Identification and Analysis of NBS-LRR Genes in Actinidia chinensis Genome


Abstract
:1. Introduction
2. Results
2.1. Identification and Classification of Kiwifruit NBS-LRR Genes
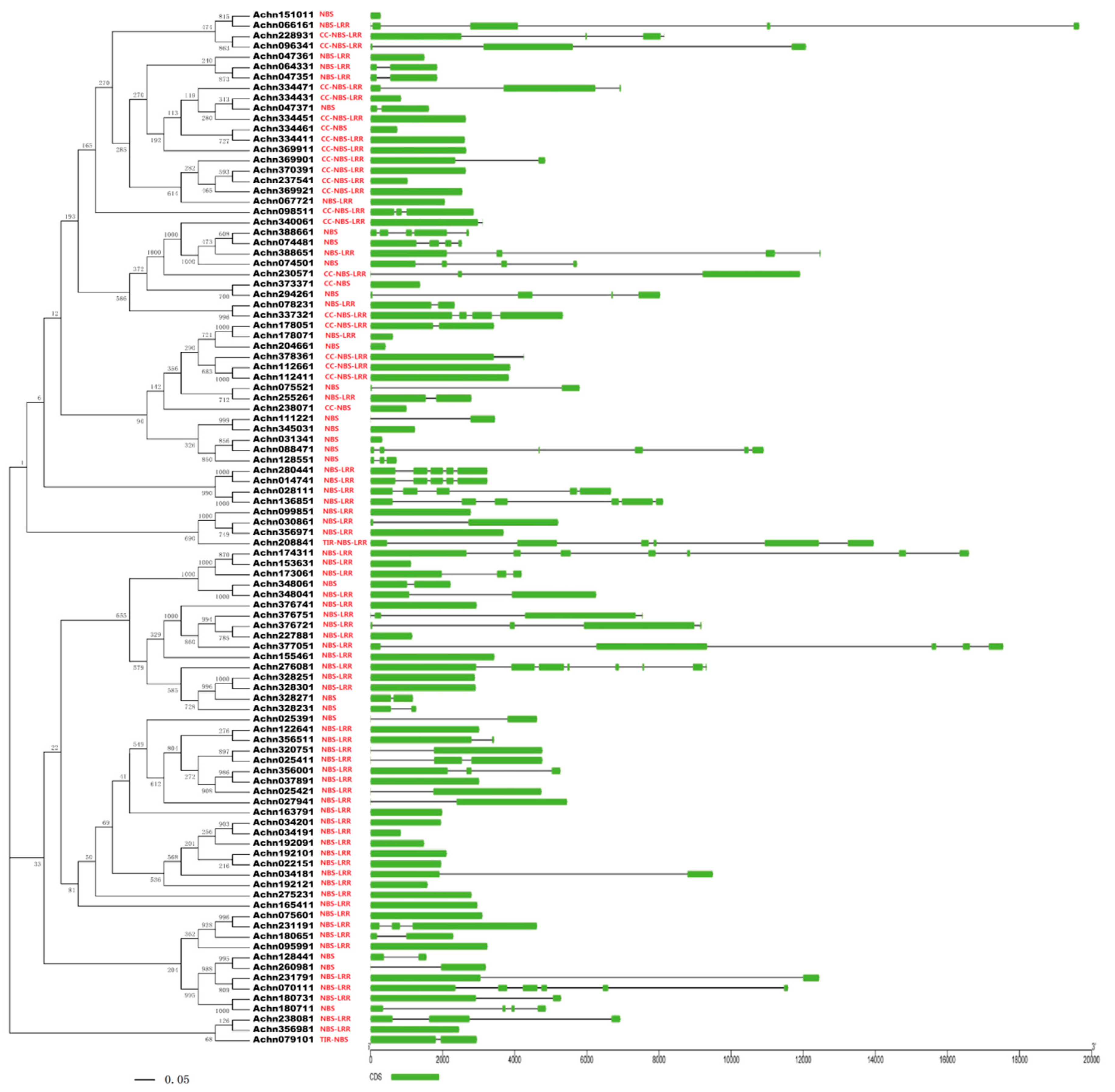
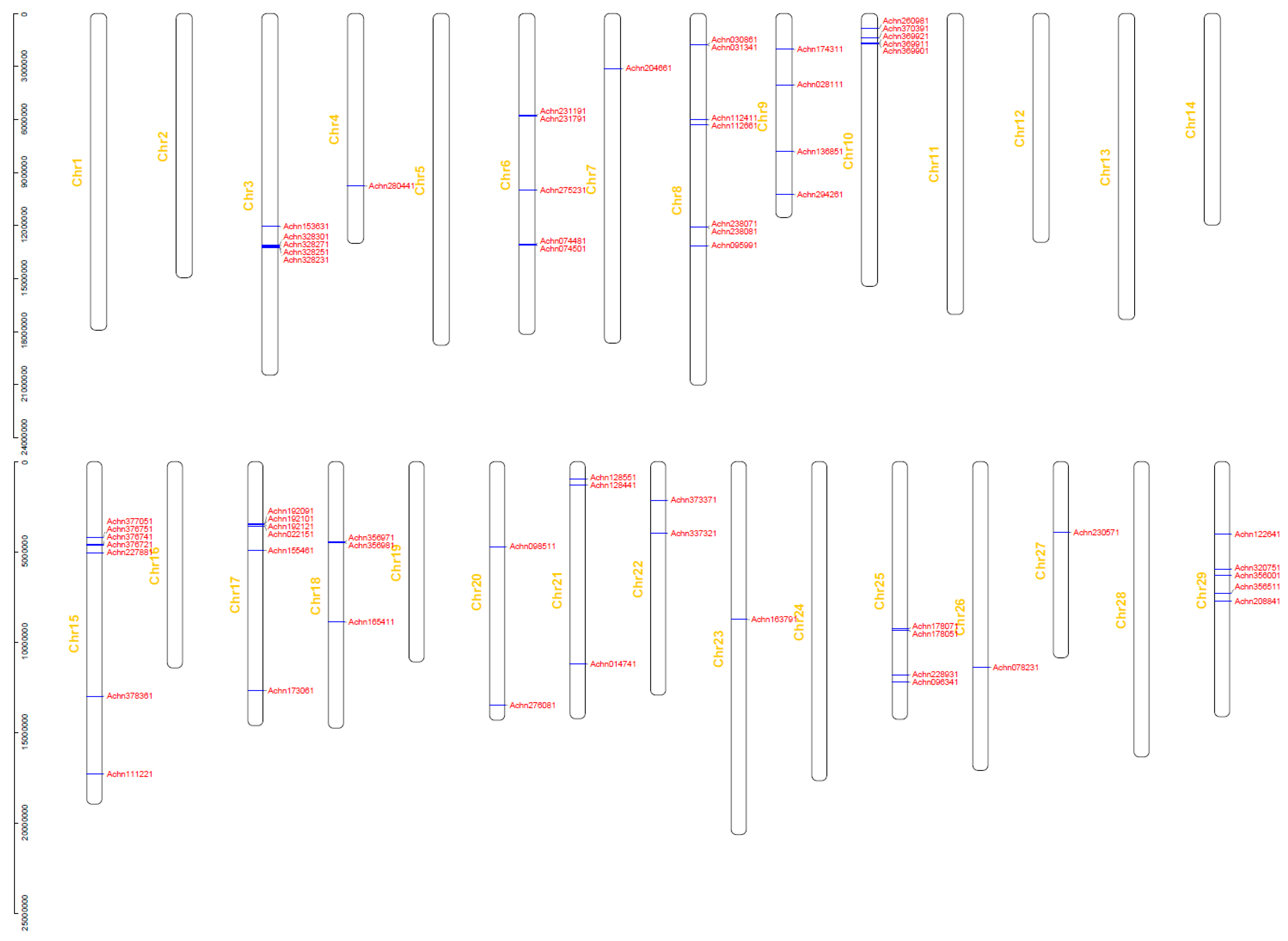
2.2. Phylogenetic Analysis, Gene Structural Characteristics, and Chromosome Localization
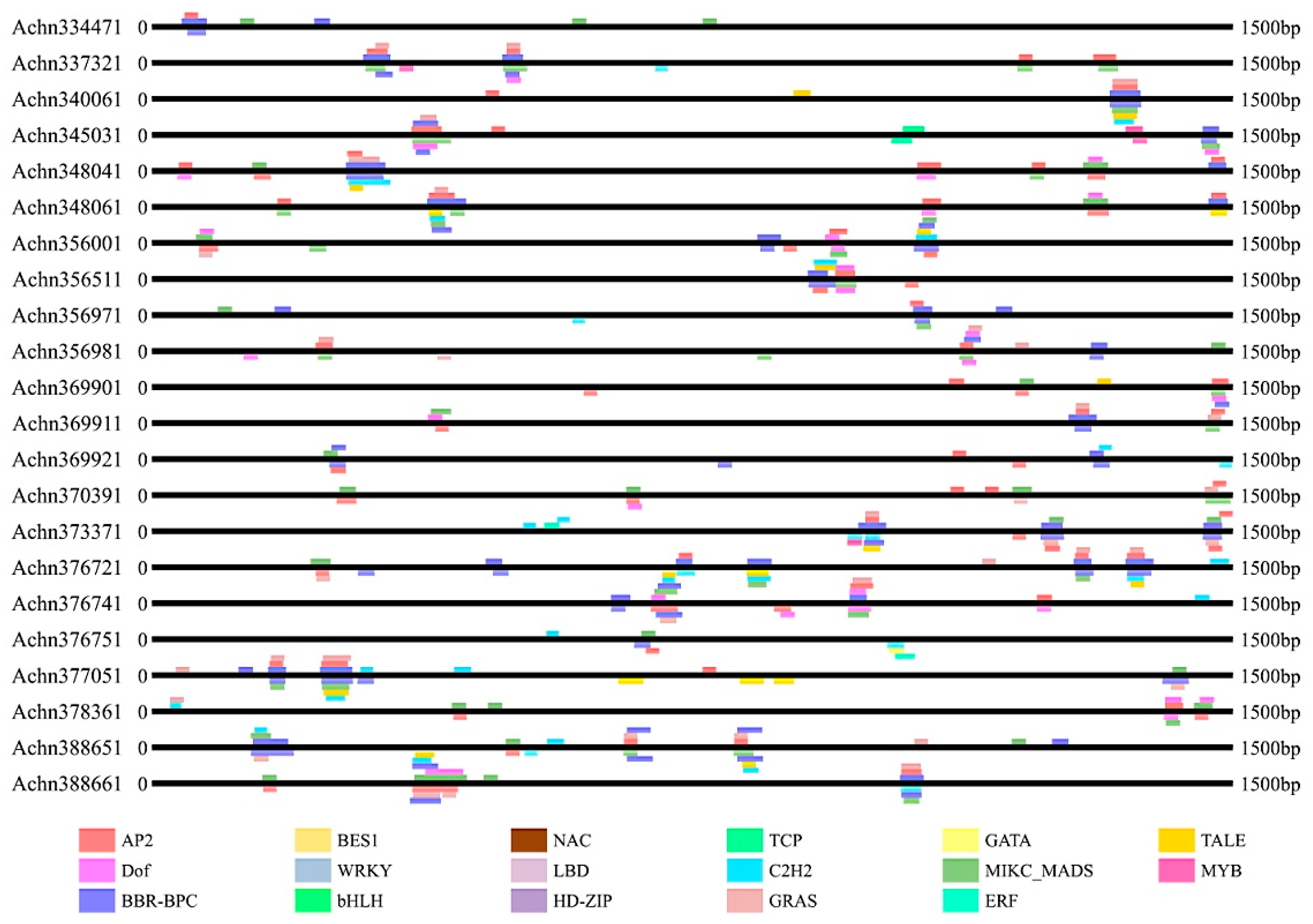
2.3. Cis-Acting Element Analysis and Subcellular Location Prediction
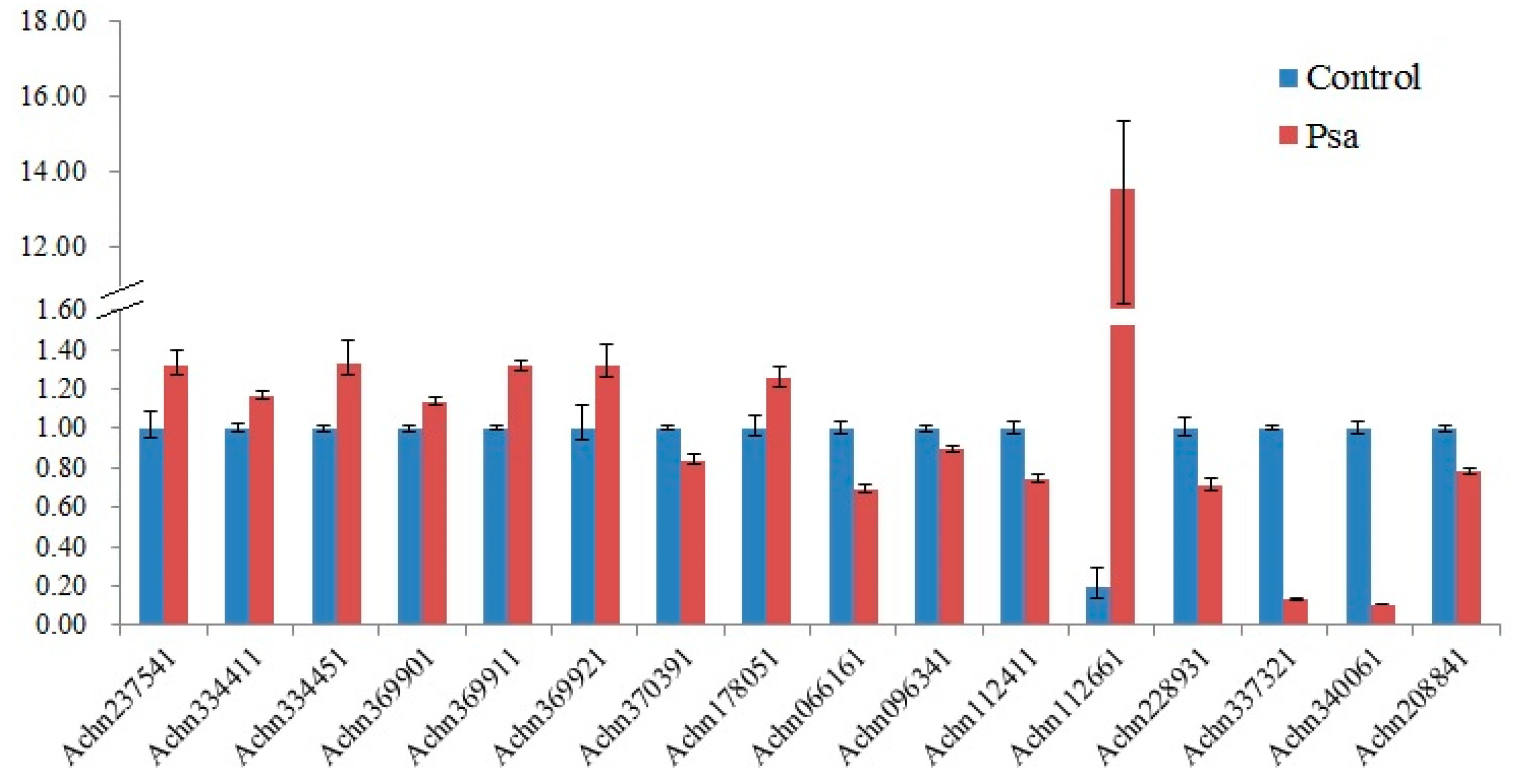
2.4. Expression Pattern of Kiwifruit NBS-LRR Genes under Psa Infection
3. Discussion
4. Materials and Methods
4.1. Identification and Classification of Kiwifruit NBS-LRR Genes
4.2. Phylogenetic Analysis, Gene Structure and Chromosome Localization
4.3. Cis-Acting Element Analysis and Subcellular Location Prediction
4.4. Inoculation of Psa and Quantitative RT-PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Li, W.; Ding, Z.; Ruan, M.; Yu, X.; Peng, M.; Liu, Y. Kiwifruit R2R3-MYB transcription factors and contribution of the novel AcMYB75 to red kiwifruit anthocyanin biosynthesis. Sci. Rep. 2017, 7, 16861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanneste, J.L.; Yu, J.; Cornish, D.A.; Tanner, D.J.; Windner, R.; Chapman, J.R.; Taylor, R.K.; Mackay, J.F.; Dowlut, S. Identification, virulence and distribution of two biovars of Pseudomonas syringae pv. actinidiae in New Zealand. Plant Dis. 2013, 97, 708–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balestra, G.M.; Renzi, M.; Mazzaglia, A. First report of bacterial canker of Actinidia deliciosa caused by Pseudomonas syringae pv. actinidiae in Portugal. New Dis. Rep. 2010, 22, 2510–2513. [Google Scholar] [CrossRef] [Green Version]
- Abelleira, A.; López, M.M.; Peñalver, J.; Aguín, O.; Mansilla, J.P.; Picoaga, A.; García, M.J. First report of bacterial canker of kiwifruit caused by Pseudomonas syringae pv. actinidiae in Spain. Plant Dis. 2011, 95, 1583. [Google Scholar] [CrossRef] [PubMed]
- Vanneste, J.L.; Poliakoff, F.; Audusseau, C.; Cornish, D.A.; Paillard, S.; Rivoal, C.; Yu, J. First report of Pseudomonas syringae pv. actinidiae, the causal agent of bacterial canker of kiwifruit in France. Plant Dis. 2011, 95, 1311–1312. [Google Scholar] [CrossRef]
- Bastas, K.K.; Karakaya, A. First report of bacterial canker of kiwifruit caused by Pseudomonas syringae pv. actinidiae in Turkey. Plant Dis. 2012, 96, 452. [Google Scholar] [CrossRef]
- Holeva, M.C.; Glynos, P.E.; Karafla, C.D. First report of bacterial canker of kiwifruit caused by Pseudomonas syringae pv. actinidiae in Greece. Plant Dis. 2015, 99, 723. [Google Scholar] [CrossRef]
- Mondragon-Palomino, M. Patterns of positive selection in the complete NBS-LRR gene family of Arabidopsis thaliana. Genom Res 2002, 12, 1305–1315. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.J.; Kim, K.H.; Shim, S.; Yoon, M.Y.; Sun, S.; Kim, M.; Van, K.; Lee, S.-H. Genome-wide mapping of NBS-LRR genes and their association with disease resistance in soybean. BMC Plant Biol. 2012, 12, 139. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Wang, P.; Li, T.; Xia, H.; Zhao, C. Genome-wide identification and evolutionary analysis of nucleotide-binding site-encoding resistance genes in Lotus japonicus (Fabaceae). Genet. Mol. Res. 2015, 14, 16024–16040. [Google Scholar] [CrossRef]
- Surendra, N.; Andersen, E.J.; Achal, N.; Nepal, M.P. Genome-wide identification of nbs-encoding resistance genes in sunflower (Helianthus annuus L.). Genes 2018, 9, 384. [Google Scholar]
- Zhou, T.; Wang, Y.; Chen, J.Q.; Araki, H.; Jing, Z.; Jiang, K.; Shen, J.; Tian, D. Genome-wide identification of NBS genes in japonica rice reveals significant expansion of divergent non-TIR NBS-LRR genes. Mol. Genet. Genom. 2004, 271, 402–415. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Si, W.; Zhao, L.; Yang, S.; Zhang, X. Dynamic evolution of NBS-LRR genes in bread wheat and its progenitors. Mol. Genet. Genom. 2015, 290, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Marone, D.; Russo, M.A.M.; Laidò, G.; de Leonardis, A.M.; Mastrangelo, A.M. Plant nucleotide binding site-leucine-rich repeat (NBS-LRR) genes: Active guardians in host defense responses. Int. J. Mol. Sci. 2013, 14, 7302–7326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leipe, D.D.; Koonin, E.V.; Aravind, L. STAND, a class of P-Loop NTPases including animal and plant regulators of programmed cell death: Multiple, complex domain architectures, unusual phyletic patterns, and evolution by horizontal gene transfer. J. Mol. Biol. 2004, 343, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Takken, F.L.; Albrecht, M.; Tameling, W.I. Resistance proteins: Molecular switches of plant defence. Curr. Opin. Plant Bio. 2006, 9, 383–390. [Google Scholar]
- Elmore, J.M.; Lin, Z.J.D.; Coaker, G. Plant NB-LRR signaling: Upstreams and downstreams. Curr. Opin. Plant Bio. 2011, 14, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Qi, D.; Innes, R.W. Recent advances in plant NLR structure, function, localization, and signaling. Front. Immunol. 2013, 4, 348. [Google Scholar] [CrossRef] [Green Version]
- Inoue, H.; Hayashi, N.; Matsushita, A.; Xinqiong, L.; Nakayama, A.; Sugano, S.; Jiang, C.-J.; Takatsuji, H. Blast resistance of CC-NB-LRR protein Pb1 is mediated by WRKY45 through protein-protein interaction. Proc. Natl. Acad. Sci. USA 2013, 110, 9577–9582. [Google Scholar] [CrossRef] [Green Version]
- Changjiang, G.; Xiaoguang, S.; Xiao, C.; Sihai, Y.; Jing, L.; Long, W.; Xiaohui, Z. Cloning of novel rice blast resistance genes from two rapidly evolving NBS-LRR gene families in rice. Plant Mol. Biol. 2016, 90, 95–105. [Google Scholar]
- Stirnweis, D.; Milani, S.D.; Jordan, T.; Keller, B.; Brunner, S. Substitutions of two amino acids in the nucleotide-binding site domain of a resistance protein enhance the hypersensitive response and enlarge the PM3F resistance spectrum in wheat. Mol. Plant Microbe. Interact. 2014, 27, 265–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashikawa, I.; Ishikawa, R. A population genetics-based approach reveals candidate gene regions important in the determination of resistance specificity for the Pik family of rice blast resistance genes. J. Gen. Plant Pathol. 2013, 79, 178–181. [Google Scholar] [CrossRef]
- Harris, C.J.; Slootweg, E.J.; Goverse, A.; Baulcombe, D.C. Stepwise artificial evolution of a plant disease resistance gene. Proc. Natl. Acad. Sci. USA 2013, 110, 21189–21194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dangl, J.L.; Jones, J.D.G. Plant pathogens and integrated defence responses to infection. Nature 2001, 411, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Scortichini, M.; Marcelletti, S.; Ferrante, P.; Petriccione, M.; Firrao, G. Pseudomonas syringae pv. actinidiae: A re-emerging, multi-faceted, pandemic pathogen. Mol. Plant Pathol. 2012, 13, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Chung, E.H.; Cunha, L.D.; Wu, A.J.; Gao, Z.; Cherkis, K.; Afzal, A.J.; Mackey, D.; Dangl, J.L. Specific threonine phosphorylation of a host target by two unrelated type III effectors activates a host innate immune receptor in plants. Cell Host Microbe. 2011, 9, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Yiping, Q.I.; Tsuda, K.; Glazebrook, J. Physical association of pattern-triggered immunity (PTI) and effector-triggered immunity (ETI) immune receptors in Arabidopsis. Mol. Plant Pathol. 2011, 12, 702–708. [Google Scholar]
- Ming, R.; Hou, S.B.; Feng, Y.; Yu, Q.Y.; Dionne-Laporte, A.; Saw, J.H.; Senin, P.; Wang, W.; Ly, B.V.; Lewis, K.L.T.; et al. The draft genome of the transgenic tropical fruit tree papaya (Carica papaya Linnaeus). Nature 2008, 452, U991–U997. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Yin, H.; Sargent, D.J.; Malnoy, M.; Cheng, Z.M. Species-specific duplications driving the recent expansion of NBS–LRR genes in five Rosaceae species. BMC Genom. 2015, 16, 77. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Zhang, X.; Yue, J.X.; Tian, D.; Chen, J.Q. Recent duplications dominate NBS-encoding gene expansion in two woody species. Mol. Genet. Genomics. 2008, 280, 187–198. [Google Scholar] [CrossRef]
- Lozano, R.; Hamblin, M.T.; Prochnik, S.; Jannink, J.L. Identification and distribution of the NBS-LRR gene family in the Cassava genome. BMC Genom. 2015, 16, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyers, B.C.; Kozik, A.; Griego, A.; Kuang, H.H.; Michelmore, R.W. Genome-wide analysis of NBS-LRR encoding genes in Arabidopsis. Plant Cell 2003, 15, 809–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.; Nan, Z. Genome-wide analysis of nucleotide-binding site disease resistance genes in Medicago truncatula. Chin. Sci. Bull. 2014, 59, 1129–1138. [Google Scholar] [CrossRef]
- Ameline-Torregrosa, C.; Wang, B.B.; O’Bleness, M.S.; Deshpande, S.; Zhu, H.; Roe, B.; Young, N.D.; Cannon, S.B. Identification and characterization of nucleotide-binding site-leucine-rich repeat genes in the model plant Medicago truncatula. Plant Physiol. 2008, 146, 5–21. [Google Scholar] [CrossRef] [Green Version]
- Andolfo, G.; Sanseverino, W.; Aversano, R.; Frusciante, L.; Ercolano, M.R. Genome-wide identification and analysis of candidate genes for disease resistance in tomato. Mol. Breed. 2014, 33, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Jing, W.; Jifeng, Z.; Lanfen, W.; Shumin, W. Genome-wide association study identifies NBS-LRR-encoding genes related with anthracnose and common bacterial blight in the common bean. Front Plant Sci. 2017, 8, 8. [Google Scholar]
- Cannon, S.B.; May, G.D.; Jackson, S.A. Three sequenced legume genomes and many crop species: Rich opportunities for translational genomics. Plant Physiol. 2009, 151, 970–977. [Google Scholar] [CrossRef] [Green Version]
- Grant, M.R.; McDowell, J.M.; Sharpe, A.G.; de Torres Zabala, M.; Lydiate, D.J.; Dangl, J.L. Independent deletions of a pathogen-resistance gene in Brassica and Arabidopsis. Proc. Natl. Acad. Sci. USA 1998, 95, 15843–15848. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.W.; Ferguson, A.R. In China: Natural diversity, phylogeographical evolution, interspecific gene flow and kiwifruit cultivar improvement. Acta Hortic. 2007, 753, 31–40. [Google Scholar] [CrossRef]
- Guo, Y.L.; Fitz, J.; Schneeberger, K.; Ossowski, S.; Cao, J.; Weigel, D. Genome-wide comparison of nucleotide-binding site-leucine-rich repeat-encoding genes in Arabidopsis. Plant Physiol. 2011, 157, 757–769. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Tehrim, S.; Zhang, F.; Tong, C.; Huang, J.; Cheng, X.; Dong, C.; Zhou, Y.; Qin, R.; Hua, W.; et al. Genome-wide comparative analysis of NBS encoding genes between Brassica species and Arabidopsis thaliana. BMC Genom. 2014, 15, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michelmore, R.; Meyers, B. Clusters of resistance genes in plants evolve by divergent selection and a birth-and-death process. Genome Res. 1998, 8, 1113–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Die, J.V.; Román, B.; Qi, X.; Rowland, L.J. Genome-scale examination of NBS-encoding genes in blueberry. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF family transcription factors in plant abiotic stress responses. BBA Gene Regul. Mech. 2012, 1819, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Gutterson, N.; Reuber, T.L. Regulation of disease resistance pathways by AP2/ERF transcription factors. Curr. Opin. Plant Biol. 2004, 7, 465–471. [Google Scholar] [CrossRef]
- Guo, Z.J.; Chen, X.J.; Wu, X.L. Overexpression of the AP2/EREBP transcription factor OPBP1 enhances disease resistance and salt tolerance in tobacco. Plant Mol. Biol. 2004, 55, 607–618. [Google Scholar] [CrossRef]
- Sun, F.; Liu, P.; Xu, J. Mutation in RAP2.6L, a transactivator of the ERF transcription factor family, enhances Arabidopsis resistance to Pseudomonas syringae. Physiol. Mol. Plant Pathol. 2010, 74, 295–302. [Google Scholar] [CrossRef]
- Mengnan, W.; Yanxun, Z.; Rui, H. Expression of Vitis amurensis VaERF20 in Arabidopsis thaliana improves resistance to Botrytis cinerea and Pseudomonas syringae pv. Tomato DC3000. Int. J. Mol. Sci. 2018, 19, 696. [Google Scholar]
- Wurms, K.V.; Hardaker, A.J.; Annette, A.C.; Judith, B.; Janet, P.; Joseph, T. Phytohormone and putative defense gene expression differentiates the response of ‘hayward’ kiwifruit to psa and pfm infections. Front. Plant Sci. 2017, 8, 1366. [Google Scholar] [CrossRef] [Green Version]
- Bittner-Eddy, P.D.; Crute, I.R.; Holub, E.B. RPP13 is a simple locus in Arabidopsis thaliana for alleles that specify downy mildew resistance to different avirulence determinants in Peronospora parasitica. Plant J. Cell Mol. Biol. 2000, 21, 177–188. [Google Scholar] [CrossRef]
- Leonelli, L.; Pelton, J.; Schoeffler, A.; Dahlbeck, D.; Berger, J.; Wemmer, D.E.; Staskawicz, B. Structural elucidation and functional characterization of the Hyaloperonospora arabidopsidis effector protein ATR13. PLoS Pathog. 2011, 7, e1002428. [Google Scholar] [CrossRef]
- Tao, W.; Gang, W.; Zhan-Hui, J.; Wang, G.; Jia, Z.-H.; Pan, D.-L. Transcriptome analysis of kiwifruit in response to Pseudomonas syringae pv. actinidiae infection. Int. J. Mol. Sci. 2018, 19, 373. [Google Scholar]
- Hung, J.H.; Weng, Z. Sequence alignment and homology search with BLAST and ClustalW. Cold Spring Harb. Protoc. 2016, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.; Wilm, A.; Lopez, R.; et al. Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Chou, K.C.; Shen, H.B. Plant-mPLoc: A top-down strategy to augment the power for predicting plant protein subcellular localization. PLoS ONE 2010, 5, e11335. [Google Scholar] [CrossRef] [Green Version]
- Cai, B.H.; Zhang, J.Y.; Gao, Z.H.; Qu, S.C.; Tong, Z.G.; Mi, L.; Qiao, Y.S.; Zhang, Z. An improved method for isolation of total RNA from the leaves of Fragaria spp. Jiangsu J. Agric. Sci. 2008, 24, 875–877. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C (T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Type | Number of Genes |
|---|---|---|
| 1 | CC-NBS | 3 |
| 2 | CC-NBS-LRR | 20 |
| 3 | TIR-NBS | 1 |
| 4 | TIR-NBS-LRR | 1 |
| 5 | NBS | 20 |
| 6 | NBS-LRR | 55 |
| Total | Total | 100 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Jia, Z.-H.; Zhang, J.-Y.; Liu, M.; Guo, Z.-R.; Wang, G. Identification and Analysis of NBS-LRR Genes in Actinidia chinensis Genome. Plants 2020, 9, 1350. https://doi.org/10.3390/plants9101350
Wang T, Jia Z-H, Zhang J-Y, Liu M, Guo Z-R, Wang G. Identification and Analysis of NBS-LRR Genes in Actinidia chinensis Genome. Plants. 2020; 9(10):1350. https://doi.org/10.3390/plants9101350
Chicago/Turabian StyleWang, Tao, Zhan-Hui Jia, Ji-Yu Zhang, Min Liu, Zhong-Ren Guo, and Gang Wang. 2020. "Identification and Analysis of NBS-LRR Genes in Actinidia chinensis Genome" Plants 9, no. 10: 1350. https://doi.org/10.3390/plants9101350
APA StyleWang, T., Jia, Z. -H., Zhang, J. -Y., Liu, M., Guo, Z. -R., & Wang, G. (2020). Identification and Analysis of NBS-LRR Genes in Actinidia chinensis Genome. Plants, 9(10), 1350. https://doi.org/10.3390/plants9101350