Natural Variation in Plant Pluripotency and Regeneration



Abstract
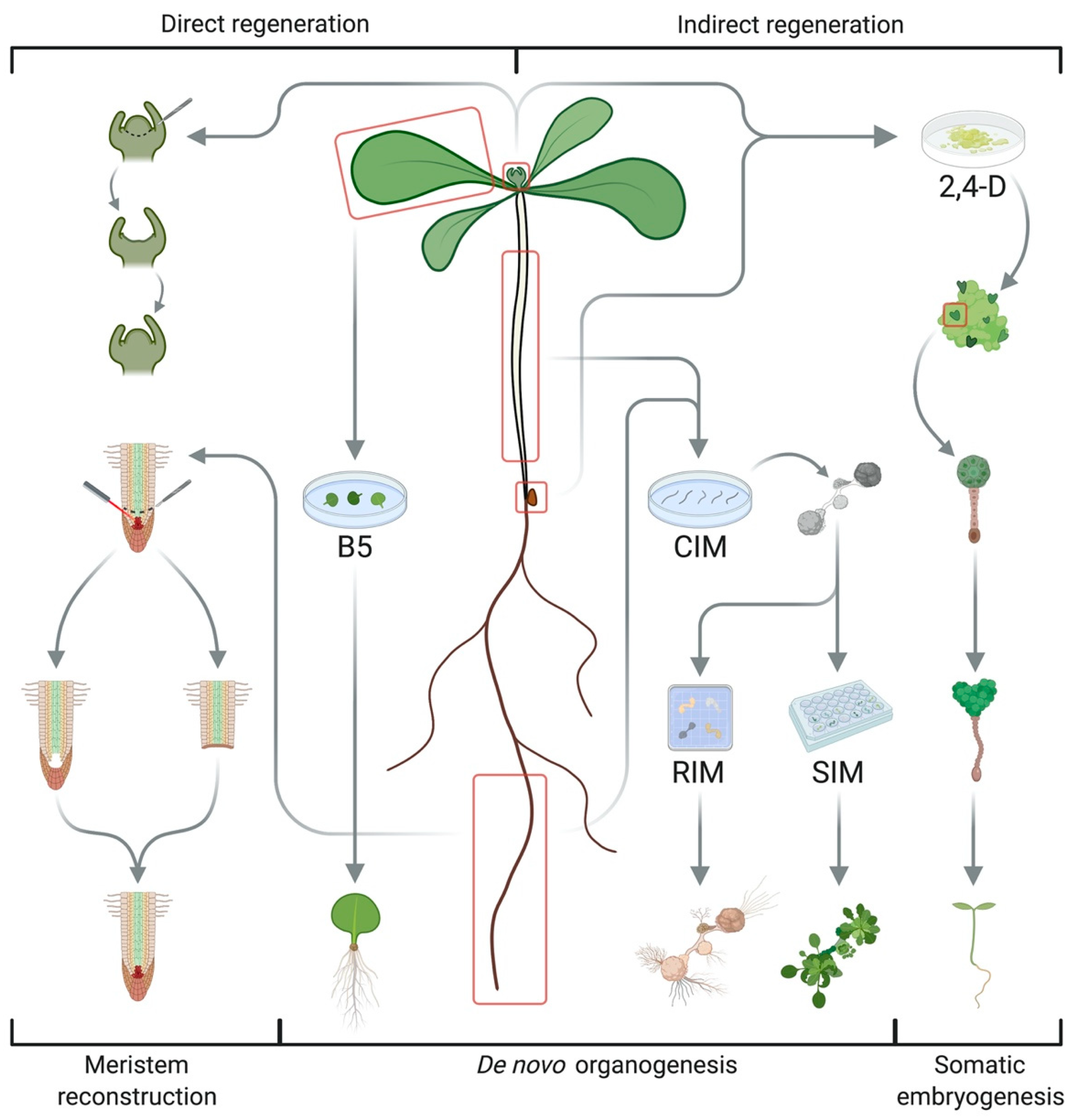
:1. Introduction: Definition, Origin, and Applications of Regeneration
2. Different Regeneration Systems
3. Cellular and Molecular Framework of de novo Shoot Organogenesis
3.1. Auxin and Cytokinin Signalling
3.2. Wound Responses
3.3. Founder Cell Specification
3.4. Callus Formation
3.5. Pluripotency Acquisition
3.6. Transdifferentiation
3.7. Shoot Promeristem Formation
3.8. SAM Patterning and Shoot Outgrowth
4. Mapping Natural Variation in the Organogenic Potential at the Genetic Level
4.1. Variation in Arabidopsis thaliana

{kind=link}
{kind=link}
| Species | Population | Method | Phenotype(s) | QTL(s) | QTG(s) | Reference |
|---|---|---|---|---|---|---|
| Arabidopsis thaliana | Col x Ler RILs | MAPMAKER QTL analysis | Callus and shoot formation from root or leaf explants | 9 (chr 1, 4, and 5) | (SRD1) | Schiantarelli et al. 2001 [83] |
| Col x Ler RILs | Composite interval mapping | Shoot regeneration from root explants | 3 (chr 1, 4, and 5) | ARR18, AGL6, HB17, AT4G36590, AT5G50820, AT1G01900, AT5G59120, AT4G26330 | Lall et al. 2004 [85] | |
| Ler x Cvi RILs | QTL interval mapping | Shoot or root regeneration from root or leaf explants | 17 (chr 1, 2, and 5) | (SRD3, RRD4, IRE1, CKH1, AXR1) | Velázquez et al. 2004 [82] | |
| Nok-3 x Ga-0 RILs | Linkage and association mapping | Shoot regeneration from root explants | 5 (chr 1–3) | RPK1 | Motte et al. 2014 [81] | |
| 48 ecotypes | Linkage disequilibrium analysis | Shoot regeneration from root explants | 1 (chr 5) | DCC1 | Zhang et al. 2018 [91] | |
| 170 natural SALK strains | GWAS | Shoot regeneration from root explants | 86 (chr 1–5) | WUS, SUP, AT3G09925, DOF4.4, EDA40, QUL2, URH1, RLP9, QKY, ARF20, MSL3, DREB1A, WAVH2, MIR393A, … | Lardon et al. 2020 [80] |
4.2. Variation in Other Species
4.2.1. Monocot Crops
| Species | Population | Method | Phenotype(s) | QTL(s) | QTG(s) | Reference |
|---|---|---|---|---|---|---|
| Oryza sativa | Norin 1 x Tadukan F2 | QTL interval mapping | Shoot regeneration from mature seed-derived callus | 2 (chr 2 and 4) | - | Takeuchi et al. 2000 [96] |
| Milyang 23 x Gihobyeo RILs | RFLP analysis | Shoot regeneration from mature seed-derived callus | 6 (chr 1–3 and 11) | - | Kwon et al. 2001 [97] | |
| Koshihikari x Kasalath BC1-3F2 | Map-based cloning | Shoot regeneration from mature seed-derived callus | 4 (chr 1–3 and 6) | PSR1/NIR | Nishimura et al. 2005 [98] | |
| Koshihikari x Kasalath BC1F3 and NILs | MAPMAKER QTL analysis | Callus induction and regeneration from mature seeds | 8 (chr 1, 4, and 9) | - | Taguchi-Shiobara et al. 2006 [93] | |
| Nipponbare x Zhenshan 97B CSSLs | Stepwise regression of CSSLs | Callus induction and regeneration from mature seeds | 29 (chr 1, 3, and 10) | - | Zhao et al. 2009 [99] | |
| Nipponbare x 93-11 RILs | Composite interval mapping | Callus induction and regeneration from mature seeds | 25 (chr 3 and 7) | - | Li et al. 2013 [94] | |
| Pei’ai 64s x Yangdao 6 RILs | QTL mapping | Callus induction from mature seeds | 8 (chr 5–7, 9, and 10) | - | Tian et al., 2013 [100] | |
| 510 natural accessions | GWAS | Callus induction from mature seeds | 88 (chr 1–12) | OsBBM1, OsSET1, OsIAA10, CRL1, … | Zhang et al. 2019 [102] | |
| Teqing x YIL25 F2 | Map-based cloning | Callus browning | 1 (chr 3) | BOC1 | Zhang et al. 2020 [103] | |
| Zea Mays | A188 x B73 NILs | Segregation distortion analysis | Callus induction and regeneration from mature embryos | 1 (chr 3) | ALD1, ZmWOX2A, ZmWOX5B | Salvo et al. 2018 [106] |
| 144 inbred lines | GWAS | Callus induction and regeneration from immature embryos | 63 (chr 1–10) | ZmWOX2, ZmOCL5A, ZmBR2, ZmKIP1, ZmDEK35, ZmSBP18, … | Ma et al. 2018 [105] | |
| Triticum aestivum | Wangshuibai x Nanda 2419 RILs | Simple and composite interval mapping | Callus induction and regeneration from mature embryos | 13 (chr 2A, 2D, 5A, 5B, and 5D) | - | Jia et al. 2007 [108] |
| Svilena x Jensen F3 | DArT-based QTL mapping | Green plantlet regeneration from microspore cultures | 2 (chr 1B and 7B) | - | Nielsen et al. 2015 [114] | |
| Chuanmai 32 x SHW-L1 RILs | DArT-based QTL mapping | Callus induction and regeneration from mature embryos | 6 (chr 1A, 1D, 3B, 4A, 5A, 6D) | - | Ma et al. 2016 [109] | |
| Hordeum Vulgare | Azumamugi x Kanto Nakate Gold RILs | Composite interval mapping | Callus induction and shoot regeneration from immature embryo cultures | 8 (chr 1–3H, 5H, and 7H) | UZU, SHD1, VRS1 | Mano et al. 2002 [111] |
| Steptoe x Morex DHs | EST-based linkage mapping | Green and albino plant regeneration | 8 (chr 1–7H) | HvSTM, HvPKL, HvLEC, HvBBM, HvESR1, HvCUC, HvAGL24, … | Tyagi et al. 2010 [112] | |
| Haruna Nijo x Golden Promise F2 | Segregation distortion analysis | Transformation amenability from immature embryos | 10 (chr 1–6H) | (HvNIR) | Hisano et al. 2016 [115] | |
| Full Pint x Golden Promise DHs | Segregation distortion analysis | Transformation amenability from immature embryos | 3 (chr 2–3H) | HvBBM, HvWUS2 | Hisano et al. 2017 [116] |
4.2.2. Dicot Crops
4.2.3. Ornamentals and Commodity Crops
| Species | Population | Method | Phenotype(s) | QTL(s) | QTG(s) | Reference |
|---|---|---|---|---|---|---|
| Lycopersicon esculentum | L. chilense x KOT BC1F2 | Bulked segregant analysis | Shoot regeneration from root explants | 1 (chr 3) | RG-2/INVCHI | Satoh et al. 2000 [118] |
| S. pennellii x Anl27 F2 and BC1 | QTL interval mapping | Shoot regeneration from leaf disks | 6 (chr 1, 3, 4, 7, and 8) | RG-3, (LESK1, ESR1&2) | Trujillo-Moya et al. 2011 [119] | |
| Cucumis sativus | 9110 Gt x 9930 RILs and 115 core accessions | QTL interval mapping and GWAS | Shoot regeneration from cotyledon explants | 4 (chr 1, 3, and 6) | ATJ3 | Wang et al. 2018 [121] |
| Raphanus sativus | GX71 x GX50 F1 | Composite interval mapping | Somatic embryogenesis from microspore cultures | 5 (chr 3, 8, and 9) | PRC2, FLAVIN-BINDING MONO-OXYGENASE, SET DOMAIN PROTEIN | Kim et al. 2020 [122] |
| Brassica oleracea | CGC 3-1 x Daehnfelt 360-7 F2 | Simple interval mapping | Shoot regeneration from protoplast-derived calli | 2 (-) | - | Holme et al. 2004 [123] |
| Rosa sp. | 96 cultivars | GWAS | Direct shoot regeneration from leaf petioles | 88 (chr 1 and 3–6) | GT2-like, RAP2.7-like, MET3-like, KNOX1-like 3, ANP1-like, GAI-like, YAB2, BIG, WUS, CUC1, SERK1, RPK1, … | Nguyen et al. 2017 [125] |
| Helianthus annuus | PAC-2 x RHA-266 RILS | Composite interval mapping | Shoot regeneration from cotyledon explants | 13 (chr 2, 6–9, 15, and 17) | - | Flores Berrios et al. 2000a [127] |
| PAC-2 x RHA-266 RILS | Composite interval mapping | Somatic embryogenesis from epidermal layers | 11 (chr 1, 3, 4, 6, 11, 13, and 15–17) | - | Flores Berrios et al. 2000b [126] | |
| Populus trichocarpa | 280 genotypes | GWAS and co-expression analysis | Callus formation from parenchyma cells | 8 (chr 3, 4, 6, 8, 9, 12, 15, and 18) | SOK1, MAPK3, SCR-like, RALF-like, WUS, LBD16, LEC1&2, CLF, TSD1, … | Tuskan et al. 2018 [128] |
5. Other Sources of Regenerative Variation
5.1. Epigenetic and Transcriptional Variation
5.2. Source of the Explant and Hormone Responsivity
5.3. Environmental Influences
6. Implications and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pulianmackal, A.J.; Kareem, A.V.K.; Durgaprasad, K.; Trivedi, Z.B.; Prasad, K. Competence and regulatory interactions during regeneration in plants. Front. Plant Sci. 2014, 5, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Huang, H. Genetic and Epigenetic Controls of Plant Regeneration, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2014; Volume 108, ISBN 9780123914989. [Google Scholar]
- Perianez-Rodriguez, J.; Manzano, C.; Moreno-Risueno, M.A. Post-embryonic organogenesis and plant regeneration from tissues: Two sides of the same coin? Front. Plant Sci. 2014, 5, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motte, H.; Vereecke, D.; Geelen, D.; Werbrouck, S. The molecular path to in vitro shoot regeneration. Biotechnol. Adv. 2014, 32, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.; Bera, T. Somatic Embryogenesis and Organogenesis; Elsevier Inc.: Amsterdam, The Netherlands, 2015; ISBN 9780128024980. [Google Scholar]
- Bidabadi, S.S.; Mohan Jain, S. Cellular, molecular, and physiological aspects of in vitro plant regeneration. Plants 2020, 9, 702. [Google Scholar] [CrossRef] [PubMed]
- Ikeuchi, M.; Favero, D.S.; Sakamoto, Y.; Iwase, A.; Coleman, D.; Rymen, B.; Sugimoto, K. Molecular Mechanisms of Plant Regeneration. Annu. Rev. Plant Biol. 2019, 70, 377–406. [Google Scholar] [CrossRef] [PubMed]
- Horstman, A.; Bemer, M.; Boutilier, K. A transcriptional view on somatic embryogenesis. Regeneration 2017, 4, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Nishihama, R.; Ishizaki, K.; Hosaka, M.; Matsuda, Y.; Kubota, A.; Kohchi, T. Phytochrome-mediated Regulation of Cell Division and Growth During Regeneration and Sporeling Development in the Liverwort Marchantia Polymorpha. J. Plant Res. 2015, 128. [Google Scholar] [CrossRef] [Green Version]
- Ikeuchi, M.; Ogawa, Y.; Iwase, A.; Sugimoto, K. Plant regeneration: Cellular origins and molecular mechanisms. Development 2016, 143, 1442–1451. [Google Scholar] [CrossRef] [Green Version]
- Kareem, A.; Radhakrishnan, D.; Sondhi, Y.; Aiyaz, M.; Roy, M.V.; Sugimoto, K.; Prasad, K. De novo assembly of plant body plan: A step ahead of Deadpool. Regeneration 2016, 3, 182–197. [Google Scholar] [CrossRef]
- Méndez-Hernández, H.A.; Ledezma-Rodríguez, M.; Avilez-Montalvo, R.N.; Juárez-Gómez, Y.L.; Skeete, A.; Avilez-Montalvo, J.; De-La-Peña, C.; Loyola-Vargas, V.M. Signaling overview of plant somatic embryogenesis. Front. Plant Sci. 2019, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Ikeda-Iwai, M.; Umehara, M.; Satoh, S.; Kamada, H. Stress-induced somatic embryogenesis in vegetative tissues of Arabidopsis thaliana. Plant J. 2003, 34, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Nic-Can, G.I.; Avilez-Montalvo, J.R.; Aviles-Montalvo, R.N.; Márquez-López, R.E.; Mellado-Mojica, E.; Galaz-Ávalos, R.M.; Loyola-Vargas, V.M. The relationship between stress and somatic embryogenesis. In Somatic Embryogenesis: Fundamental Aspects and Applications; Springer: Berlin/Heidelberg, Germany, 2016; pp. 121–170. ISBN 9783319337050. [Google Scholar]
- Xu, L. De novo root regeneration from leaf explants: Wounding, auxin, and cell fate transition. Curr. Opin. Plant Biol. 2018, 41, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Qu, Y.; Sheng, L.; Liu, J.; Huang, H.; Xu, L. A simple method suitable to study de novo root organogenesis. Front. Plant Sci. 2014, 5, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Sheng, L.; Xu, Y.; Li, J.; Yang, Z.; Huang, H.; Xu, L. WOX11 and 12 are involved in the first-step cell fate transition during de novo root organogenesis in Arabidopsis. Plant Cell 2014, 26, 1081–1093. [Google Scholar] [CrossRef] [Green Version]
- Valvekens, D.; Van Montagu, M.; Van Lijsebettens, M. Agrobacterium tumefaciens-mediated transformation of Arabidopsis thaliana root explants by using kanamycin selection. Proc. Natl. Acad. Sci. USA 1988, 85, 5536–5540. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.; Bae, S.; Seo, P.J. De novo shoot organogenesis during plant regeneration. J. Exp. Bot. 2020, 71, 63–72. [Google Scholar] [CrossRef]
- Radhakrishnan, D.; Kareem, A.; Durgaprasad, K.; Sreeraj, E.; Sugimoto, K.; Prasad, K. Shoot regeneration: A journey from acquisition of competence to completion. Curr. Opin. Plant Biol. 2018, 41, 23–31. [Google Scholar] [CrossRef]
- Tian, X.; Zhang, C.; Xu, J. Control of Cell Fate Reprogramming Towards de Novo Shoot Organogenesis. Plant Cell Physiol. 2018, 59, 708–714. [Google Scholar] [CrossRef] [Green Version]
- Sang, Y.L.; Cheng, Z.J.; Zhang, X.S. Plant stem cells and de novo organogenesis. New Phytol. 2018, 218, 1334–1339. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.H.; Zhang, X.S. The Hormonal Control of Regeneration in Plants. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 108, pp. 35–69. [Google Scholar]
- Schaller, G.E.; Bishopp, A.; Kieber, J.J. The yin-yang of hormones: Cytokinin and auxin interactions in plant development. Plant Cell 2015, 27, 44–63. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.J.; Wang, L.; Sun, W.; Zhang, Y.; Zhou, C.; Su, Y.H.; Li, W.; Sun, T.T.; Zhao, X.Y.; Li, X.G.; et al. Pattern of auxin and cytokinin responses for shoot meristem induction results from the regulation of cytokinin biosynthesis by AUXIN RESPONSE FACTOR3. Plant Physiol. 2013, 161, 240–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.-Q.; Lian, H.; Zhou, C.-M.; Xu, L.; Jiao, Y.; Wang, J.-W. A Two-Step Model for de Novo Activation of WUSCHEL during Plant Shoot Regeneration. Plant Cell Online 2017, 29, 1073–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwase, A.; Mita, K.; Nonaka, S.; Ikeuchi, M.; Koizuka, C.; Ohnuma, M.; Ezura, H.; Imamura, J.; Sugimoto, K. WIND1-based acquisition of regeneration competency in Arabidopsis and rapeseed. J. Plant Res. 2015, 128, 389–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeuchi, M.; Iwase, A.; Rymen, B.; Lambolez, A.; Kojima, M.; Takebayashi, Y.; Heyman, J.; Watanabe, S.; Seo, M.; De Veylder, L.; et al. Wounding triggers callus formation via dynamic hormonal and transcriptional changes. Plant Physiol. 2017, 175, 1158–1174. [Google Scholar] [CrossRef] [Green Version]
- Iwase, A.; Mitsuda, N.; Koyama, T.; Hiratsu, K.; Kojima, M.; Arai, T.; Inoue, Y.; Seki, M.; Sakakibara, H.; Sugimoto, K.; et al. The AP2/ERF transcription factor WIND1 controls cell dedifferentiation in arabidopsis. Curr. Biol. 2011, 21, 508–514. [Google Scholar] [CrossRef] [Green Version]
- Iwase, A.; Ohme-Takagi, M.; Sugimoto, K. WIND1: A key molecular switch for plant cell dedifferentiation. Plant Signal. Behav. 2011, 6, 1943–1945. [Google Scholar] [CrossRef] [Green Version]
- Iwase, A.; Harashima, H.; Ikeuchi, M.; Rymen, B.; Ohnuma, M.; Komaki, S.; Morohashi, K.; Kurata, T.; Nakata, M.; Ohme-Takagi, M.; et al. WIND1 Promotes Shoot Regeneration through Transcriptional Activation of ENHANCER OF SHOOT REGENERATION1 in Arabidopsis. Plant Cell 2017, 29, 54–69. [Google Scholar] [CrossRef] [Green Version]
- Rymen, B.; Kawamura, A.; Lambolez, A.; Inagaki, S.; Takebayashi, A.; Iwase, A.; Sakamoto, Y.; Sako, K.; Favero, D.S.; Ikeuchi, M.; et al. Histone acetylation orchestrates wound-induced transcriptional activation and cellular reprogramming in Arabidopsis. Commun. Biol. 2019, 2, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.; Seo, P.J. Varying auxin levels induce distinct pluripotent states in callus cells. Front. Plant Sci. 2018, 871, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Ikeuchi, M.; Sugimoto, K.; Iwase, A. Plant callus: Mechanisms of induction and repression. Plant Cell 2013, 25, 3159–3173. [Google Scholar] [CrossRef] [Green Version]
- Che, P.; Lall, S.; Howell, S.H. Developmental steps in acquiring competence for shoot development in Arabidopsis tissue culture. Planta 2007, 226, 1183–1194. [Google Scholar] [CrossRef] [PubMed]
- Atta, R.; Laurens, L.; Boucheron-Dubuisson, E.; Guivarc’h, A.; Carnero, E.; Giraudat-Pautot, V.; Rech, P.; Chriqui, D. Pluripotency of Arabidopsis xylem pericycle underlies shoot regeneration from root and hypocotyl explants grown in vitro. Plant J. 2009, 57, 626–644. [Google Scholar] [CrossRef] [PubMed]
- Duclercq, J.; Sangwan-Norreel, B.; Catterou, M.; Sangwan, R.S. De novo shoot organogenesis: From art to science. Trends Plant Sci. 2011, 16, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Shang, B.; Xu, C.; Zhang, X.; Cao, H.; Xin, W.; Hu, Y. Very-long-chain fatty acids restrict regeneration capacity by confining pericycle competence for callus formation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2016, 113, 5101–5106. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, K.; Jiao, Y.; Meyerowitz, E.M. Arabidopsis regeneration from multiple tissues occurs via a root development pathway. Dev. Cell 2010, 18, 463–471. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Liu, J.; Fan, M.; Xin, W.; Hu, Y.; Xu, C. A genome-wide transcriptome profiling reveals the early molecular events during callus initiation in Arabidopsis multiple organs. Genomics 2012, 100, 116–124. [Google Scholar] [CrossRef] [Green Version]
- Fan, M.; Xu, C.; Xu, K.; Hu, Y. LATERAL ORGAN BOUNDARIES DOMAIN transcription factors direct callus formation in Arabidopsis regeneration. Cell Res. 2012, 22, 1169–1180. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Park, O.S.; Seo, P.J. Arabidopsis ATXR2 deposits H3K36me3 at the promoters of LBD genes to facilitate cellular dedifferentiation. Sci. Signal. 2017, 10. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Park, O.-S.; Seo, P.J. JMJ30-mediated demethylation of H3K9me3 drives tissue identity changes to promote callus formation in Arabidopsis. Plant J. 2018, 95, 961–975. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Seo, P.J. High-temperature promotion of callus formation requires the BIN2-ARF-LBD axis in Arabidopsis. Planta 2017, 246, 797–802. [Google Scholar] [CrossRef]
- Liu, H.; Ma, X.; Han, H.N.; Hao, Y.J.; Zhang, X.S. AtPRMT5 Regulates Shoot Regeneration through Mediating Histone H4R3 Dimethylation on KRPs and Pre-mRNA Splicing of RKP in Arabidopsis. Mol. Plant 2016, 9, 1634–1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Cao, H.; Xu, E.; Zhang, S.; Hu, Y. Genome-Wide Identification of Arabidopsis LBD29 Target Genes Reveals the Molecular Events behind Auxin-Induced Cell Reprogramming during Callus Formation. Plant Cell Physiol. 2018, 59, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Cao, H.; Zhang, Q.; Wang, H.; Xin, W.; Xu, E.; Zhang, S.; Yu, R.; Yu, D.; Hu, Y. Control of auxin-induced callus formation by bZIP59-LBD complex in Arabidopsis regeneration. Nat. Plants 2018, 4, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Buechel, S.; Leibfried, A.; To, J.P.C.; Zhao, Z.; Andersen, S.U.; Kieber, J.J.; Lohmann, J.U. Role of A-type Arabidopsis Response Regulators in meristem maintenance and regeneration. Eur. J. Cell Biol. 2010, 89, 279–284. [Google Scholar] [CrossRef]
- Liu, Z.; Li, J.; Wang, L.; Li, Q.; Lu, Q.; Yu, Y.; Li, S.; Bai, M.-y.; Hu, Y.; Xiang, F. Repression of callus initiation by the miRNA-directed interaction of auxin–cytokinin in Arabidopsis thaliana. Plant J. 2016, 87, 391–402. [Google Scholar] [CrossRef]
- Kim, J.; Yang, W.; Forner, J.; Lohmann, J.U.; Noh, B.; Noh, Y. Epigenetic reprogramming by histone acetyltransferase HAG1/AtGCN5 is required for pluripotency acquisition in Arabidopsis. EMBO J. 2018, 37, 1–16. [Google Scholar] [CrossRef]
- Kareem, A.; Durgaprasad, K.; Sugimoto, K.; Du, Y.; Pulianmackal, A.J.; Trivedi, Z.B.; Abhayadev, P.V.; Pinon, V.; Meyerowitz, E.M.; Scheres, B.; et al. PLETHORA genes control regeneration by a two-step mechanism. Curr. Biol. 2015, 25, 1017–1030. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Hu, X.; Qin, P.; Prasad, K.; Hu, Y.; Xu, L. The WOX11-LBD16 Pathway Promotes Pluripotency Acquisition in Callus Cells during de Novo Shoot Regeneration in Tissue Culture. Plant Cell Physiol. 2018, 59, 734–743. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Chen, X.; Huang, H.; Xu, L. Reprogramming of H3K27me3 Is Critical for Acquisition of Pluripotency from Cultured Arabidopsis Tissues. PLoS Genet. 2012, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Park, O.S.; Jung, S.J.; Seo, P.J. Histone deacetylation-mediated cellular dedifferentiation in Arabidopsis. J. Plant Physiol. 2016, 191, 95–100. [Google Scholar] [CrossRef]
- Lafos, M.; Kroll, P.; Hohenstatt, M.L.; Thorpe, F.L.; Clarenz, O.; Schubert, D. Dynamic regulation of H3K27 trimethylation during Arabidopsis differentiation. PLoS Genet. 2011, 7, e1002040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishihara, H.; Sugimoto, K.; Tarr, P.T.; Temman, H.; Kadokura, S.; Inui, Y.; Sakamoto, T.; Sasaki, T.; Aida, M.; Suzuki, T.; et al. Primed histone demethylation regulates shoot regenerative competency. Nat. Commun. 2019, 10, 1786. [Google Scholar] [CrossRef] [Green Version]
- Shemer, O.; Landau, U.; Candela, H.; Zemach, A.; Eshed Williams, L. Competency for shoot regeneration from Arabidopsis root explants is regulated by DNA methylation. Plant Sci. 2015, 238, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, K.; Gordon, S.P.; Meyerowitz, E.M. Regeneration in plants and animals: Dedifferentiation, transdifferentiation, or just differentiation? Trends Cell Biol. 2011, 21, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Rosspopoff, O.; Chelysheva, L.; Saffar, J.; Lecorgne, L.; Gey, D.; Caillieux, E.; Colot, V.; Roudier, F.; Hilson, P.; Berthomé, R.; et al. Direct conversion of root primordium into shoot meristem relies on timing of stem cell niche development. Development 2017, 144, 1187–1200. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S.P.; Heisler, M.G.; Reddy, G.V.; Ohno, C.; Das, P.; Meyerowitz, E.M. Pattern formation during de novo assembly of the Arabidopsis shoot meristem. Development 2007, 134, 3539–3548. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, Y.; Banno, H.; Niu, Q.W.; Howell, S.H.; Chua, N.H. The ENHANCER of SHOOT REGENERATION 2 gene in Arabidopsis regulates CUP-SHAPED COTYLEDON 1 at the transcriptional level and controls cotyledon development. Plant Cell Physiol. 2006, 47, 1443–1456. [Google Scholar] [CrossRef] [Green Version]
- Takeda, S.; Hanano, K.; Kariya, A.; Shimizu, S.; Zhao, L.; Matsui, M.; Tasaka, M.; Aida, M. CUP-SHAPED COTYLEDON1 transcription factor activates the expression of LSH4 and LSH3, two members of the ALOG gene family, in shoot organ boundary cells. Plant J. 2011, 66, 1066–1077. [Google Scholar] [CrossRef]
- Dai, X.; Liu, Z.; Qiao, M.; Li, J.; Li, S.; Xiang, F. ARR12 promotes de novo shoot regeneration in Arabidopsis thaliana via activation of WUSCHEL expression. J. Integr. Plant Biol. 2017, 59, 747–758. [Google Scholar] [CrossRef] [Green Version]
- Meng, W.J.; Cheng, Z.J.; Sang, Y.L.; Zhang, M.M.; Rong, X.F.; Wang, Z.W.; Tang, Y.Y.; Zhang, X.S. Type-B ARABIDOPSIS RESPONSE REGULATORs Specify the Shoot Stem Cell Niche by Dual Regulation of WUSCHEL. Plant Cell 2017, 29, 1357–1372. [Google Scholar] [CrossRef] [Green Version]
- Zubo, Y.O.; Blakley, I.C.; Yamburenko, M.V.; Worthen, J.M.; Street, I.H.; Franco-Zorrilla, J.M.; Zhang, W.; Hill, K.; Raines, T.; Solano, R.; et al. Cytokinin induces genome-wide binding of the type-B response regulator ARR10 to regulate growth and development in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, E5995–E6004. [Google Scholar] [CrossRef] [Green Version]
- Xue, T.; Dai, X.; Wang, R.; Wang, J.; Liu, Z.; Xiang, F. ARGONAUTE10 inhibits in vitro shoot regeneration via repression of miR165/166 in arabidopsis thaliana. Plant Cell Physiol. 2017, 58, 1789–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Poretska, O.; Sieberer, T. ALTERED MERISTEM PROGRAM1 restricts shoot meristem proliferation and regeneration by limiting HD-ZIP III-mediated expression of RAP2.6l. Plant Physiol. 2018, 177, 1580–1594. [Google Scholar] [CrossRef] [PubMed]
- Shi, B.; Zhang, C.; Tian, C.; Wang, J.; Wang, Q.; Xu, T.; Xu, Y.; Ohno, C.; Sablowski, R.; Heisler, M.G.; et al. Two-Step Regulation of a Meristematic Cell Population Acting in Shoot Branching in Arabidopsis. PLoS Genet. 2016, 12, e1006168. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Miotk, A.; Šutiković, Z.; Ermakova, O.; Wenzl, C.; Medzihradszky, A.; Gaillochet, C.; Forner, J.; Utan, G.; Brackmann, K.; et al. WUSCHEL acts as an auxin response rheostat to maintain apical stem cells in Arabidopsis. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Leibfried, A.; To, J.P.C.; Busch, W.; Stehling, S.; Kehle, A.; Demar, M.; Kieber, J.J.; Lohmann, J.U. WUSCHEL controls meristem function by direct regulation of cytokinin-inducible response regulators. Nature 2005, 438, 1172–1175. [Google Scholar] [CrossRef]
- Negin, B.; Shemer, O.; Sorek, Y.; Eshed Williams, L. Shoot stem cell specification in roots by the WUSCHEL transcription factor. PLoS ONE 2017, 12, e0176093. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Liu, H.; Cheng, Z.J.; Su, Y.H.; Han, H.N.; Zhang, Y.; Zhang, X.S. DNA Methylation and Histone Modifications Regulate De Novo Shoot Regeneration in Arabidopsis by Modulating WUSCHEL Expression and Auxin Signaling. PLoS Genet. 2011, 7, e1002243. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zhang, H.; Dong, Y.X.; Hao, Y.J.; Zhang, X.S. DNA METHYLTRANSFERASE1-mediated shoot regeneration is regulated by cytokinin-induced cell cycle in Arabidopsis. New Phytol. 2017, 217, 219–232. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.-Q.; Lian, H.; Tang, H.; Dolezal, K.; Zhou, C.-M.; Yu, S.; Chen, J.-H.; Chen, Q.; Liu, H.; Ljung, K.; et al. An intrinsic microRNA timer regulates progressive decline in shoot regenerative capacity in plants. Plant Cell 2015, 27, 349–360. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S.P.; Chickarmane, V.S.; Ohno, C.; Meyerowitz, E.M. Multiple feedback loops through cytokinin signaling control stem cell number within the Arabidopsis shoot meristem. Proc. Natl. Acad. Sci. USA 2009, 106, 16529–16534. [Google Scholar] [CrossRef] [Green Version]
- Perales, M.; Reddy, G.V. Stem cell maintenance in shoot apical meristems. Curr. Opin. Plant Biol. 2012, 15, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, N.; Banno, H. Arabidopsis ENHANCER OF SHOOT REGENERATION 2 and PINOID are involved in in vitro shoot regeneration. Plant Biotechnol. 2012, 29, 367–372. [Google Scholar] [CrossRef] [Green Version]
- Reinhardt, D.; Pesce, E.R.; Stieger, P.; Mandel, T.; Baltensperger, K.; Bennett, M.; Traas, J.; Friml, J.; Kuhlemeier, C. Regulation of phyllotaxis by polar auxin transport. Nature 2003, 426, 255–260. [Google Scholar] [CrossRef]
- Lee, K.; Park, O.S.; Choi, C.Y.; Seo, P.J. ARABIDOPSIS TRITHORAX 4 Facilitates Shoot Identity Establishment during the Plant Regeneration Process. Plant Cell Physiol. 2019, 60, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Lardon, R.; Wijnker, E.; Keurentjes, J.; Geelen, D. The genetic framework of shoot regeneration in Arabidopsis comprises master regulators and conditional fine-tuning factors. Commsbio 2020. accepted. [Google Scholar]
- Motte, H.; Vercauteren, A.; Depuydt, S.; Landschoot, S.; Geelen, D.; Werbrouck, S.; Goormachtig, S.; Vuylsteke, M.; Vereecke, D. Combining linkage and association mapping identifies RECEPTOR-LIKE PROTEIN KINASE1 as an essential Arabidopsis shoot regeneration gene. Proc. Natl. Acad. Sci. USA 2014, 111, 8305–8310. [Google Scholar] [CrossRef] [Green Version]
- Velázquez, I.; Valencia, S.; López-Lera, A.; de La Peña, A.; Candela, M. Analysis of natural allelic variation in in vitro organogenesis of Arabidopsis thaliana. Euphytica 2004, 137, 73–79. [Google Scholar] [CrossRef]
- Schiantarelli, E.; De la Peña, A.; Candela, M. Use of recombinant inbred lines (RILs) to identify, locate and map major genes and quantitative trait loci involved with in vitro regeneration ability in Arabidopsis thaliana. Theor. Appl. Genet. 2001, 102, 335–341. [Google Scholar] [CrossRef]
- Ozawa, S.; Yasutani, I.; Fukuda, H.; Komamine, A.; Sugiyama, M. Organogenic responses in tissue culture of srd mutants of Arabidopsis thaliana. Development 1998, 125, 135–142. [Google Scholar]
- Lall, S.; Nettleton, D.; DeCook, R.; Che, P.; Howell, S.H. Quantitative trait loci associated with adventitious shoot formation in tissue culture and the program of shoot development in Arabidopsis. Genetics 2004, 167, 1883–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nodine, M.D.; Yadegari, R.; Tax, F.E. RPK1 and TOAD2 Are Two Receptor-like Kinases Redundantly Required for Arabidopsis Embryonic Pattern Formation. Dev. Cell 2007, 12, 943–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nodine, M.D.; Tax, F.E. Two receptor-like kinases required together for the establishment of Arabidopsis cotyledon primordia. Dev. Biol. 2008, 314, 161–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luichtl, M.; Fiesselmann, B.S.; Matthes, M.; Yang, X.; Peis, O.; Brunner, A.; Torres-Ruiz, R.A. Mutations in the Arabidopsis RPK1 gene uncouple cotyledon anlagen and primordia by modulating epidermal cell shape and polarity. Biol. Open 2013, 2, 1093–1102. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, A.; Betsuyaku, S.; Osakabe, Y.; Mizuno, S.; Nagawa, S.; Stahl, Y.; Simon, R.; Yamaguchi-Shinozaki, K.; Fukuda, H.; Sawa, S. RPK2 is an essential receptor-like kinase that transmits the CLV3 signal in Arabidopsis. Development 2010, 137, 3911–3920. [Google Scholar] [CrossRef] [Green Version]
- Fiesselmann, B.S.; Luichtl, M.; Yang, X.; Matthes, M.; Peis, O.; Torres-Ruiz, R.A. Ectopic shoot meristem generation in monocotyledonous rpk1 mutants is linked to SAM loss and altered seedling morphology. BMC Plant Biol. 2015, 15, 171. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, T.T.; Liu, H.; Shi, D.Y.; Wang, M.; Bie, X.M.; Li, X.G.; Zhang, X.S. Thioredoxin-mediated ROS homeostasis explains natural variation in plant regeneration. Plant Physiol. 2018, 176, 2231–2250. [Google Scholar] [CrossRef] [Green Version]
- Marchadier, E.; Hanemian, M.; Tisné, S.; Bach, L.; Bazakos, C.; Gilbault, E.; Haddadi, P.; Virlouvet, L.; Loudet, O. The complex genetic architecture of shoot growth natural variation in Arabidopsis thaliana. PLoS Genet. 2019, 15, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Taguchi-Shiobara, F.; Yamamoto, T.; Yano, M.; Oka, S. Mapping QTLs that control the performance of rice tissue culture and evaluation of derived near-isogenic lines. Theor. Appl. Genet. 2006, 112, 968–976. [Google Scholar] [CrossRef]
- Li, S.; Yan, S.; Wang, A.; Zou, G.; Huang, X.; Han, B.; Qian, Q.; Tao, Y. Identification of QTLs associated with tissue culture response through sequencing-based genotyping of RILs derived from 93-11 × Nipponbare in rice (Oryza sativa). Plant Cell Rep. 2013, 32, 103–116. [Google Scholar] [CrossRef]
- Taguchi-Shiobara, F.; Lin, S.Y.; Tanno, K.; Komatsuda, T.; Yano, M.; Sasaki, T.; Oka, S. Mapping quantitative trait loci associated with regeneration ability of seed callus in rice, Oryza sativa L. Theor. Appl. Genet. 1997, 95, 828–833. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Abe, T.; Sasahara, T. RFLP mapping of QTLs influencing shoot regeneration from mature seed-derived calli in rice. Crop. Sci. 2000, 40, 245–247. [Google Scholar] [CrossRef]
- Kwon, Y.S.; Eun, M.Y.; Sohn, J.K. Marker-assisted selection for identification of plant regeneration ability of seed-derived calli in rice (Oryza sativa L.). Mol. Cells 2001, 12, 103–106. [Google Scholar]
- Nishimura, A.; Ashikari, M.; Lin, S.; Takashi, T.; Angeles, E.R.; Yamamoto, T.; Matsuoka, M. Isolation of a rice regeneration quantitative trait loci gene and its application to transformation systems. Proc. Natl. Acad. Sci. USA 2005, 102, 11940–11944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Zhou, H.; Lu, L.; Liu, L.; Li, X.; Lin, Y.; Yu, S. Identification of quantitative trait loci controlling rice mature seed culturability using chromosomal segment substitution lines. Plant Cell Rep. 2009, 28, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.-K.; Ruan, B.-P.; Yan, M.X.; Ye, S.F.; Peng, Y.L.; Dong, G.J.; Zhu, L.; Hu, J.; Yan, H.L.; Guo, L.B.; et al. Genetic analysis and QTl mapping of mature seed culturability in indica rice. Rice Sci. 2013, 20, 313–319. [Google Scholar] [CrossRef]
- Bolibok, H.; Rakoczy-Trojanowska, M. Genetic mapping of QTLs for tissue-culture response in plants. Euphytica 2006, 149, 73–83. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhao, H.; Li, W.; Wu, J.; Zhou, Z.; Zhou, F.; Chen, H.; Lin, Y. Genome-wide association study of callus induction variation to explore the callus formation mechanism of rice. J. Integr. Plant Biol. 2019, 61, 1134–1150. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Su, J.; Xu, M.; Zhou, Z.; Zhu, X.; Ma, X.; Hou, J.; Tan, L.; Zhu, Z.; Cai, H.; et al. A common wild rice-derived BOC1 allele reduces callus browning in indica rice transformation. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, C.L.; Romero-Severson, J.; Hodges, T.K. Improved tissue culture response of an elite maize inbred through backcross breeding, and identification of chromosomal regions important for regeneration by RFLP analysis. Theor. Appl. Genet. 1992, 84, 755–762. [Google Scholar] [CrossRef]
- Ma, L.; Liu, M.; Yan, Y.; Qing, C.; Zhang, X.; Zhang, Y.; Long, Y.; Wang, L.; Pan, L.; Zou, C.; et al. Genetic Dissection of Maize Embryonic Callus Regenerative Capacity Using Multi-Locus Genome-Wide Association Studies. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvo, S.; Cook, J.; Carlson, A.R.; Hirsch, C.N.; Kaeppler, S.M.; Kaeppler, H.F. Genetic Fine-Mapping of a Quantitative Trait Locus (QTL) Associated with Embryogenic Tissue Culture Response and Plant Regeneration Ability in Maize (Zea mays L.). Plant Genome 2018, 11, 170111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Amer, I.M.; Korzun, V.; Worland, A.J.; Börner, A. Genetic mapping of QTL controlling tissue-culture response on chromosome 2B of wheat (Triticum aestivum L.) in relation to major genes and RFLP markers. Theor. Appl. Genet. 1997, 94, 1047–1052. [Google Scholar] [CrossRef]
- Jia, H.; Yi, D.; Yu, J.; Xue, S.; Xiang, Y.; Zhang, C.; Zhang, Z.; Zhang, L.; Ma, Z. Mapping QTLs for tissue culture response of mature wheat embryos. Mol. Cells 2007, 23, 323–330. [Google Scholar]
- Ma, J.; Deng, M.; Lv, S.Y.; Yang, Q.; Jiang, Q.T.; Qi, P.F.; Li, W.; Chen, G.Y.; Lan, X.J.; Wei, Y.M. Identification of QTLs associated with tissue culture response of mature wheat embryos. Springerplus 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Mano, Y.; Takahashi, H.; Sato, K.; Takeda, K. Mapping genes for callus growth and shoot regeneration in barley (Hordeum vulgare L.). Breed. Sci. 1996, 46, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Mano, Y.; Komatsuda, T. Identification of QTLs controlling tissue-culture traits in barley (Hordeum vulgare L.). Theor. Appl. Genet. 2002, 105, 708–715. [Google Scholar] [CrossRef]
- Tyagi, N.; Dahleen, L.S.; Bregitzer, P. Candidate genes within tissue culture regeneration QTL revisited with a linkage map based on transcript-derived markers. Crop. Sci. 2010, 50, 1697–1707. [Google Scholar] [CrossRef] [Green Version]
- Lowe, K.; Wu, E.; Wang, N.; Hoerster, G.; Hastings, C.; Cho, M.J.; Scelonge, C.; Lenderts, B.; Chamberlin, M.; Cushatt, J.; et al. Morphogenic regulators Baby boom and Wuschel improve monocot transformation. Plant Cell 2016, 28, 1998–2015. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, N.H.; Andersen, S.U.; Stougaard, J.; Jensen, A.; Backes, G.; Jahoor, A. Chromosomal regions associated with the in vitro culture response of wheat (Triticum aestivum L.) microspores. Plant Breed. 2015, 134, 255–263. [Google Scholar] [CrossRef]
- Hisano, H.; Sato, K. Genomic regions responsible for amenability to Agrobacterium-mediated transformation in barley. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hisano, H.; Meints, B.; Moscou, M.J.; Cistue, L.; Echávarri, B.; Sato, K.; Hayes, P.M. Selection of transformation-efficient barley genotypes based on TFA (transformation amenability) haplotype and higher resolution mapping of the TFA loci. Plant Cell Rep. 2017, 36, 611–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koornneef, M.; Bade, J.; Hanhart, C.; Horsman, K.; Schel, J.; Soppe, W.; Verkerk, R.; Zabel, P. Characterization and mapping of a gene controlling shoot regeneration in tomato. Plant J. 1993, 3, 131–141. [Google Scholar] [CrossRef]
- Satoh, H.; Takashina, T.; Escalante, A.; Egashira, H.; Imanishi, S. Molecular Markers Mapped around the High Shoot Regeneration Capacity Gene Rg-2 in Lycopersicon chilense. Breed. Sci. 2000, 50, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Trujillo-Moya, C.; Gisbert, C.; Vilanova, S.; Nuez, F. Localization of QTLs for in vitro plant regeneration in tomato. BMC Plant Biol. 2011, 11, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombardi-Crestana, S.; da Silva Azevedo, M.; e Silva, G.F.F.; Pino, L.E.; Appezzato-da-Glória, B.; Figueira, A.; Nogueira, F.T.S.; Peres, L.E.P. The tomato (Solanum lycopersicum cv Micro-Tom) natural methylation and chromatin patterning genetic variation Rg1 and the DELLA mutant procera control the competence necessary to form adventitious roots and shoots. J. Exp. Bot. 2012, 63, 5689–5703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhou, Q.; Zhu, G.; Wang, S.; Ma, Y.; Miao, H.; Zhang, S.; Huang, S.; Zhang, Z.; Gu, X. Genetic analysis and identification of a candidate gene associated with in vitro regeneration ability of cucumber. Theor. Appl. Genet. 2018, 131, 2663–2675. [Google Scholar] [CrossRef]
- Kim, K.; Kang, Y.; Lee, S.J.; Choi, S.H.; Jeon, D.H.; Park, M.Y.; Park, S.; Lim, Y.P.; Kim, C. Quantitative trait loci (QTLs) associated with microspore culture in Raphanus sativus L. (radish). Genes 2020, 11, 337. [Google Scholar] [CrossRef] [Green Version]
- Holme, I.B.; Torp, A.M.; Hansen, L.N.; Andersen, S.B. Quantitative trait loci affecting plant regeneration from protoplasts of Brassica oleracea. Theor. Appl. Genet. 2004, 108, 1513–1520. [Google Scholar] [CrossRef]
- Van Sint, J.; Laublin, G.; Birhman, R.K.; Cappadocia, M. Genetic analysis of leaf explant regenerability in Solanum chacoense. Plant Cell. Tissue Organ. Cult. 1996, 47, 9–13. [Google Scholar] [CrossRef]
- Nguyen, T.H.N.; Schulz, D.; Winkelmann, T.; Debener, T. Genetic dissection of adventitious shoot regeneration in roses by employing genome-wide association studies. Plant Cell Rep. 2017, 36, 1493–1505. [Google Scholar] [CrossRef] [PubMed]
- Flores Berrios, E.; Sarrafi, A.; Fabre, F.; Alibert, G.; Gentzbittel, L. Genotypic variation and chromosomal location of QTLs for somatic embryogenesis revealed by epidermal layers culture of recombinant inbred lines in the sunflower (Helianthus annuus L.). Theor. Appl. Genet. 2000, 101, 1307–1312. [Google Scholar] [CrossRef]
- Flores Berrios, E.; Gentzbittel, L.; Kayyal, H.; Alibert, G.; Sarrafi, A. AFLP mapping of QTLS for in vitro organogenesis traits, using recombinant inbred lines in sunflower (Helianthus annuus L.). Theor. Appl. Genet. 2000, 101, 1299–1306. [Google Scholar] [CrossRef]
- Tuskan, G.A.; Mewalal, R.; Gunter, L.E.; Palla, K.J.; Carter, K.; Jacobson, D.A.; Jones, P.C.; Garcia, B.J.; Weighill, D.A.; Hyatt, P.D.; et al. Defining the genetic components of callus formation: A GWAS approach. PLoS ONE 2018, 13, e0202519. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.; Jia, H.; Zhang, Y.; Li, J.; Lu, M.; Hu, J. Deciphering genetic architecture of adventitious root and related shoot traits in populus using qtl mapping and RNA-seq data. Int. J. Mol. Sci. 2019, 20, 6114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.; Park, O.; Seo, P.J. RNA-Seq Analysis of the Arabidopsis Transcriptome in Pluripotent Calli. Mol. Cells 2016, 39, 484–494. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.Y.; Latzel, V.; Fischer, M.; Bossdorf, O. Understanding the evolutionary potential of epigenetic variation: A comparison of heritable phenotypic variation in epiRILs, RILs, and natural ecotypes of Arabidopsis thaliana. Heredity (Edinb) 2018, 121, 257–265. [Google Scholar] [CrossRef]
- Schmid, M.W.; Heichinger, C.; Coman Schmid, D.; Guthörl, D.; Gagliardini, V.; Bruggmann, R.; Aluri, S.; Aquino, C.; Schmid, B.; Turnbull, L.A.; et al. Contribution of epigenetic variation to adaptation in Arabidopsis. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Becker, C.; Hagmann, J.; Müller, J.; Koenig, D.; Stegle, O.; Borgwardt, K.; Weigel, D. Spontaneous epigenetic variation in the Arabidopsis thaliana methylome. Nature 2011, 480, 245–249. [Google Scholar] [CrossRef]
- Kawakatsu, T.; Huang, S.C.; Jupe, F.; Sasaki, E.; Schmitz, R.J.J.; Urich, M.A.A.; Castanon, R.; Nery, J.R.R.; Barragan, C.; He, Y.; et al. Epigenomic Diversity in a Global Collection of Arabidopsis thaliana Accessions. Cell 2016, 166, 492–505. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Shiu, S.; Cal, A.; Borevitz, J.O. Global analysis of genetic, epigenetic and transcriptional polymorphisms in Arabidopsis thaliana using whole genome tiling arrays. PLoS Genet. 2008, 4. [Google Scholar] [CrossRef]
- Vaughn, M.W.; Tanurdžić, M.; Lippman, Z.; Jiang, H.; Carrasquillo, R.; Rabinowicz, P.D.; Dedhia, N.; McCombie, W.R.; Agier, N.; Bulski, A.; et al. Epigenetic natural variation in Arabidopsis thaliana. PLoS Biol. 2007, 5, 1617–1629. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wendte, J.M.; Ji, L.; Schmitz, R.J. Natural variation in DNA methylation homeostasis and the emergence of epialleles. Proc. Natl. Acad. Sci. USA 2020, 117, 4874–4884. [Google Scholar] [CrossRef] [PubMed]
- Zhai, J.; Liu, J.; Liu, B.; Li, P.; Meyers, B.C.; Chen, X.; Cao, X. Small RNA-directed epigenetic natural variation in Arabidopsis thaliana. PLoS Genet. 2008, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pignatta, D.; Erdmann, R.M.; Scheer, E.; Picard, C.L.; Bell, G.W.; Gehring, M. Natural epigenetic polymorphisms lead to intraspecific variation in Arabidopsis gene imprinting. Elife 2014, 3, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kooke, R.; Johannes, F.; Wardenaar, R.; Becker, F.; Etcheverry, M.; Colot, V.; Vreugdenhil, D.; Keurentjes, J.J.B. Epigenetic basis of morphological variation and phenotypic plasticity in Arabidopsis thaliana. Plant Cell 2015, 27, 337–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, Z.; Xie, S.; Zhu, J.K. The 1001 Arabidopsis DNA Methylomes: An Important Resource for Studying Natural Genetic, Epigenetic, and Phenotypic Variation. Trends Plant Sci. 2016, 21, 906–908. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Reimer, J.; Göbel, U.; Engelhorn, J.; He, F.; Schoof, H.; Turck, F. Natural variation of H3K27me3 distribution between two Arabidopsis accessions and its association with flanking transposable elements. Genome Biol. 2012, 13, R117. [Google Scholar] [CrossRef] [Green Version]
- Candela, M.; Velázquez, I.; De la Cruz, B.; Sendino, A.M.; De la Peña, A. Differences in in vitro plant regeneration ability among four Arabidopsis thaliana ecotypes. Vitr. Cell. Dev. Biol. Plant. 2001, 37, 638–643. [Google Scholar] [CrossRef]
- Zhang, X.; Hause, R.J.; Borevitz, J.O.; Borevitz, J.O. Natural Genetic Variation for Growth and Development Revealed by High-Throughput Phenotyping in Arabidopsis thaliana. G3 (Bethesda) 2012, 2, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Atwell, S.; Huang, Y.S.; Vilhjálmsson, B.J.; Willems, G.; Horton, M.; Li, Y.; Meng, D.; Platt, A.; Tarone, A.M.; Hu, T.T.; et al. Genome-wide association study of 107 phenotypes in Arabidopsis thaliana inbred lines. Nature 2010, 465, 627–631. [Google Scholar] [CrossRef] [PubMed]
- Cary, A.; Uttamchandani, S.J.; Smets, R.; Van Onckelen, H.A.; Howell, S.H. Arabidopsis mutants with increased organ regeneration in tissue culture are more competent to respond to hormonal signals. Planta 2001, 213, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, A.; Hosseini, S.A.; Hajirezaei, M.-R.; Druege, U.; Geelen, D. Adventitious rooting declines with the vegetative to reproductive switch and involves a changed auxin homeostasis. J. Exp. Bot. 2015, 66, 1437–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Sergeeva, L.I.; Vreugdenhil, D. Natural variation of hormone levels in Arabidopsis roots and correlations with complex root architecture. J. Integr. Plant Biol. 2018, 60, 292–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatfield, S.P.; Raizada, M.N. Ethylene and shoot regeneration: Hookless1 modulates de novo shoot organogenesis in Arabidopsis thaliana. Plant Cell Rep. 2008, 27, 655–666. [Google Scholar] [CrossRef] [PubMed]
- Delker, C.; Pöschschl, Y.; Raschke, A.; Ullrich, K.; Ettingshausen, S.; Hauptmann, V.; Grosse, I.; Quint, M. Natural variation of transcriptional auxin response networks in Arabidopsis thaliana. Plant Cell 2010, 22, 2184–2200. [Google Scholar] [CrossRef] [Green Version]
- Durgaprasad, K.; Roy, M.V.; Venugopal, A.; Kareem, A.; Raj, K.; Willemsen, V.; Mähönen, A.P.; Scheres, B.; Prasad, K. Gradient Expression of Transcription Factor Imposes a Boundary on Organ Regeneration Potential in Plants. Cell Rep. 2019, 29, 453–463.e3. [Google Scholar] [CrossRef] [Green Version]
- Togninalli, M.; Seren, Ü.; Meng, D.; Fitz, J.; Nordborg, M.; Weigel, D.; Borgwardt, K.; Korte, A.; Grimm, D.G. The AraGWAS Catalog: A curated and standardized Arabidopsis thaliana GWAS catalog. Nucleic Acids Res. 2018, 46, D1150–D1156. [Google Scholar] [CrossRef]
- Seren, Ü.; Grimm, D.; Fitz, J.; Weigel, D.; Nordborg, M.; Borgwardt, K.; Korte, A. AraPheno: A public database for Arabidopsis thaliana phenotypes. Nucleic Acids Res. 2017, 45, 1054–1059. [Google Scholar] [CrossRef] [Green Version]
- Exposito-Alonso, M.; Exposito-Alonso, M.; Gómez Rodríguez, R.; Barragán, C.; Capovilla, G.; Chae, E.; Devos, J.; Dogan, E.S.; Friedemann, C.; Gross, C.; et al. Natural selection on the Arabidopsis thaliana genome in present and future climates. Nature 2019, 573, 126–129. [Google Scholar] [CrossRef]
- Julkowska, M.M.; Koevoets, I.T.; Mol, S.; Hoefsloot, H.; Feron, R.; Tester, M.A.; Keurentjes, J.J.B.; Korte, A.; Haring, M.A.; De Boer, G.J.; et al. Genetic components of root architecture remodeling in response to salt stress. Plant Cell 2017, 29, 3198–3213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strauch, R.C.; Svedin, E.; Dilkes, B.; Chapple, C.; Li, X. Discovery of a novel amino acid racemase through exploration of natural variation in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2015, 112, 11726–11731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nameth, B.; Dinka, S.J.; Chatfield, S.P.; Morris, A.; English, J.; Lewis, D.; Oro, R.; Raizada, M.N. The shoot regeneration capacity of excised Arabidopsis cotyledons is established during the initial hours after injury and is modulated by a complex genetic network of light signalling. Plant Cell Environ. 2013, 36, 68–86. [Google Scholar] [CrossRef] [PubMed]
- Negrutiu, I.; Jacobs, M. Factors which Enhance in vitro Morphogenesis of Arabidopsis thaliana. Z. Pflanzenphysiol. 1978, 90, 423–430. [Google Scholar] [CrossRef]
- Gupta, S.K.; Singh, P.K.; Sawant, S.V.; Chaturvedi, R.; Tuli, R. Effect of light intensity on in vitro multiple shoot induction and regeneration of cotton (Gossypium hirsutum L. cv Khandawa-2). Indian J. Exp. Biol. 2000, 38, 399–401. [Google Scholar]
- Kwon, A.R.; Cui, H.Y.; Lee, H.; Shin, H.; Kang, K.S.; Park, S.Y. Light quality affects shoot regeneration, cell division, and wood formation in elite clones of Populus euramericana. Acta Physiol. Plant. 2015, 37, 1–9. [Google Scholar] [CrossRef]
- Reuveni, M.; Evenor, D. On the effect of light on shoot regeneration in petunia. Plant Cell. Tissue Organ. Cult. 2007, 89, 49–54. [Google Scholar] [CrossRef]
- Bhatia, P.; Ashwath, N. Effect of duration of light: Dark cycles on in vitro shoot regeneration of tomato. Asian J. Plant Sci. 2005, 3, 255–260. [Google Scholar]
- Dimasi-Theriou, K.; Economou, A. Effect of Cold Temperature on Shoot Regeneration in Vitro from Aged Cultures of GF-677 (Prunus Persica × Prunus Amygdalus). In Plant Aging; Springer US: Boston, MA, USA, 1990; pp. 345–349. [Google Scholar]
- Lee, S.-T.; Huang, W.-L. Osmotic stress stimulates shoot organogenesis in callus of rice (Oryza sativa L.) via auxin signaling and carbohydrate metabolism regulation. Plant Growth Regul. 2014, 73, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Wilson, M.E.; Mixdorf, M.; Berg, R.H.; Haswell, E.S. Plastid osmotic stress influences cell differentiation at the plant shoot apex. Development 2016, 143, 3382–3393. [Google Scholar] [CrossRef] [Green Version]
- Piwowarczyk, B.; Kamińska, I.; Rybiński, W. Influence of PEG Generated Osmotic Stress on Shoot Regeneration and Some Biochemical Parameters in Lathyrus Culture. Orig. Pap. Czech. J. Genet. Plant Breed. 2014, 50, 2014–2077. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Tan, H.J.; Liu, Y.B.; Li, X.R.; Chen, G.X. Effect of salt stress on growth and osmotic regulation in Thellungiella and Arabidopsis callus. Plant Cell. Tissue Organ. Cult. 2009, 98, 97–103. [Google Scholar] [CrossRef]
- Gleddie, S. Plant regeneration from cell suspension cultures of Arabidopsis thaliana heynh. Plant Cell Rep. 1989, 8, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Dovzhenko, A.; Dal Bosco, C.; Meurer, J.; Koop, H.U. Efficient regeneration from cotyledon protoplasts in Arabidopsis thaliana. Protoplasma 2003, 222, 107–111. [Google Scholar] [CrossRef]
- Scholl, R.L.; Keathley, D.E.; Baribault, T.J. Enhancement of Root Formation and Fertility in Shoots Regenerated From Anther- and Seedling-Derived Callus Cultures of Arabidopsis thaliana. Z. Pflanzenphysiol. 1981, 104, 225–231. [Google Scholar] [CrossRef]
- Paulraj, S.; Yeung, E.C. Improved shoot regeneration from root explants using an abscisic acid-containing medium. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2012; Volume 877, pp. 183–189. ISBN 9781617798177. [Google Scholar]
- Gordon-Kamm, B.; Sardesai, N.; Arling, M.; Lowe, K.; Hoerster, G.; Betts, S.; Jones, T. Using Morphogenic Genes to Improve Recovery and Regeneration of Transgenic Plants. Plants 2019, 8, 38. [Google Scholar] [CrossRef] [Green Version]
- Manske, U.; Landsmann, J.; Dietz-Pfeilstetter, A. Comparison of different methods for the establishment of RNA silencing in plants. Plant Biotechnol. Rep. 2017, 11, 115–125. [Google Scholar] [CrossRef]
- Zhai, Z.; Sooksa-nguan, T.; Vatamaniuk, O.K. Establishing RNA interference as a reverse-genetic approach for gene functional analysis in protoplasts. Plant Physiol. 2009, 149, 642–652. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.S.; Fei, J.F.; Xie, Q.; Chua, N.H. A chemical-regulated inducible RNAi system in plants. Plant J. 2003, 34, 383–392. [Google Scholar] [CrossRef]
- Fukudome, A.; Goldman, J.S.; Finlayson, S.A.; Koiwa, H. Silencing Arabidopsis CARBOXYL-TERMINAL DOMAIN PHOSPHATASE-LIKE 4 induces cytokinin-oversensitive de novo shoot organogenesis. Plant J. 2018, 94, 799–812. [Google Scholar] [CrossRef] [Green Version]
- Hesami, M.; Naderi, R.; Tohidfar, M. Modeling and Optimizing Medium Composition for Shoot Regeneration of Chrysanthemum via Radial Basis Function-Non-dominated Sorting Genetic Algorithm-II (RBF-NSGAII). Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hesami, M.; Condori-Apfata, J.A.; Valderrama Valencia, M.; Mohammadi, M. Application of Artificial Neural Network for Modeling and Studying In Vitro Genotype-Independent Shoot Regeneration in Wheat. Appl. Sci. 2020, 10, 5370. [Google Scholar] [CrossRef]
- Hesami, M.; Naderi, R.; Tohidfar, M.; Yoosefzadeh-Najafabadi, M. Development of support vector machine-based model and comparative analysis with artificial neural network for modeling the plant tissue culture procedures: Effect of plant growth regulators on somatic embryogenesis of chrysanthemum, as a case study. Plant Methods 2020, 16, 112. [Google Scholar] [CrossRef]
- Kaur, P.; Gupta, R.C.; Dey, A.; Malik, T.; Pandey, D.K. Optimization of salicylic acid and chitosan treatment for bitter secoiridoid and xanthone glycosides production in shoot cultures of Swertia paniculata using response surface methodology and artificial neural network. BMC Plant Biol. 2020, 20, 225. [Google Scholar] [CrossRef]
- García-Pérez, P.; Lozano-Milo, E.; Landín, M.; Gallego, P.P. Combining medicinal plant in vitro culture with machine learning technologies for maximizing the production of phenolic compounds. Antioxidants 2020, 9, 210. [Google Scholar] [CrossRef] [Green Version]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lardon, R.; Geelen, D. Natural Variation in Plant Pluripotency and Regeneration. Plants 2020, 9, 1261. https://doi.org/10.3390/plants9101261
Lardon R, Geelen D. Natural Variation in Plant Pluripotency and Regeneration. Plants. 2020; 9(10):1261. https://doi.org/10.3390/plants9101261
Chicago/Turabian StyleLardon, Robin, and Danny Geelen. 2020. "Natural Variation in Plant Pluripotency and Regeneration" Plants 9, no. 10: 1261. https://doi.org/10.3390/plants9101261
APA StyleLardon, R., & Geelen, D. (2020). Natural Variation in Plant Pluripotency and Regeneration. Plants, 9(10), 1261. https://doi.org/10.3390/plants9101261