Managing an Invasive Weed Species, Parthenium hysterophorus, with Suppressive Plant Species in Australian Grasslands


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Suppression of Growth and Abundance of Parthenium Weed
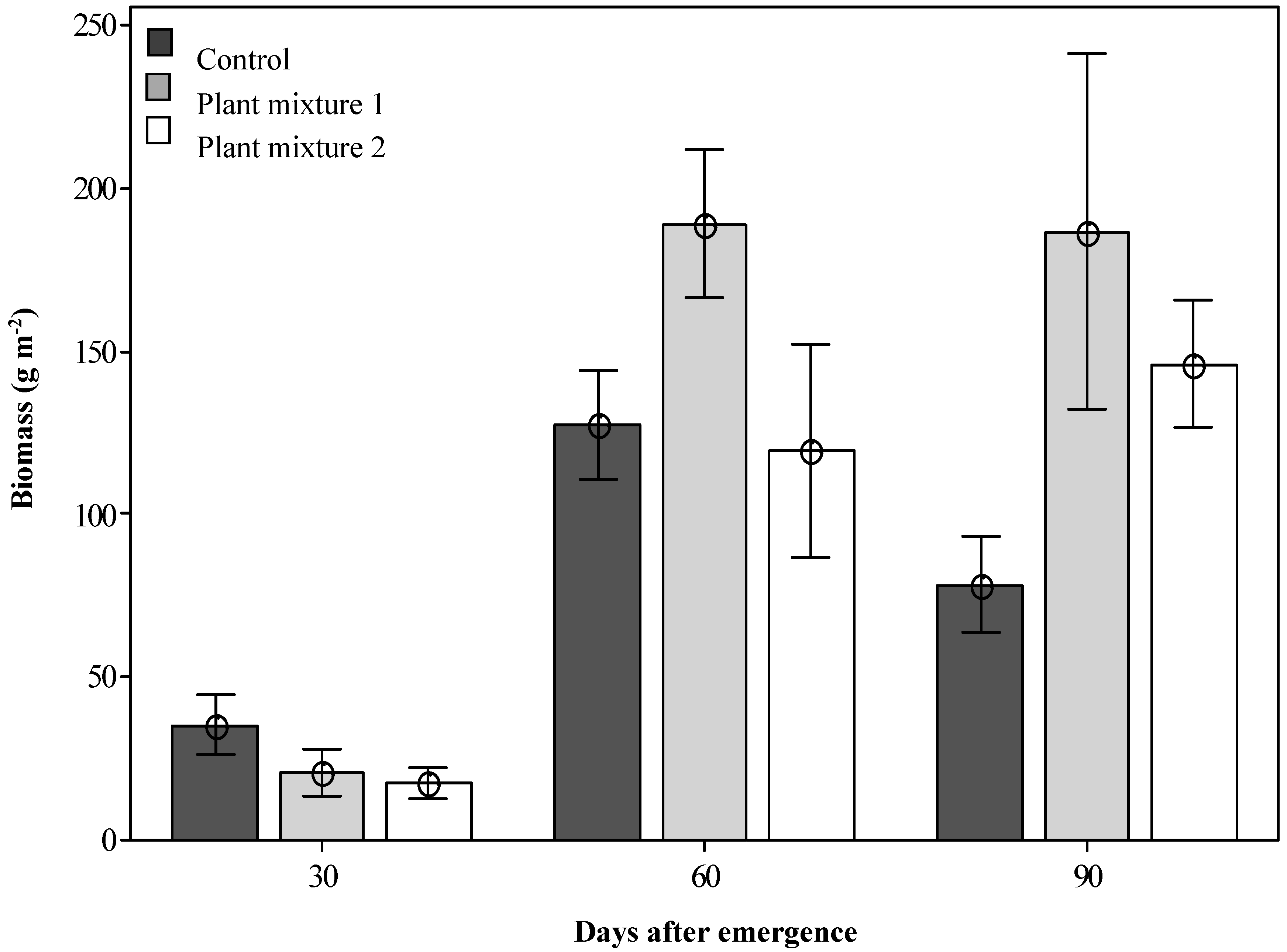
2.2. Biomass Production of the Plant Mixtures
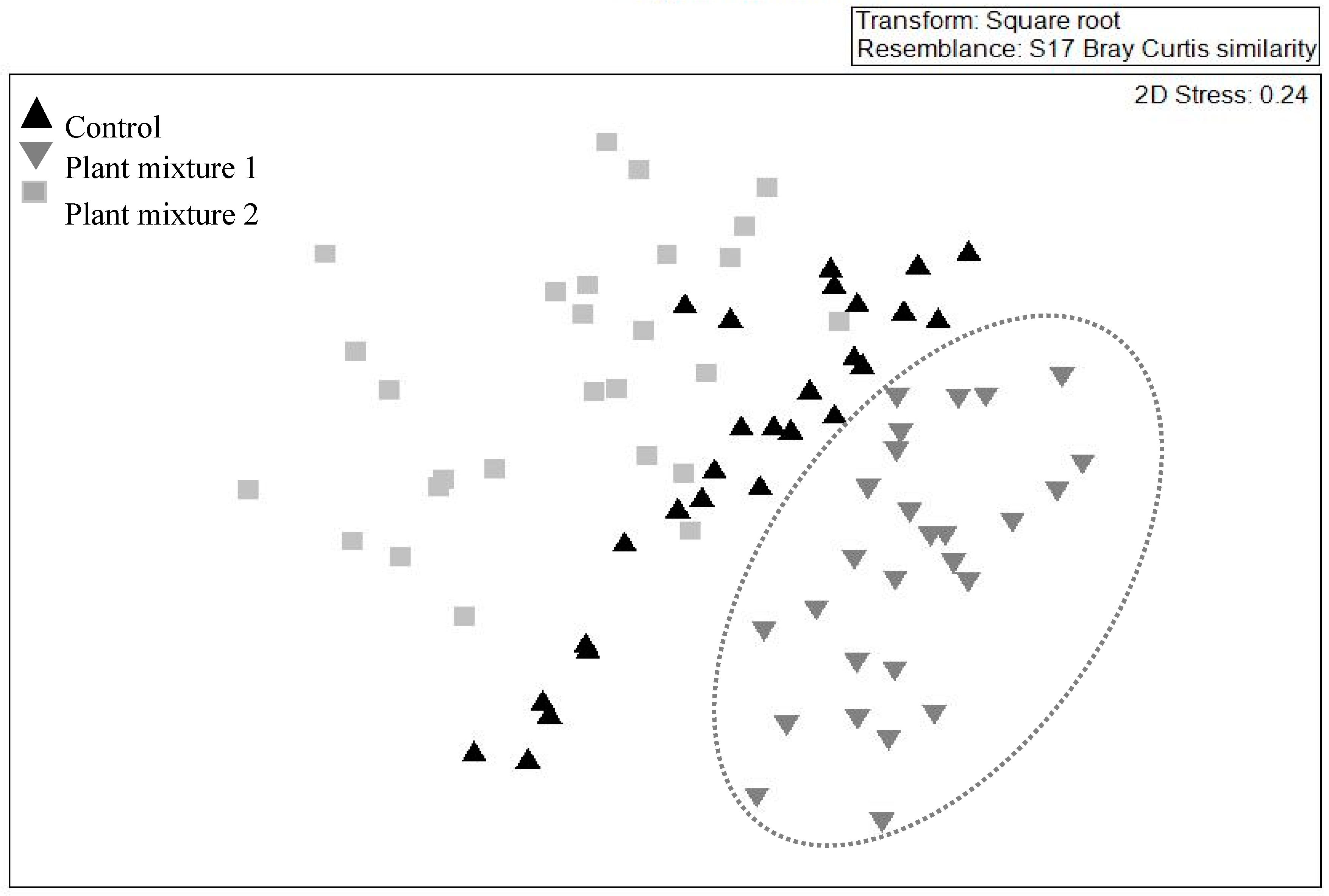
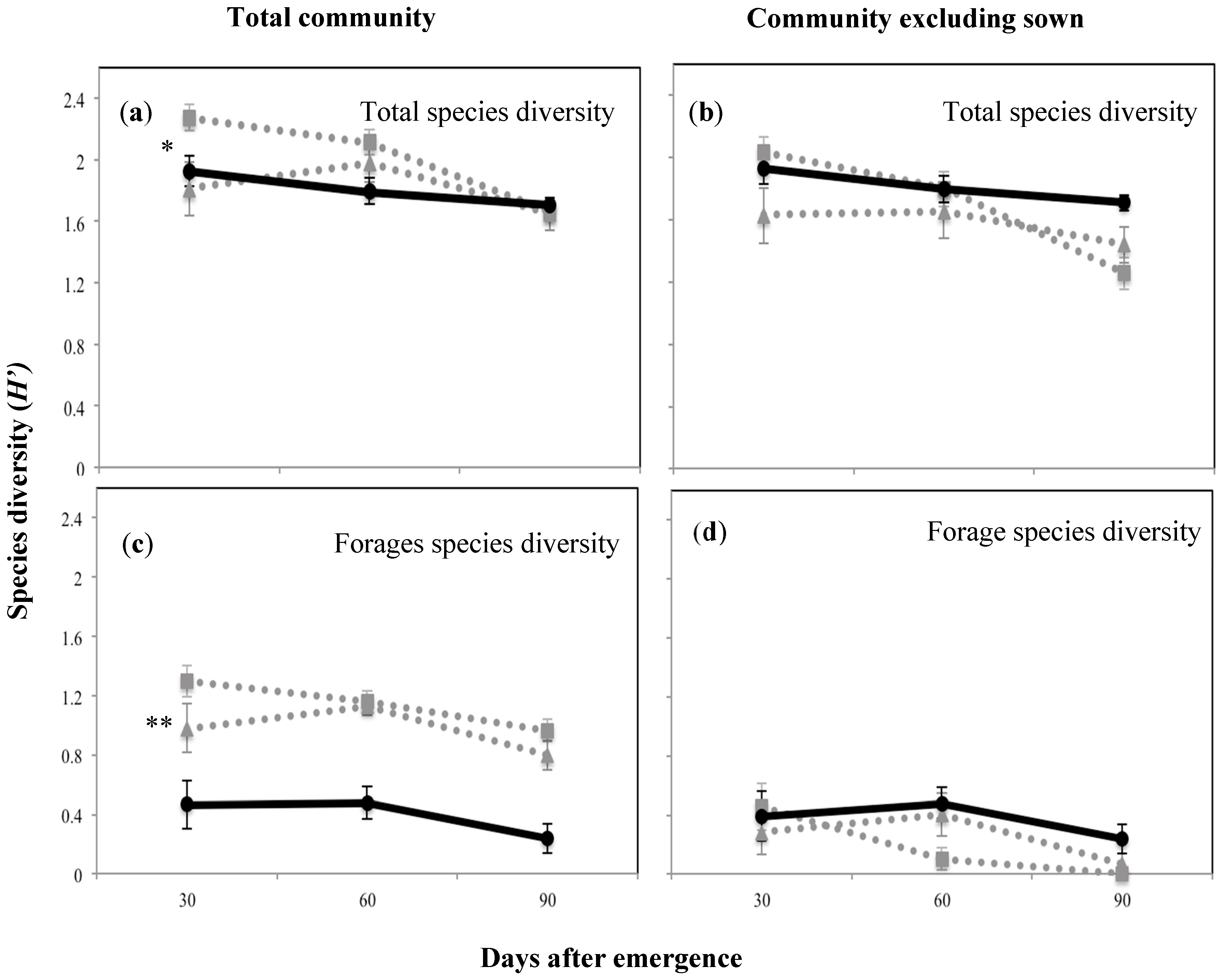
2.3. Below- and Above-Ground Species Composition and Diversity
3. Discussion
4. Materials and Methods
4.1. Study Location
4.2. Experimental Design and Treatments
4.3. Weed Population
4.4. Data Collection
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hejda, M.; Pysek, P.; Jarosik, V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- Ziska, L.H.; McConnell, L.L. Climate change, carbon dioxide, and pest biology: Monitor, mitigate, manage. J. Agric. Food Chem. 2015, 64, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Adkins, S.W.; Shabbir, A. Biology, ecology and management of the invasive parthenium weed (Parthenium hysterophorus L.). Pest. Manag. Sci. 2014, 70, 1023–1029. [Google Scholar] [CrossRef] [PubMed]
- Bajwa, A.A.; Chauhan, B.S.; Farooq, M.; Shabbir, A.; Adkins, S.W. What do we really know about alien plant invasion? a review of the invasion mechanism of one of the world’s worst weeds. Planta 2016, 244, 39–57. [Google Scholar] [CrossRef]
- Bajwa, A.A.; Chauhan, B.S.; Adkins, S.W. Morphological, physiological and biochemical responses of two Australian biotypes of Parthenium hysterophorus to different soil moisture regimes. Environ. Sci. Pollut. Res. 2017, 24, 16186–16194. [Google Scholar] [CrossRef]
- Timsina, B.; Shrestha, B.B.; Rokaya, M.B.; Münzbergová, Z. Impact of Parthenium hysterophorus L. invasion on plant species composition and soil properties of grassland communities in Nepal. Flora 2011, 206, 233–240. [Google Scholar] [CrossRef]
- Navie, S.C.; Panetta, F.D.; McFadyen, R.E.; Adkins, S.W. Germinable soil seedbanks of central Queensland rangelands invaded by the exotic weed Parthenium hysterophorus L. Weed Biol. Manag. 2004, 4, 154–167. [Google Scholar] [CrossRef]
- Nigatu, L.; Hassen, A.; Sharma, J.; Adkins, S.W. Impact of Parthenium hysterophorus on grazing land communities in north-eastern Ethiopia. Weed Biol. Manag. 2010, 10, 143–152. [Google Scholar] [CrossRef]
- Nguyen, T.; Bajwa, A.A.; Belgeri, A.; Navie, S.; O’Donnell, C.; Adkins, S. Impact of an invasive weed, Parthenium hysterophorus, on a pasture community in south east Queensland, Australia. Environ. Sci. Pollut. Res. 2017, 24, 27188–27200. [Google Scholar] [CrossRef]
- Bajwa, A.A.; Ullah, A.; Farooq, M.; Chauhan, B.S.; Adkins, S. Effect of different densities of parthenium weed (Parthenium hysterophorus L.) on the performance of direct-seeded rice under aerobic conditions. Arch. Agron. Soil Sci. 2018, 65, 796–808. [Google Scholar] [CrossRef]
- Bajwa, A.A.; Nguyen, T.; Navie, S.; O’Donnell, C.; Adkins, S. Weed seed spread and its prevention: The role of roadside wash down. J. Environ. Manag. 2018, 208, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Bajwa, A.A.; Ullah, A.; Farooq, M.; Chauhan, B.S.; Adkins, S. Chemical control of parthenium weed (Parthenium hysterophorus L.) in two contrasting cultivars of rice under direct-seeded conditions. Crop Prot. 2019, 117, 26–36. [Google Scholar] [CrossRef]
- Bajwa, A.A.; Chauhan, B.S.; Adkins, S.W. Germination ecology of two Australian biotypes of ragweed parthenium (Parthenium hysterophorus) relates to their invasiveness. Weed Sci. 2018, 66, 62–70. [Google Scholar] [CrossRef]
- Nguyen, T.L.T.; Bajwa, A.A.; Navie, S.C.; O’Donnell, C.; Adkins, S.W. The soil seedbank of pasture communities in central Queensland invaded by Parthenium hysterophorus L. Range Ecol. Manag. 2017, 70, 244–254. [Google Scholar] [CrossRef]
- Adamson, D.C. Introducing Dynamic Considerations When Economically Evaluating Weeds. Master’s Thesis, The University of Queensland, Brisbane, Australia, 1996. [Google Scholar]
- Belgeri, A.; Navie, S.C.; Vivian-Smith, G.; Adkins, S.W. Early recovery signs of an Australian grassland following the management of Parthenium hysterophorus L. Flora 2014, 209, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Shabbir, A.; Dhileepan, K.; Zalucki, M.P.; Khan, N.; Adkins, S.W. Reducing the fitness of an invasive weed, Parthenium hysterophorus: Complementing biological control with plant competition. J. Environ. Manag. 2020, 254, 109790. [Google Scholar] [CrossRef] [PubMed]
- Shabbir, A.; Dhileepan, K.; O’Donnell, C.; Adkins, S.W. Complementing biological control with plant suppression: Implications for improved management of parthenium weed (Parthenium hysterophorus L.). Biol. Control 2013, 64, 270–275. [Google Scholar] [CrossRef]
- McFadyen, R.C. Biological control against parthenium weed in Australia. Crop. Prot. 1992, 11, 400–407. [Google Scholar] [CrossRef]
- Dhileepan, K. Effectiveness of introduced biocontrol insects on the weed Parthenium hysterophorus (Asteraceae) in Australia. Bull. Entomol. Res. 2001, 91, 167–176. [Google Scholar]
- Dhileepan, K.; Setter, S.D.; McFadyen, R.E. Impact of defoliation by the biocontrol agent Zygogramma bicolorata on the weed Parthenium hysterophorus in Australia. BioControl 2000, 45, 501–512. [Google Scholar] [CrossRef]
- Bajwa, A.A.; Walsh, M.; Chauhan, B.S. Weed management using crop competition in Australia. Crop Prot. 2017, 95, 8–13. [Google Scholar] [CrossRef]
- Khan, N.; O’Donnell, C.; George, D.; Adkins, S.W. Suppressive ability of selected fodder plants on the growth of Parthenium hysterophorus. Weed Res. 2013, 53, 61–68. [Google Scholar] [CrossRef]
- O’Donnell, C.; Adkins, W. Management of parthenium weed through competitive displacement with beneficial plants. Weed Biol. Manag. 2005, 5, 77–79. [Google Scholar] [CrossRef]
- Khan, N.; Shabbir, A.; George, D.; Hassan, G.; Adkins, S.W. Suppressive fodder plants as part of an integrated management program for Parthenium hysterophorus L. Field Crops Res. 2014, 156, 172–179. [Google Scholar] [CrossRef]
- Sanderson, M.A.; Skinner, R.H.; Barker, D.J.; Edwards, G.R.; Tracy, B.F.; Wedin, D.A. Plant species diversity and management of temperate forage and grazing land ecosystems. Crop Sci. 2004, 44, 1132–1144. [Google Scholar] [CrossRef]
- Sanderson, M.A. Stability of production and plant species diversity in managed grasslands: A retrospective study. Basic Appl. Ecol. 2010, 11, 216–224. [Google Scholar] [CrossRef]
- Butler, D.W.; Russell, J.F. Buffel Grass and Fire in a Gidgee and Brigalow Woodland: A Case of Study from Central Queensland (Ecological Management and Restoration); The Queensland Herbarium: Mount Coot-tha, Australia, 2003.
- Tracy, B.F.; Sanderson, M.A. Forage productivity, species evenness and weed invasion in pasture communities. Agric. Ecosyst. Environ. 2004, 102, 175–183. [Google Scholar] [CrossRef]
- Tozer, K.N.; Barker, G.M.; Cameron, C.A.; James, T.K. Relationship between seedbank and above-ground botanical composition during spring. N. Z. Plant. Prot. 2010, 63, 90–95. [Google Scholar]
- Shabbir, A.; Bajwa, A.A.; Dhileepan, K.; Zalucki, M.; Khan, N.; Adkins, S. Integrated use of biological approaches provides effective control of parthenium weed. Arch. Agron. Soil Sci. 2018, 64, 1861–1878. [Google Scholar] [CrossRef]
- Shabbir, A.; Ali, S.; Khan, A.I.; Belgeri, A.; Khan, N.; Adkins, S. Suppressing parthenium weed with beneficial plants in Australian grasslands. Int. J. Pest. Manag. 2019. [Google Scholar] [CrossRef]
- Stanley, T.D.; Ross, E.M. Flora of South-Eastern Queensland; Queensland Department of Primary Industries: Brisbane, Australia, 1989.
- PlantNET. National Herbarium of New South Wales. 2011. Available online: http://plantnet.rbgsyd.nsw.gov.au (accessed on 19 July 2014).
- Khan, N. Long Term, Sustainable Management of Parthenium Weed (Parthenium hysterophorus L.) Using Suppressive Plants. Ph.D. Thesis, The University of Queensland, St. Lucia, QLD, Australia, 2011. [Google Scholar]
- Belgeri, A.; Navie, S.; Adkins, S.W. Screening parthenium weed (Parthenium hysterophorus L.) seedlings for their allelopathic potential. Pak. J. Weed Sci. Res. 2012, 18, 727–731. [Google Scholar]
- Belgeri, A.; Adkins, S.W. Allelopathic potential of invasive parthenium weed (Parthenium hysterophorus L.) seedlings on grassland species in Australia. Allelopath. J. 2015, 36, 1–14. [Google Scholar]
- Moles, A.T.; Westoby, M. Seed size and plant strategy across the whole life cycle. Oikos 2006, 113, 91–105. [Google Scholar] [CrossRef]
- Low, T. Tropical pasture plants as weeds. Trop. Grassl. 1997, 31, 337–343. [Google Scholar]
- Maranon, T.R.; Zamora, R.; Villar, M.A.; Zavala, J.L.; Quero, I.; Pérez-Ramos, I.; Mendoza, C.J. Regeneration of tree species and restoration under contrasted Mediterranean habitats: Field and glasshouse experiments. J. Ecol. Environ. Sci. 2004, 30, 187–196. [Google Scholar]
- Altesor, A.; Oesterheld, M.; Leoni, E.; Lezama, F.; Rodriguez, C. Effect of grazing on community structure and productivity of a Uruguayan grassland. Plant. Ecol. 2005, 179, 83–91. [Google Scholar] [CrossRef]
- Bureau of Meteorology (BOM). 2012. Available online: http://www.bom.gov.au/ (accessed on 19 September 2012).
- Sanyal, D.; Bhowmik, P.C.; Anderson, R.L.; Shrestha, A. Revisiting the perspective and progress of integrated weed management. Weed Sci. 2008, 56, 161–167. [Google Scholar] [CrossRef]
- Krebs, C.J. Ecology: The Experimental Analysis of Distribution and Abundance, 3rd ed.; Harper & Row: Sydney, Australia, 1985. [Google Scholar]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]


 ) are enclosed as only this mixture did not overlap with the control.
) are enclosed as only this mixture did not overlap with the control.
) are enclosed as only this mixture did not overlap with the control.
) are enclosed as only this mixture did not overlap with the control.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pasture Mixture | Plant Species | Plant Establishment at the Three Survey Occasions (DAE; Plants m−2) | Height at the Simulated Grazing Times (cm) | ||||
|---|---|---|---|---|---|---|---|
| 30 | 60 | 90 | 30 | 60 | 90 | ||
| One | Rhodes grass | 5.6 ± 0.8 | 5.9 ± 0.8 | 6.6 ± 0.3 | 50.6 ± 6.8 | 98.7 ± 4.5 | 131.7 ± 8.1 |
| Bisset bluegrass | 4.0 ± 1.2 | 1.0 ± 0.0 | 1.0 ± 0.0 | 27.6 ± 8.6 | 63.0 ± 1.0 | 88.5 ± 3.3 | |
| Butterfly pea | 2.8 ± 0.4 | 4.8 ± 0.6 | 1.8 ± 0.3 | 15.2 ± 1.3 | 45.3 ± 4.8 | 67.8 ± 4.9 | |
| Green panic | 3.3 ± 0.9 | 3.1± 0.5 | 4.9 ± 1.1 | 43.5 ± 9.2 | 114.9 ± 3.4 | 143.7 ± 8.0 | |
| Total | 15.7 ± 0.9 | 14.8 ± 1.1 | 14.3 ± 1.2 | - | - | - | |
| Two | Forest bluegrass | 4.4 ± 1.3 | 1.0 ± 0.0 | 2.0 ± 0.0 | 18.7 ± 1.7 | 53.5 ± 0.5 | 112.5 ± 3.5 |
| Queensland bluegrass | 5.8 ± 0.9 | 1.3 ± 0.1 | 1.0 ± 0.0 | 28.3 ± 1.9 | 60.2 ± 4.0 | 102.3 ± 23.0 | |
| Siratro | 2.1 ± 0.4 | 4.1 ± 0.4 | 3.4 ± 0.6 | 14.5 ± 0.3 | 54.8 ± 2.4 | 65.6 ± 7.9 | |
| Buffel grass | 4.3 ± 0.7 | 4.9 ± 0.7 | 6.3 ± 0.8 | 25.18 ± 1.9 | 103.0 ± 3.0 | 147.9 ± 67.3 | |
| Total | 16.6 ± 1.0 | 11.1 ± 0.9 | 12.7 ± 1.0 | - | - | - | |
| Family | Species | Status 1 | Germinable Seeds (m−2) |
|---|---|---|---|
| Amaranthaceae | Alternanthera nana R.Br | W | 141 |
| * Amaranthus spinosus L. | W | 234 | |
| Apiaceae | * Cyclospermum leptophyllum (Pers.) Sprague | W | 63 |
| * Soliva sp. | W | 16 | |
| Asteraceae | * Gamochaeta pensylvanica (Willd.) Cabrera | W | 63 |
| * Cirsium vulgare (Savi) Ten. | W | 31 | |
| * Conyza bonariensis (L.) Cronq. | W | 63 | |
| * Conyza sumatrensis (Retz.) E.H. Walker | W | 16 | |
| * Crassocephalum crepidioides (Benth.) S Moore | W | 16 | |
| * Gamochaeta americana (Mill.) Wedd | W | 16 | |
| * Parthenium hysterophorus L. | W | 1969 | |
| Brassicaceae | * Lepidium africanum (Burm.f.) DC. | W | 1000 |
| * Lepidium bonariense L. | W | 63 | |
| * Lepidium didymum L. | W | 625 | |
| Campanulaceae | Wahlenbergia stricta (R. Br.) Sweet | W | 63 |
| Chennopodiaceae | Dysphania carinata (R.Br.) Mosyakin & Clemants | W | 78 |
| Dysphania pumilio (R.Br.) Mosyakin & Clemants | W | 31 | |
| Einadia trigonos (Schult.) Paul G. Wilson | W | 47 | |
| Crassulaceae | Crassula sieberiana (Schult. & Schult.f.) Druce | W | 47 |
| Cyperaceae | * Cyperus brevifolius (Rottb.) Hassk. | W | 531 |
| Cyperus gracilis R.Br. | W | 6172 | |
| Cyperus iria L. | W | 16 | |
| Fabaceae | * Medicago polymorpha L. | D | 23 |
| Gentianaceae | * Schenkia spicata (L.) Mansion | W | 141 |
| Iridaceae | * Sisyrinchium sp. Peregian | W | 47 |
| Malvaceae | * Sida cordifolia L. | W | 31 |
| * Sida rhombifolia L. | W | 31 | |
| Oxalidaceae | Oxalis exilis A. Cunn. | W | 1031 |
| Oxalis purpurea L. | W | 0 | |
| Plantaginaceae | Plantago debilis R.Br. | W | 47 |
| Poaceae | Chloris divaricata R.Br. | D | 94 |
| * Chloris gayana Kunth | D | 94 | |
| * Cynodon dactylon (L.) Pers | D | 78 | |
| Digitaria didactyla Willd. | D | 1000 | |
| * Eleusine indica (L.) Gaertn. | W | 141 | |
| Paspalidium distans (Trin.) Hughes | D | 234 | |
| Sporobolus creber De Nardi | D | 47 | |
| Sporobolus elongatus R.Br. | D | 375 | |
| * Urochloa panicoides Beauv. | W | 16 | |
| Polygonaceae | Rumex brownii Campd. | W | 16 |
| Portulacaceae | * Portulaca oleracea L. | W | 250 |
| Pasture Mixture | Plant Species | Life Form 2 | Status 3 | Contribution (%) | |
|---|---|---|---|---|---|
| Common Name | Scientific Name | ||||
| Control (similarity 59.5%) | Liverseed grass | Urochloa panicoides Beauv. | G | W | 16.9 |
| Paddy’s lucerne | Sida rhombifolia L. | Sb | W | 15.0 | |
| Parthenium weed | Parthenium hysterophorus L. | H | W | 12.7 | |
| Bala | Sida cordifolia L. | Sb | W | 12.3 | |
| Goose grass | Eleusine indica (L.) Gaertn. | G | W | 11.4 | |
| Spiny amaranthus | Amaranthus spinosus L. | H | W | 11.3 | |
| Green crumbweed | Dysphania carinata (R.Br.) Mosyakin & Clemants * | H | W | 6.3 | |
| Blue couch | Digitaria didactyla Willd. * | G | D | 3.5 | |
| Spiked malvastrum | Malvastrum coromandelianum (L.) Garcke | Sb | W | 2.7 | |
| One (similarity 68.2%) | Spiny amaranthus | Amaranthus spinosus L. | H | W | 12.8 |
| Green crumbweed | Dysphania carinata (R.Br.) Mosyakin & Clemants * | H | W | 11.9 | |
| Paddy’s lucerne | Sida rhombifolia L. | Sb | W | 11.5 | |
| Parthenium weed | Parthenium hysterophorus L. | H | W | 11.0 | |
| Green panic | Panicum maximum Jacq. | G | D | 10.1 | |
| Butterfly pea | Clitoria ternatea L. | H | D | 8.3 | |
| Rhodes grass | Chloris gayana Kunth | G | D | 5.8 | |
| Spear thistle | Cirsium vulgare (Savi) Ten. | H | W | 5.3 | |
| Bala | Sida cordifolia L. | Sb | W | 4.4 | |
| Bisset bluegrass | Bothriochloa insculpta Hochst. Ex A.Rich A.Camus | G | D | 3.6 | |
| Yellow vine | Tribulus micrococcus Domin * | H | W | 2.8 | |
| Purslane | Portulaca oleracea L. | H | W | 2.7 | |
| Two (similarity 55.4%) | Siratro | Macropitilum artropurpureum (DC.) Urb. | H | D | 16.2 |
| Buffel grass | Cenchrus ciliaris L. | G | D | 13.5 | |
| Liverseed grass | Urochloa panicoides Beauv. | G | W | 11.3 | |
| Green crumbweed | Dysphania carinata (R.Br.) Mosyakin & Clemants * | H | W | 11.1 | |
| Paddy’s lucerne | Sida rhombifolia L. | Sb | W | 10.6 | |
| Spiny amaranthus | Amaranthus spinosus L. | H | W | 8.6 | |
| Bala | Sida cordifolia L. | Sb | W | 7.3 | |
| Parthenium weed | Parthenium hysterophorus L. | H | W | 5.8 | |
| Yellow vine | Tribulus micrococcus Domin * | H | W | 3.7 | |
| Goosegrass | Eleusine indica (L.) Gaertn. | G | W | 3.0 | |
| Plant Mixture | Common Name | Scientific Name | Family | Origin | SI * | FS (%) * | Seed Rate (seed m−2) |
|---|---|---|---|---|---|---|---|
| One | Rhodes grass | Chloris gayana Kunth cv. Callide | Poaceae | Introduced | - | 79 | 245 |
| Bisset bluegrass | Bothriochloa insculpta Hochst. ex A. Rich A. Camus cv. Bisset | Poaceae | Introduced | 3.2 | 62 | 33 | |
| Butterfly pea | Clitoria ternatea L. cv. Milgarra | Fabaceae | Introduced | 2.9 | 70 | 10 | |
| Green panic | Panicum maximum Jacq. | Poaceae | Introduced | - | - | 118 | |
| Bull Mitchell grass | Astrebla squarrosa C.E. Hubb. | Poaceae | Native | 1.4 | 62 | 9 | |
| Two | Forest bluegrass | Bothriochloa bladhii (Retz.) | Poaceae | Introduced | 2.0 | - | 38 |
| Red grass | Bothriochloa macra (Steud.) S. T. Blake | Poaceae | Native | 0.9 | - | 10 | |
| Queensland bluegrass | Dichanthium sericeum (R. Br.) A. Camus | Poaceae | Native | 1.7 | 70 | 41 | |
| Siratro | Macroptilum artropurpureum (DC.) Urb. | Fabaceae | Introduced | - | - | 65 | |
| Buffel grass | Cenchrus ciliaris L. | Poaceae | Introduced | 1.8 | 76 | 100 ** | |
| Hoop Mitchell grass | Astrebla elymoides F. Muell. | Poaceae | Native | 1.1 | - | 38 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belgeri, A.; Bajwa, A.A.; Shabbir, A.; Navie, S.; Vivian-Smith, G.; Adkins, S. Managing an Invasive Weed Species, Parthenium hysterophorus, with Suppressive Plant Species in Australian Grasslands. Plants 2020, 9, 1587. https://doi.org/10.3390/plants9111587
Belgeri A, Bajwa AA, Shabbir A, Navie S, Vivian-Smith G, Adkins S. Managing an Invasive Weed Species, Parthenium hysterophorus, with Suppressive Plant Species in Australian Grasslands. Plants. 2020; 9(11):1587. https://doi.org/10.3390/plants9111587
Chicago/Turabian StyleBelgeri, Amalia, Ali Ahsan Bajwa, Asad Shabbir, Sheldon Navie, Gabrielle Vivian-Smith, and Steve Adkins. 2020. "Managing an Invasive Weed Species, Parthenium hysterophorus, with Suppressive Plant Species in Australian Grasslands" Plants 9, no. 11: 1587. https://doi.org/10.3390/plants9111587
APA StyleBelgeri, A., Bajwa, A. A., Shabbir, A., Navie, S., Vivian-Smith, G., & Adkins, S. (2020). Managing an Invasive Weed Species, Parthenium hysterophorus, with Suppressive Plant Species in Australian Grasslands. Plants, 9(11), 1587. https://doi.org/10.3390/plants9111587