Phylogeny of the genus Crocus (Iridaceae) based on sequence data from five plastid regions. Taxon 2008, 57, 487–499. [Google Scholar]
- Tsaftaris, A.; Pasentsis, K.; Argiriou, A. Cloning and characterization of flowering locus T-like genes from the perennial geophyte saffron crocus (Crocus sativus). Plant Mol. Biol. Rep. 2013, 31, 1558–1568. [Google Scholar] [CrossRef]
- Tsaftaris, A.; Pasentsis, K.; Kalivas, A.; Michailidou, S.; Madesis, P.; Argiriou, A. Isolation of a centroradialis/terminal flower1 homolog in saffron (Crocus sativus L.): Characterization and expression analysis. Mol. Biol. Rep. 2012, 39, 7899–7910. [Google Scholar] [CrossRef] [PubMed]
- Haghighi, R.; Tabatabaei, B.E.S.; Maibody, S.A.M.M.; Talebi, M.; Molina, R.V.; Nebauer, S.G.; Renau-Morata, B. A flowering inhibitor of the temperature-dependent pathway in Crocus sativus L. Mol. Biol. Rep. 2020, 47, 2171–2179. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Storage Temperature before Planting | Temperature Regime after Planting | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Jun | Jul | Aug | Sep | Oct | Nov | Dec | Jan | Feb | Mar | Apr | May | ||
| 1 | Autumn Flowering C. serotinus | 25 °C | |||||||||||
| 2 | 23 °C | 17 °C | |||||||||||
| 3 | ↓ | 10 °C | |||||||||||
| 4 | Spring Flowering C. nevadensis C. vernus | 25 °C | |||||||||||
| 5 | ↓ | 10 °C | |||||||||||
| 6 | 23 °C | ↓ | 5–2–5 °C | 10 °C | |||||||||
| 7 | ↓ | 5–2–5 °C | ↑ | 17 °C | |||||||||
| Species | Population Number | Flowering Period | Altitude (m.a.s.l) | Temperature during Flowering (°C) | |||
|---|---|---|---|---|---|---|---|
| Earliest | Latest | DTF | Max. | Min. | |||
| C. serotinus | 14 | 17 Sep | 28 Oct | 64 a | 320–1454 | 18 | 6 |
| C. nudiflorus | 13 | 12 Sep | 15 Oct | 59 a | 910–2100 | 10 | 4 |
| C. clusii | 8 | 01 Nov | 02 Nov | 93 b | 14–88 | 19 | 18 |
| C.cambessedesii | 4 | 07 Nov | 09 Nov | 99 b | 30–250 | 20 | 18 |
| C. nevadensis | 12 | 16 Feb | 04 Mar | 210 c | 600–1800 | 11 | 4 |
| C. carpetanus | 5 | 24 Feb | 14 Mar | 211 c | 1100–1468 | 8 | 6 |
| C. vernus | 2 | 25 Apr | 03 May | 271 d | 1560 | 10 | 9 |
| Storage Temperature (°C) | Percentage of Flower Initiation | |||||||
|---|---|---|---|---|---|---|---|---|
| C. serotinus | C. nudiflorus | C. nevadensis | C. vernus | |||||
| 5 | - | 0 | c | - | - | |||
| 10 | 0 | c | 15.4 | b | 0 * | b | 0 * | c |
| 23 | 100 | a | 75 | a | 77.8 | a | 72.7 | a |
| 30 | 66.7 | b | 0 * | c | 0 | b | 7.6 | b |
| 35 | 0 | c | - | - | - | |||
| Species | Growth Temperatures | Flowering Period (days) | Flower Length (mm) | Flower Life (days) | |||
|---|---|---|---|---|---|---|---|
| C. serotinus | 10 °C | 17 | a | 223 | b | 8 | b |
| 17 °C | 9 | b | 204 | a | 6 | a | |
| C. nevadensis | 10 °C | 18 | b | 88 | a | 8 | b |
| Cold + 10 °C | 7 | a | 101 | b | 8 | b | |
| Cold + 17 °C | 4 | a | 85 | a | 5 | a | |
| C. vernus | 10 °C | 16 | b | 97 | a | 10 | b |
| Cold + 10 °C | 16 | b | 108 | b | 9 | b | |
| Cold + 17 °C | 7 | a | 96 | a | 7 | a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pastor-Férriz, T.; De-los-Mozos-Pascual, M.; Renau-Morata, B.; Nebauer, S.G.; Sanchis, E.; Busconi, M.; Fernández, J.-A.; Kamenetsky, R.; Molina, R.V. Ongoing Evolution in the Genus Crocus: Diversity of Flowering Strategies on the Way to Hysteranthy. Plants 2021, 10, 477. https://doi.org/10.3390/plants10030477
Pastor-Férriz T, De-los-Mozos-Pascual M, Renau-Morata B, Nebauer SG, Sanchis E, Busconi M, Fernández J-A, Kamenetsky R, Molina RV. Ongoing Evolution in the Genus Crocus: Diversity of Flowering Strategies on the Way to Hysteranthy. Plants. 2021; 10(3):477. https://doi.org/10.3390/plants10030477
Chicago/Turabian StylePastor-Férriz, Teresa, Marcelino De-los-Mozos-Pascual, Begoña Renau-Morata, Sergio G. Nebauer, Enrique Sanchis, Matteo Busconi, José-Antonio Fernández, Rina Kamenetsky, and Rosa V. Molina. 2021. "Ongoing Evolution in the Genus Crocus: Diversity of Flowering Strategies on the Way to Hysteranthy" Plants 10, no. 3: 477. https://doi.org/10.3390/plants10030477
APA StylePastor-Férriz, T., De-los-Mozos-Pascual, M., Renau-Morata, B., Nebauer, S. G., Sanchis, E., Busconi, M., Fernández, J. -A., Kamenetsky, R., & Molina, R. V. (2021). Ongoing Evolution in the Genus Crocus: Diversity of Flowering Strategies on the Way to Hysteranthy. Plants, 10(3), 477. https://doi.org/10.3390/plants10030477