Variable Fall Climate Conditions on Carbon Assimilation and Spring Phenology of Young Peach Trees


Abstract
:1. Introduction
2. Results
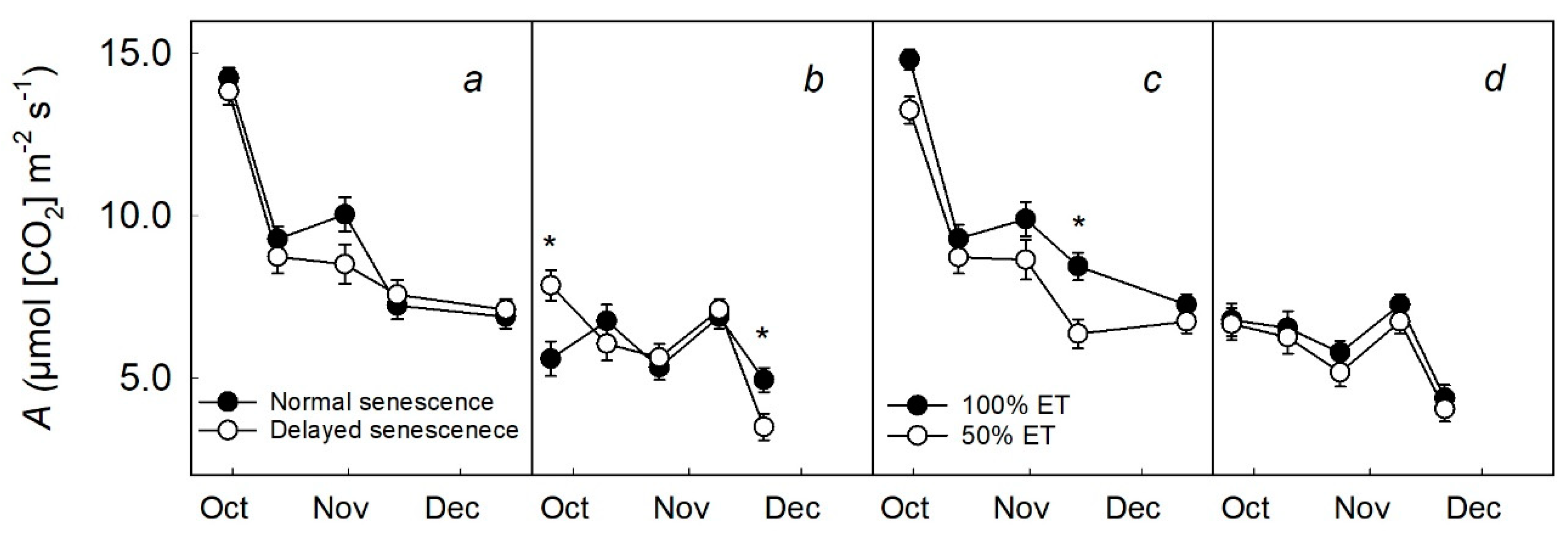
2.1. Carbon Assimilation
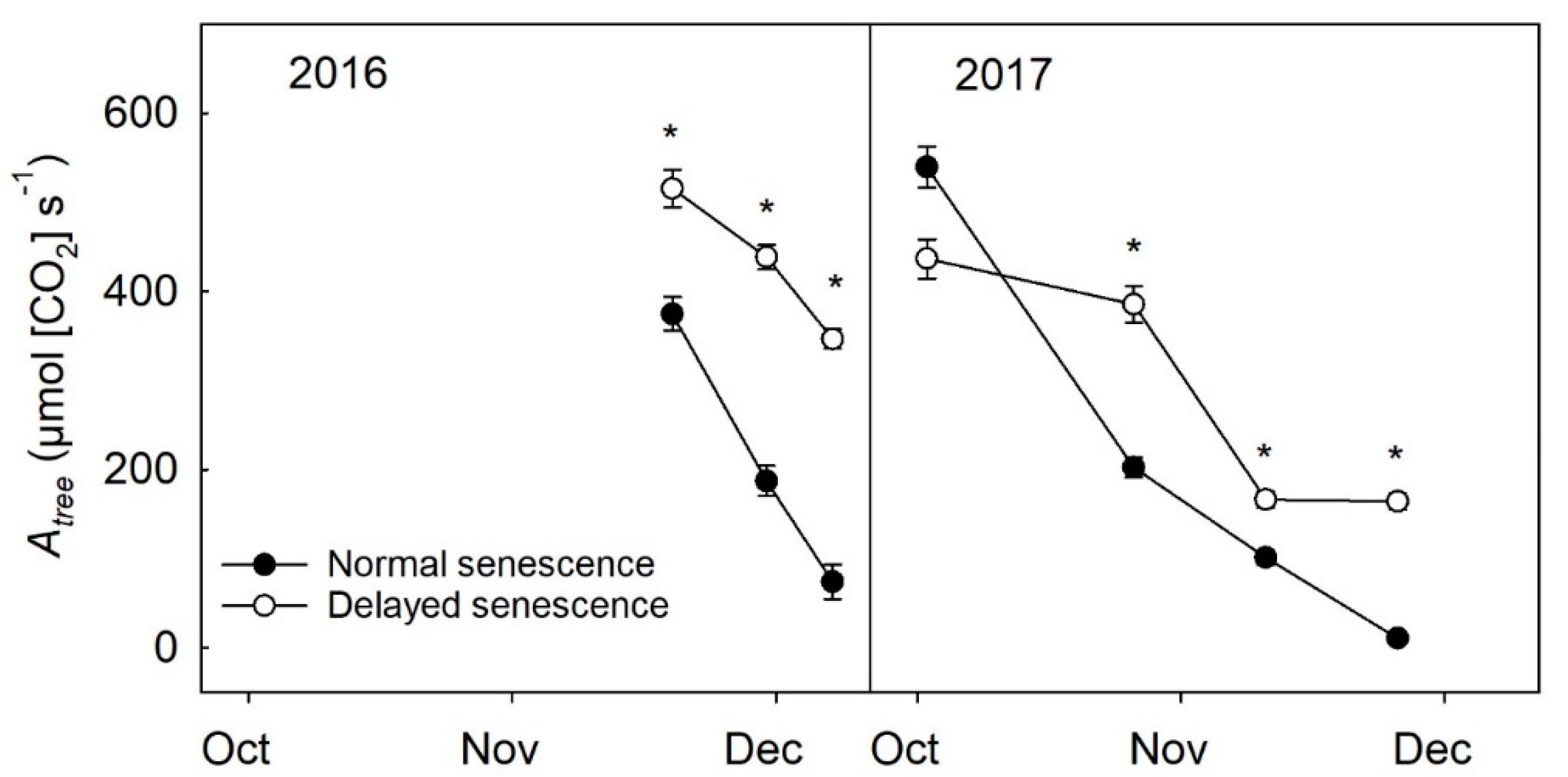
2.2. Leaf Count and Atree during Senescence
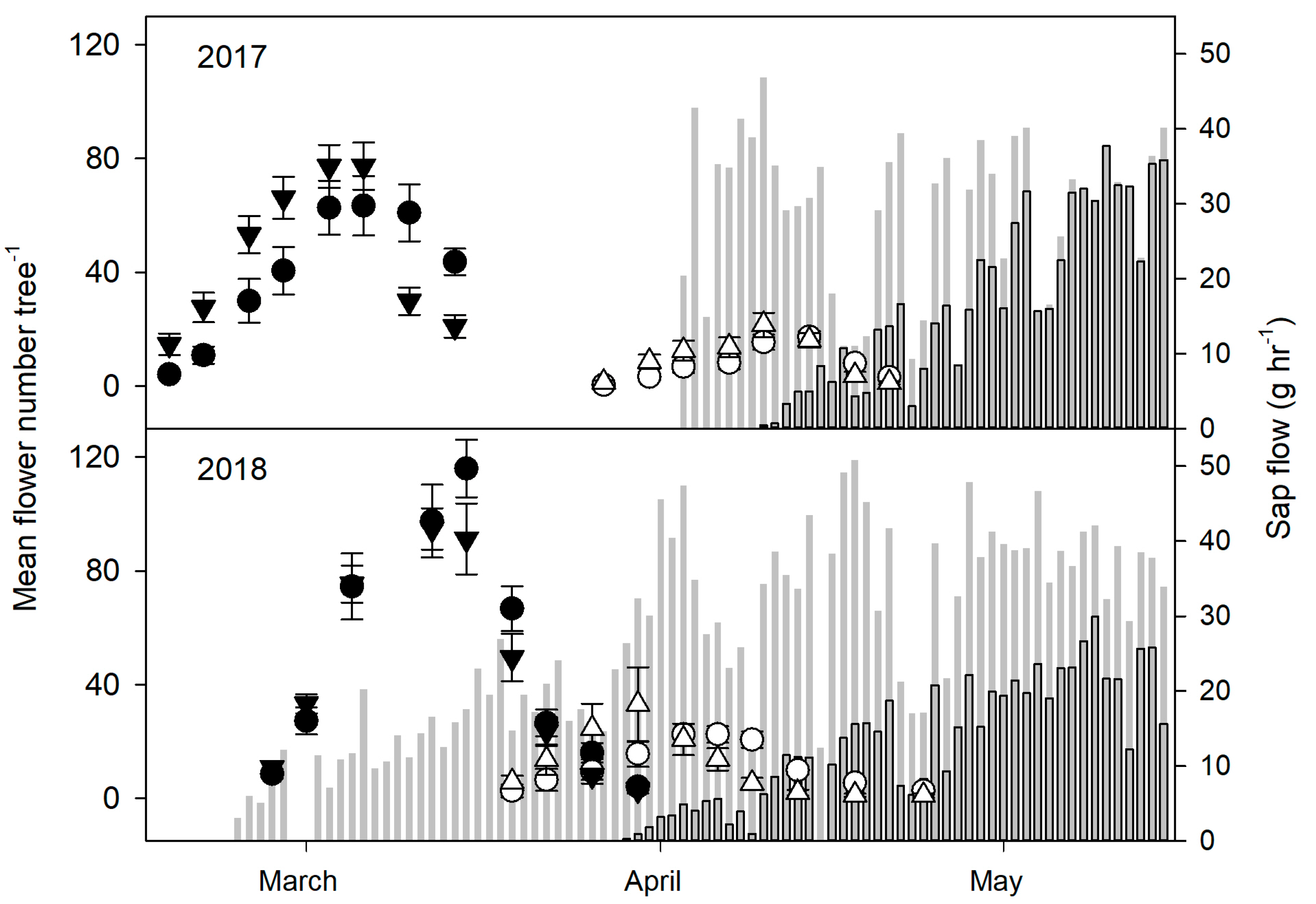
2.3. Biomass and Spring Phenology
3. Discussion
4. Materials and Methods
4.1. Study Location, Establishment and Design
4.2. Environmental and Gas Exchange Measurements
4.3. Biomass and Spring Phenology
4.4. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Badeck, F.W.; Bondeau, A.; Böttcher, K.; Doktor, D.; Lucht, W.; Schaber, J.; Sitch, S. Responses of spring phenology to climate change. New Phytol. 2004, 162, 295–309. [Google Scholar] [CrossRef]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. Forest Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Chmielewski, F.M.; Rötzer, T. Response of tree phenology to climate change across Europe. Agric. Forest Meteorol. 2001, 108, 101–112. [Google Scholar] [CrossRef]
- Way, D.A. Tree phenology responses to warming: Spring forward, fall back? Tree Physiol. 2011, 31, 469–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonhomme, M.; Rageau, R.; Lacointe, A.; Gendraud, M. Influences of cold deprivation during dormancy on carbohydrate contents of vegetative and floral primordia and nearby structures of peach buds (Prunus persica L. Batch). Sci. Hortic. 2005, 105, 223–240. [Google Scholar] [CrossRef]
- Charrier, G.; Lacointe, A.; Améglio, T. Dynamic modeling of carbon metabolism during the dormant period accurately predicts the changes in frost hardiness in walnut trees Juglans regia L. Front. Plant Sci. 2018, 9, 1746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luedeling, E. Climate change impacts on winter chill for temperate fruit and nut production: A review. Sci. Hortic. 2012, 144, 218–229. [Google Scholar] [CrossRef] [Green Version]
- Parker, L.E.; Abatzoglou, J.T. Warming winters reduce chill accumulation for peach production in the Southeastern United States. Climate 2019, 7, 94. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.H.; Piao, S.; Delpierre, N.; Hao, F.; Hänninen, H.; Liu, Y.; Sun, W.; Janssens, I.A.; Campioli, M. Larger temperature response of autumn leaf senescence than spring leaf-out phenology. Glob. Chang. Biol. 2018, 24, 2159–2168. [Google Scholar] [CrossRef]
- Gill, A.L.; Gallinat, A.S.; Sanders-DeMott, R.; Rigden, A.J.; Short Gianotti, D.J.; Mantooth, J.A.; Templer, P.H. Changes in autumn senescence in northern hemisphere deciduous trees: A meta-analysis of autumn phenology studies. Ann. Bot. 2015, 116, 875–888. [Google Scholar] [CrossRef] [Green Version]
- Gunderson, C.A.; Edwards, N.T.; Walker, A.V.; O’Hara, K.H.; Campion, C.M.; Hanson, P.J. Forest phenology and a warmer climate–growing season extension in relation to climatic provenance. Glob. Chang. Biol. 2012, 18, 2008–2025. [Google Scholar] [CrossRef]
- Estiarte, M.; Peñuelas, J. Alteration of the phenology of leaf senescence and fall in winter deciduous species by climate change: Effects on nutrient proficiency. Glob. Chang. Biol. 2015, 21, 1005–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Wang, X.; Silander, J.A. Deciduous forest responses to temperature, precipitation, and drought imply complex climate change impacts. Proc. Natl. Acad. Sci. USA 2015, 112, 13585–13590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wibbe, M.L.; Blanke, M.M.; Lenz, F. Respiration of apple trees between leaf fall and leaf emergence. Environ. Exp. Bot. 1994, 24, 25–30. [Google Scholar] [CrossRef]
- Dragoni, D.; Schmid, H.P.; Wayson, C.A.; Potter, H.; Grimmond, C.S.B.; Randolph, J.C. Evidence of increased net ecosystem productivity associated with a longer vegetated season in a deciduous forest in south-central Indiana, USA. Glob. Chang. Biol. 2011, 17, 886–897. [Google Scholar] [CrossRef]
- Tixier, A.; Gambetta, G.A.; Godfrey, J.M.; Orozco, J.; Zwieniecki, M.A. Non-structural carbohydrates in dormant perennials; the tale of winter survival and spring arrival. Front. For. Glob. Chang. 2019, 2, 18. [Google Scholar] [CrossRef] [Green Version]
- Dichio, B.; Xiloyannis, C.; Sofo, A.; Montanaro, G. Effects of post-harvest regulated deficit irrigation on carbohydrate and nitrogen partitioning, yield quality and vegetative growth of peach trees. Plant Soil 2007, 290, 127. [Google Scholar] [CrossRef]
- Pérez-Pastor, A.; Domingo, R.; Torrecillas, A.; Ruiz-Sánchez, M.C. Response of apricot trees to deficit irrigation strategies. Irrig. Sci. 2009, 27, 231–242. [Google Scholar] [CrossRef]
- Goldhamer, D.A.; Viveros, M. Effects of preharvest irrigation cutoff durations and postharvest water deprivation on almond tree performance. Irrig. Sci. 2000, 19, 125–131. [Google Scholar] [CrossRef]
- Torrecillas, A.; Domingo, R.; Galego, R.; Ruiz-Sánchez, M.C. Apricot tree response to withholding irrigation at different phenological periods. Sci. Hortic. 2000, 85, 201–215. [Google Scholar] [CrossRef]
- Dong, B.; Dai, A. The influence of the interdecadal Pacific oscillation on temperature and precipitation over the globe. Clim. Dyn. 2015, 45, 2667–2681. [Google Scholar] [CrossRef]
- Jarvis, C.; Darbyshire, R.; Eckard, R.; Goodwin, I.; Barlow, E. Influence of El Niño-Southern oscillation and the Indian Ocean Dipole on winegrape maturity in Australia. Agric. For. Meteorol. 2018, 248, 502–510. [Google Scholar] [CrossRef]
- Jones, G.V.; Goodrich, G.B. Influence of climate variability on wine regions in the western USA and on wine quality in the Napa Valley. Clim. Res. 2008, 35, 241–254. [Google Scholar] [CrossRef]
- Uzun, A.; Ustaoğlu, B. Impacts of El Nino Southern Oscillation (ENSO) and North Atlantic Oscillation (NAO) on the olive yield in the mediterranean region, Turkey. In Proceedings of the 8th International Conference on Agro-Geoinformatics (Agro-Geoinformatics), Istanbul, Turkey, 16–19 July 2019; pp. 1–6. [Google Scholar] [CrossRef]
- Lawrence, B.T.; Melgar, J.C. Variable fall climate influences nutrient resorption and reserve storage in young peach trees. Front. Plant Sci. 2018, 9, 1819. [Google Scholar] [CrossRef] [Green Version]
- Gariglio, N.; Weber, M.; Castro, D.; Micheloud, N. Influence of the Environmental Conditions, the Variety, and Different Cultural Practices on the Phenology of Peach in the Central Area of Santa Fe (Argentina). In Phenology and Climate Change; Zhang, X., Ed.; InTech: Shanghai, China, 2012; pp. 217–240. [Google Scholar]
- Weber, M.E.; Pilatti, R.A.; Sordo, M.H.; García, M.S.; Gariglio, N.F. Changes in the vegetative growth of the low-chill peach tree in response to reproductive shoot pruning after harvesting. New Zeal. J. Crop Hort. 2011, 39, 153–160. [Google Scholar] [CrossRef]
- Archetti, M.; Richardson, A.D.; O’Keefe, J.; Delpierre, N. Predicting climate change impacts on the amount and duration of autumn colors in a New England forest. PLoS ONE 2013, 8, e57373. [Google Scholar] [CrossRef]
- Naschitz, S.; Naor, A.; Wolf, S.; Goldschmidt, E.E. The effects of temperature and drought on autumnal senescence and leaf shed in apple under warm, east mediterranean climate. Trees 2014, 28, 879–890. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Yang, Q.Y.; Lee, D.W.; Goldstein, G.; Cao, K.F. Extended leaf senescence promotes carbon gain and nutrient resorption: Importance of maintaining winter photosynthesis in subtropical forests. Oecologia 2013, 173, 721–730. [Google Scholar] [CrossRef]
- Bauerle, W.L.; Oren, R.; Way, D.A.; Qian, S.S.; Stoy, P.C.; Thornton, P.E.; Bowden, J.D.; Hoffman, F.M.; Reynolds, R.F. Photoperiodic regulation of the seasonal pattern of photosynthetic capacity and the implications for carbon cycling. Proc. Natl. Acad. Sci. USA 2012, 109, 8612–8617. [Google Scholar] [CrossRef] [Green Version]
- Charrier, G.; Améglio, T. The timing of leaf fall affects cold acclimation by interactions with air temperature through water and carbohydrate contents. Environ. Exp. Bot. 2011, 72, 351–357. [Google Scholar] [CrossRef]
- Schönbeck, L.; Gessler, A.; Hoch, G.; McDowell, N.G.; Rigling, A.; Schaub, M.; Li, M.H. Homeostatic levels of nonstructural carbohydrates after 13 yr of drought and irrigation in Pinus sylvestris. New Phytol. 2018, 219, 1314–1324. [Google Scholar] [CrossRef] [Green Version]
- Piao, S.; Ciais, P.; Friedlingstein, P.; Peylin, P.; Reichstein, M.; Luyssaert, S.; Margolis, H.; Fang, J.; Barr, A.; Chen, A.; et al. Net carbon dioxide losses of northern ecosystems in response to autumn warming. Nature 2008, 451, 49–52. [Google Scholar] [CrossRef]
- Collier, D.E.; Thibodeau, B.A. Changes in respiration and chemical content during autumnal senescence of Populus tremuloides and Quercus rubra leaves. Tree Physiol. 1995, 15, 759–764. [Google Scholar] [CrossRef]
- Luo, Z.; Guan, H.; Zhang, X.; Liu, N. Photosynthetic capacity of senescent leaves for a subtropical broadleaf deciduous tree species Liquidambar formosana Hance. Sci. Rep. 2017, 7, 6323. [Google Scholar] [CrossRef] [Green Version]
- Kalcsits, L.A.; Silim, S.; Tanino, K. Warm temperature accelerates short photoperiod-induced growth cessation and dormancy induction in hybrid poplar (Populus × spp.). Trees 2009, 23, 971–979. [Google Scholar] [CrossRef]
- Zhang, C.; Stratopoulos, L.M.F.; Pretzsch, H.; Rötzer, T. How do Tilia cordata Greenspire trees cope with drought stress regarding their biomass allocation and ecosystem services? Forests 2019, 10, 676. [Google Scholar] [CrossRef] [Green Version]
- Landeros, J.; Guevara, L.P.; Badii, M.H.; Flores, A.E.; Pámanes, A. Effect of different densities of the twospotted spider mite Tetranychus urticae on CO2 assimilation, transpiration, and stomatal behaviour in rose leaves. Exp. Appl. Acarol. 2004, 32, 187–198. [Google Scholar] [CrossRef]
- Campoy, J.A.; Ruiz, D.; Egea, J. Dormancy in temperate fruit trees in a global warming context: A review. Sci. Hortic. 2011, 130, 357–372. [Google Scholar] [CrossRef]
- Okie, W.R.; Blackburn, B. Interactive effects of light and chilling on peach flower and leaf budbreak. HortScience 2011, 46, 1056–1062. [Google Scholar] [CrossRef] [Green Version]
- Greer, D.H.; Wünsche, J.N. Late-season temperature effects on the carbon economy and tree performance of ‘Royal Gala’ apple (Malus domestica) trees. New Zeal. J. Crop Hort. 2003, 31, 235–245. [Google Scholar] [CrossRef]
- Romero-Conde, A.; Kusakabe, A.; Melgar, J.C. Physiological responses of citrus to partial rootzone drying irrigation. Sci. Hortic. 2014, 169, 234–238. [Google Scholar] [CrossRef]
- Pornon, A.; Lamaze, T. Nitrogen resorption and photosynthetic activity over leaf life span in an evergreen shrub, Rhododendron ferrugineum, in a subalpine environment. New Phytol. 2007, 175, 301–310. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Treatment | 2017 | 2018 | ||
|---|---|---|---|---|
| R:S | p | R:S | p | |
| NS | 0.53 a | <0.01 | 0.84 a | 0.08 |
| DS | 0.43 b | 0.75 a | ||
| 100% ET | 0.48 a | 0.95 | 0.78 a | 0.94 |
| 50% ET | 0.48 a | 0.79 a | ||
| Year and Treatment | Date and VPD (kPa) | |||||
|---|---|---|---|---|---|---|
| 2016 | September 30 | October 13 | October 31 | November 14 | December 13 | |
| NS | 2.32 ± 0.06 b | 2.02 ± 0.07 b | 2.12 ± 0.07 b | 1.46 ± 0.05 b | 1.24 ± 0.02 a | |
| DS | 2.80 ± 0.07 a | 2.47 ± 0.09 a | 2.68 ± 0.07 a | 2.00 ± 0.06 a | 1.00 ± 0.02 b | |
| 2017 | September 25 | October 10 | October 24 | November 9 | November 21 | |
| NS | 2.53 ± 0.08 a | 2.43 ± 0.02 b | 1.89 ± 0.06 b | 1.23 ± 0.04 b | 1.74 ± 0.03 b | |
| DS | 1.89 ± 0.07 b | 2.52 ± 0.02 a | 2.25 ± 0.06 a | 1.78 ± 0.05 a | 1.92 ± 0.03 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lawrence, B.T.; Melgar, J.C. Variable Fall Climate Conditions on Carbon Assimilation and Spring Phenology of Young Peach Trees. Plants 2020, 9, 1353. https://doi.org/10.3390/plants9101353
Lawrence BT, Melgar JC. Variable Fall Climate Conditions on Carbon Assimilation and Spring Phenology of Young Peach Trees. Plants. 2020; 9(10):1353. https://doi.org/10.3390/plants9101353
Chicago/Turabian StyleLawrence, Brian T., and Juan Carlos Melgar. 2020. "Variable Fall Climate Conditions on Carbon Assimilation and Spring Phenology of Young Peach Trees" Plants 9, no. 10: 1353. https://doi.org/10.3390/plants9101353
APA StyleLawrence, B. T., & Melgar, J. C. (2020). Variable Fall Climate Conditions on Carbon Assimilation and Spring Phenology of Young Peach Trees. Plants, 9(10), 1353. https://doi.org/10.3390/plants9101353