Long-Term Effect of Cover Crops on Species Abundance and Diversity of Weed Flora

 ,
,  ,
, 
 ,
, 

 and
and 
Abstract
:1. Introduction
2. Results
2.1. Effect of Cover Cropping on Weed Diversity
2.2. Effect of Cover Cropping on Weed Abundance
2.3. Aboveground Weed Species Composition
3. Discussion
4. Materials and Methods
4.1. Experimental Site and Set-Up
4.2. Monitoring, Sampling and Aboveground Biomass Determination
4.3. Weed Flora Analysis
4.4. Meteorological Trend
4.5. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Scavo, A.; Restuccia, A.; Lombardo, S.; Fontanazza, S.; Abbate, C.; Pandino, G.; Anastasi, U.; Onofri, A.; Mauromicale, G. Improving soil health, weed management and nitrogen dynamics by Trifolium subterraneum cover cropping. Agron. Sustain. Dev. 2020, 40, 18. [Google Scholar] [CrossRef]
- Restuccia, A.; Lombardo, S.; Mauromicale, G. Impact of a cultivation system upon the weed seedbank size and composition in a Mediterranean environment. Agriculture 2019, 9, 192. [Google Scholar] [CrossRef] [Green Version]
- Scavo, A.; Mauromicale, G. Integrated weed management in herbaceous field crops. Agronomy 2020, 10, 466. [Google Scholar] [CrossRef] [Green Version]
- Scavo, A.; Restuccia, A.; Abbate, C.; Mauromicale, G. Seeming field allelopathic activity of Cynara cardunculus L. reduces the soil weed seed bank. Agron. Sustain. Dev. 2019, 39, 41. [Google Scholar] [CrossRef]
- Scavo, A.; Pandino, G.; Restuccia, A.; Lombardo, S.; Pesce, G.R.; Mauromicale, G. Allelopathic potential of leaf aqueous extracts from Cynara cardunculus L. on the seedling growth of two cosmopolitan weed species. Ital. J. Agron. 2019, 14, 78–83. [Google Scholar] [CrossRef] [Green Version]
- Landis, D.A. Designing agricultural landscapes for biodiversity-based ecosystem services. Basic Appl. Ecol. 2017, 18, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Nkoa, R.; Owen, M.D.K.; Swanton, C.J. Weed abundance, distribution, diversity, and community analyses. Weed Sci. 2015, 63, 64–90. [Google Scholar] [CrossRef] [Green Version]
- Santín-Montanyá, M.I.; Martín-Lammerding, D.; Walter, I.; Zambrana, E.; Tenorio, J.L. Effects of tillage, crop systems and fertilization on weed abundance and diversity in 4-year dry land winter wheat. Eur. J. Agron. 2013, 48, 43–49. [Google Scholar] [CrossRef]
- Fried, G.; Norton, L.R.; Reboud, X. Environmental and management factors determining weed species composition and diversity in France. Agric. Ecosyst. Environ. 2008, 128, 68–76. [Google Scholar] [CrossRef]
- Gresta, F.; Cristaudo, A.; Onofri, A.; Restuccia, A.; Avola, G. Germination response of four pasture species to temperature, light, and post-harvest period. Plant Biosyst. 2010, 144, 849–856. [Google Scholar] [CrossRef]
- Bàrberi, P. Weed management in organic agriculture: Are we addressing the right issues? Weed Res. 2002, 42, 177–193. [Google Scholar] [CrossRef]
- Jordan, N.; Vatovec, C. Agroecological benefits from weeds. In Weed Biology and Management; Inderjit, Ed.; Springer: Dordrecht, The Netherlands, 2004; pp. 137–158. [Google Scholar] [CrossRef]
- Lemessa, F.; Wakjira, M. Mechanisms of ecological weed management by cover cropping: A review. J. Biol. Sci. 2014, 14, 452–459. [Google Scholar] [CrossRef] [Green Version]
- Gerhards, R.; Schappert, A. Advancing cover cropping in temperate integrated weed management. Pest Manag. Sci. 2020, 76, 42–46. [Google Scholar] [CrossRef]
- Mauromicale, G.; Occhipinti, A.; Mauro, R. Selection of shade-adapted subterranean clover species for cover cropping in orchards. Agron. Sustain. Dev. 2010, 30, 473–480. [Google Scholar] [CrossRef] [Green Version]
- Mauro, R.P.; Occhipinti, A.; Longo, A.M.G.; Mauromicale, G. Effects of shading on chlorophyll content, chlorophyll fluorescence and photosynthesis of subterranean clover. J. Agron. Crop Sci. 2011, 197, 57–66. [Google Scholar] [CrossRef]
- Zohary, M.; Heller, D. The Genus Trifolium; Israel Academy of Sciences and Humanities: Jerusalem, Israel, 1984. [Google Scholar]
- Katznelson, J.; Morley, F.H.W. A taxonomic revision of sect. Calycomorphum of the genus Trifolium. I. The geocarpic species. Isr. J. Bot. 1965, 14, 112–134. [Google Scholar]
- Moonen, A.C.; Bàrberi, P. Size and composition of the weed seedbank after 7 years of different cover crop- maize management systems. Weed Res. 2004, 44, 163–177. [Google Scholar] [CrossRef]
- Bàrberi, P.; Mazzoncini, M. Changes in weed community composition as influenced by cover crop and management system in continuous corn. Weed Sci. 2001, 49, 491–499. [Google Scholar] [CrossRef]
- Den Hollander, N.G.; Bastiaans, L.; Kropff, M.J. Clover as a cover crop for weed suppression in an intercropping design: II. Competitive ability of several clover species. Eur. J. Agron. 2007, 26, 104–112. [Google Scholar] [CrossRef]
- Liu, Q.; Xu, R.; Yan, Z.Q.; Jin, H.; Cui, H.Y.; Lu, L.Q.; Zhang, D.H.; Qin, B. Phytotoxic allelochemicals from roots and root exudates of Trifolium pratense. J. Agric. Food Chem. 2013, 61, 6321–6327. [Google Scholar] [CrossRef]
- Scavo, A.; Abbate, C.; Mauromicale, G. Plant allelochemicals: Agronomic, nutritional and ecological relevance in the soil system. Plant Soil 2019, 442, 23–48. [Google Scholar] [CrossRef]
- Ngouajio, M.; McGiffen, M.E., Jr.; Hutchinson, C.M. Effect of cover crop and management system on weed populations in lettuce. Crop Prot. 2003, 22, 57–64. [Google Scholar] [CrossRef]
- Mirsky, S.B.; Ryan, M.R.; Teasdale, J.R.; Curran, W.S.; Reberg-Horton, C.S.; Spargo, J.T.; Wells, M.S.; Keene, C.L.; Moyer, J.W. Overcoming weed management challenges in cover crop–based organic rotational no-till soybean production in the eastern United States. Weed Technol. 2013, 27, 193–203. [Google Scholar] [CrossRef]
- Concenço, G.; de Farias, P.M.; Quintero, N.F.A.; Schreiber, F.; Galon, L.; Tomazi, M.; Moisinho, I.S.; Coradini, M.C.; Ceolin, W.C.; Andres, A. Phytosociological surveys in weed science: Old concept, new approach. In Plant Ecology-Traditional Approaches to Recent Trends; Yousaf, Z., Ed.; IntechOpen: London, UK, 2017; pp. 121–146. [Google Scholar] [CrossRef] [Green Version]
- Swanton, C.J.; Shrestha, A.; Roy, R.C.; Ball-Coelho, B.R.; Knezevic, S.Z. Effect of tillage systems, N, and cover crop on the composition of weed flora. Weed Sci. 1999, 47, 454–461. [Google Scholar] [CrossRef]
- Shrestha, A.; Knezevic, S.Z.; Roy, R.C.; Ball-Coelho, B.R.; Swanton, C.J. Effect of tillage, cover crop and crop rotation on the composition of weed flora in a sandy soil. Weed Res. 2002, 42, 76–87. [Google Scholar] [CrossRef]
- Soil Survey Staff. Soil Taxonomy: A Basic System of Soil Classification for Making and Interpreting Soil Surveys, 2nd ed.; U.S. Gov. Print. Office: Washington, DC, USA, 1999.
- Conti, F.; Abbate, G.; Alessandrini, A.; Blasi, C. An Annotated Checklist of the Italian Vascular Flora; Palombi Editore: Rome, Italy, 2005. [Google Scholar]
- Travlos, I.S.; Cheimona, N.; Roussis, I.; Bilalis, D.J. Weed-species abundance and diversity indices in relation to tillage systems and fertilization. Front. Environ. Sci. 2018, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Real, R.; Vargas, J.M. The probabilistic basis of Jaccard’s index of similarity. Syst. Biol. 1996, 45, 380–385. Available online: https://www.jstor.org/stable/2413572 (accessed on 15 August 2020). [CrossRef]
- Deligios, P.A.; Carboni, G.; Farci, R.; Solinas, S.; Ledda, L. The influence of herbicide underdosage on the composition and diversity of weeds in oilseed rape (Brassica napus L. var. oleifera DC) Mediterranean fields. Sustainability 2019, 11, 1653. [Google Scholar] [CrossRef] [Green Version]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 2012; pp. 429–463. [Google Scholar]






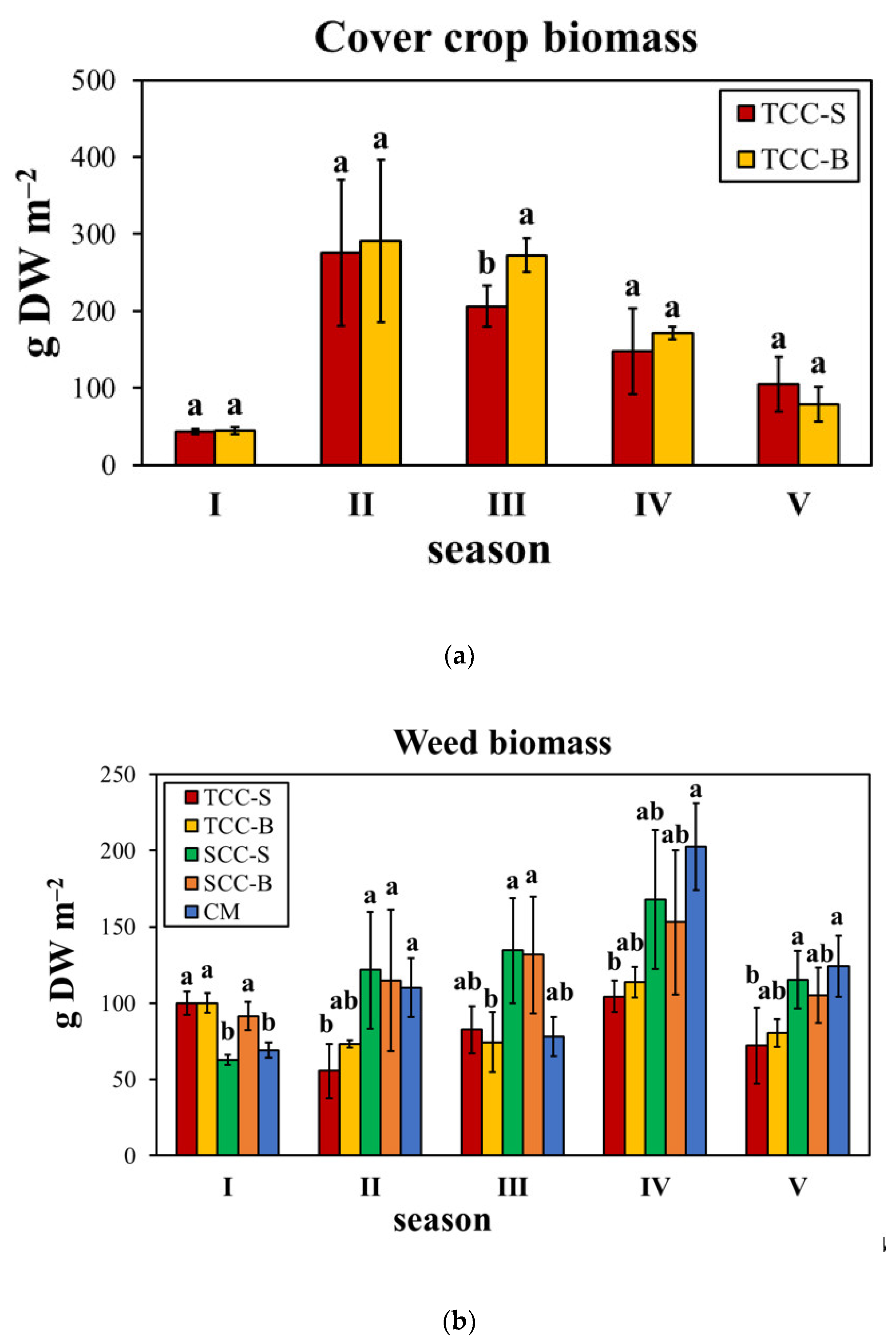{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weed Species | Botanical Family | Life Cycle | EG | BG | F (%) 1 | RF (%) 1 |
|---|---|---|---|---|---|---|
| Avena sp. | Poaceae | Annual | Sp | T | 5.0 | 2.34 |
| Adonis annua L. subsp. cupaniana (Guss.) Steinberg | Ranunculaceae | Annual | Sp | T | 8.0 | 2.13 |
| Anagallis arvensis L. | Primulaceae | Annual | Au-Wi | T | 18.0 | 5.84 |
| Beta vulgaris L. | Amaranthaceae | Perennial | Su | H | 11.0 | 3.43 |
| Brassica rapa L. subsp. campestris (L.) A.R. Clapham | Brassicaceae | Perennial | Sp-Su | H | 1.2 | 0.17 |
| Capsella bursa-pastoris (L.) Medik. | Brassicaceae | Biennial | Ind | H | 1.0 | 0.18 |
| Chenopodium opulifolium Schrad. ex W.D.J. Koch & Ziz | Chenopodiaceae | Annual | Su | T | 1.0 | 0.44 |
| Chenopodium sp. | Chenopodiaceae | Annual | Su | T | 5.0 | 1.09 |
| Cichorium intybus L. | Asteraceae | Perennial | Ind | H | 14.0 | 3.23 |
| Cirsium arvense (L.) Scop. | Asteraceae | Perennial | Su | G | 2.0 | 1.0 |
| Convolvulus arvensis L. | Convolvulaceae | Perennial | Ind | G | 5.0 | 2.44 |
| Conyza canadensis L. | Asteraceae | Annual | Sp-Su | T | 3.0 | 1.20 |
| Daucus carota L. | Apiaceae | Biennial | Sp-Su-Au | H | 2.2 | 0.54 |
| Diplotaxis erucoides (L.) DC. | Brassicaceae | Annual | Ind | T | 8.0 | 2.54 |
| Dittrichia viscosa (L.) Greuter subsp. viscosa | Asteraceae | Perennial | Au | H | 1.0 | 0.22 |
| Ecballium elaterium (L.) A. Rich. | Cucurbitaceae | Annual | Su | T | 22.0 | 7.86 |
| Erigeron sumatrensis Retz. | Asteraceae | Annual | Su | T | 9.0 | 2.58 |
| Foeniculum vulgare Mill. | Apiaceae | Perennial | Su | H | 1.0 | 0.50 |
| Fumaria officinalis L. | Fumariacee | Annual | Sp-Su-Au | T | 10.2 | 2.52 |
| Galactites elegans (All.) Soldano | Asteraceae | Biennial | Sp-Su | H | 1.0 | 0.24 |
| Galium aparine L. | Rubiaceae | Annual | Sp-Su-Au | T | 15.8 | 3.68 |
| Glebionis coronaria (L.) Spach | Asteraceae | Annual | Sp-Su | T | 2.0 | 0.80 |
| Helminthotheca echioides (L.) Holub | Asteraceae | Annual | Su-Au | T | 48.6 | 13.16 |
| Hypochaeris radicata L. | Asteraceae | Perennial | Sp | H | 6.6 | 1.61 |
| Lamium amplexicaule L. | Lamiaceae | Annual | Ind | T | 2.0 | 0.36 |
| Malva sylvestris L. | Malvaceae | Perennial | Ind | H | 4.2 | 0.99 |
| Medicago polymorpha L. | Fabaceae | Annual | Sp | T | 2.2 | 0.35 |
| Papaver rhoeas L. | Papaveraceae | Annual | Wi | T | 9.2 | 2.39 |
| Reichardia picroides (L.) Roth | Asteraceae | Perennial | Ind | H | 9.8 | 1.77 |
| Setaria verticillata (L.) P. Beauv. | Poaceae | Annual | Su | T | 31.0 | 7.41 |
| Setaria italica subsp. viridis (L.) Thell. | Poaceae | Annual | Su-Au | T | 27.0 | 8.28 |
| Silene sp. | Caryophyllaceae | Perennial | Sp-Su | H | 3.8 | 0.64 |
| Sinapis arvensis L. | Brassicaceae | Annual | Sp | T | 51.0 | 14.79 |
| Sonchus asper (L.) Hill | Asteraceae | Biennial | Ind | H | 61.0 | 16.87 |
| Sonchus oleraceus L. | Asteraceae | Biennial | Ind | H | 11.0 | 2.76 |
| Stellaria media (L.) Vill. | Caryophyllaceae | Biennial | Ind | H | 1.0 | 0.40 |
| Trigonella foenum-graecum L. | Fabaceae | Annual | Sp | T | 11.0 | 3.59 |
| Vicia faba L. | Fabaceae | Annual | Sp | T | 5.0 | 1.25 |
| Treatment | No. | Season | No. |
|---|---|---|---|
| TCC-S | 6.8 b | I | 4.4 b |
| TCC-B | 10.2 a | II | 9.0 a |
| SCC-S | 8.0 a | III | 7.4 ab |
| SCC-B | 8.0 a | IV | 9.6 a |
| CM | 8.6 a | V | 11.2 a |
| F-test | * | F-test | ** |
| SED 1 | 1.98 | SED 1 | 1.37 |
| Treatments | I | II | III | IV | V | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| J | S | J | S | J | S | J | S | J | S | |
| TCC-S × TCC-B | 22.2 | 36.4 | 66.7 | 80.0 | 30.0 | 46.2 | 42.9 | 60.0 | 50.0 | 66.7 |
| TCC-S × SCC-S | 12.5 | 22.2 | 50.0 | 66.7 | 30.0 | 46.2 | 55.6 | 71.4 | 42.9 | 60.0 |
| TCC-S × SCC-B | 14.3 | 25.0 | 23.1 | 37.5 | 25.0 | 40.0 | 36.4 | 53.3 | 53.8 | 70.0 |
| TCC-S × CM | 12.5 | 22.2 | 50.0 | 66.7 | 16.7 | 28.6 | 42.9 | 60.0 | 45.5 | 62.5 |
| TCC-B × SCC-S | 11.1 | 20.0 | 66.7 | 80.0 | 16.7 | 28.6 | 33.3 | 50.0 | 50.0 | 66.7 |
| TCC-B × SCC-B | 28.6 | 44.4 | 38.5 | 55.6 | 23.1 | 37.5 | 23.5 | 38.1 | 92.9 | 96.3 |
| TCC-B × CM | 11.1 | 20.0 | 42.9 | 60.0 | 15.4 | 26.7 | 36.8 | 53.8 | 43.8 | 60.9 |
| SCC-S × SCC-B | 40.0 | 57.1 | 33.3 | 50.0 | 60.0 | 75.0 | 66.7 | 80.0 | 44.4 | 61.5 |
| SCC-S × CM | 14.3 | 25.0 | 50.0 | 66.7 | 66.7 | 80.0 | 42.9 | 60.0 | 37.5 | 54.5 |
| SCC-B × CM | 40.0 | 57.1 | 33.3 | 50.0 | 70.0 | 82.4 | 50.0 | 66.7 | 37.5 | 54.5 |
| Treatments | Aboveground Biomass (g DW m‒2) | |||
|---|---|---|---|---|
| Trifolium Subterraneum | Weeds | Total | ||
| CC | TCC-S | 155.5 (43.1) a | 82.9 (15.2) b | 238.4 (40.7) a |
| TCC-B | 171.7 (32.7) a | 88.3 (9.5) b | 260.0 (36.7) a | |
| SCC-S | 0.0 | 120.4 (28.0) a | 120.4 (28.0) b | |
| SCC-B | 0.0 | 119.2 (31.8) a | 119.2 (31.8) b | |
| CM | 0.0 | 116.8 (17.2) a | 116.8 (17.2) b | |
| S | I | 44.4 (4.1) d | 84.7 (6.4) b | 102.5 (6.5) c |
| II | 283.1 (99.9) a | 95.1 (24.8) b | 208.3 (35.6) a | |
| III | 239.2 (24.5) a | 100.3 (24.1) b | 195.8 (31.8) a | |
| IV | 159.4 (32.1) b | 148.3 (28.3) a | 212.0 (38.2) a | |
| V | 92.0 (29.1) c | 99.4 (18.2) b | 136.2 (19.7) b | |
| ANOVA | ||||
| CC | 0.4 NS | 9.2 *** | 64.1 *** | |
| S | 63.6 *** | 14.2 *** | 31.6 *** | |
| CC × S | 1.3 NS | 4.0 *** | 4.2 *** | |
| Variable | PC1 | PC2 | PC3 | PC4 |
|---|---|---|---|---|
| ANAAR | 0.383 | −0.211 | −0.167 | 0.182 |
| BEVULG | −0.035 | −0.015 | −0.752 | −0.056 |
| CICIN | −0.363 | −0.015 | 0.079 | 0.534 |
| ECBAL | 0.317 | 0.382 | −0.056 | −0.181 |
| GALAP | 0.352 | 0.050 | 0.319 | 0.388 |
| HELEC | 0.124 | −0.549 | −0.042 | 0.211 |
| SETVER | −0.430 | 0.030 | 0.019 | −0.027 |
| SETIT | −0.404 | −0.070 | 0.206 | 0.182 |
| SINAR | −0.290 | 0.426 | −0.094 | −0.088 |
| SONCAS | 0.043 | 0.329 | −0.410 | 0.628 |
| TRIGO | 0.215 | 0.458 | 0.273 | 0.121 |
| Season | Emergence | Flowering | Length of the Biological Cycle 1 |
|---|---|---|---|
| I | 15 December 2015 | 22 May 2016 | ~200 days |
| II | 22 November 2016 | 12 April 2017 | ~220 days |
| III | 10 October 2017 | 29 April 2018 | ~250 days |
| IV | 13 October 2018 | 26 April 2019 | ~240 days |
| V | 5 November 2019 | 4 April 2020 | ~230 days |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Restuccia, A.; Scavo, A.; Lombardo, S.; Pandino, G.; Fontanazza, S.; Anastasi, U.; Abbate, C.; Mauromicale, G. Long-Term Effect of Cover Crops on Species Abundance and Diversity of Weed Flora. Plants 2020, 9, 1506. https://doi.org/10.3390/plants9111506
Restuccia A, Scavo A, Lombardo S, Pandino G, Fontanazza S, Anastasi U, Abbate C, Mauromicale G. Long-Term Effect of Cover Crops on Species Abundance and Diversity of Weed Flora. Plants. 2020; 9(11):1506. https://doi.org/10.3390/plants9111506
Chicago/Turabian StyleRestuccia, Alessia, Aurelio Scavo, Sara Lombardo, Gaetano Pandino, Stefania Fontanazza, Umberto Anastasi, Cristina Abbate, and Giovanni Mauromicale. 2020. "Long-Term Effect of Cover Crops on Species Abundance and Diversity of Weed Flora" Plants 9, no. 11: 1506. https://doi.org/10.3390/plants9111506
APA StyleRestuccia, A., Scavo, A., Lombardo, S., Pandino, G., Fontanazza, S., Anastasi, U., Abbate, C., & Mauromicale, G. (2020). Long-Term Effect of Cover Crops on Species Abundance and Diversity of Weed Flora. Plants, 9(11), 1506. https://doi.org/10.3390/plants9111506