Production of Flavonoids in Callus Cultures of Sophora flavescens Aiton

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. In Vitro Germination of S. flavescens
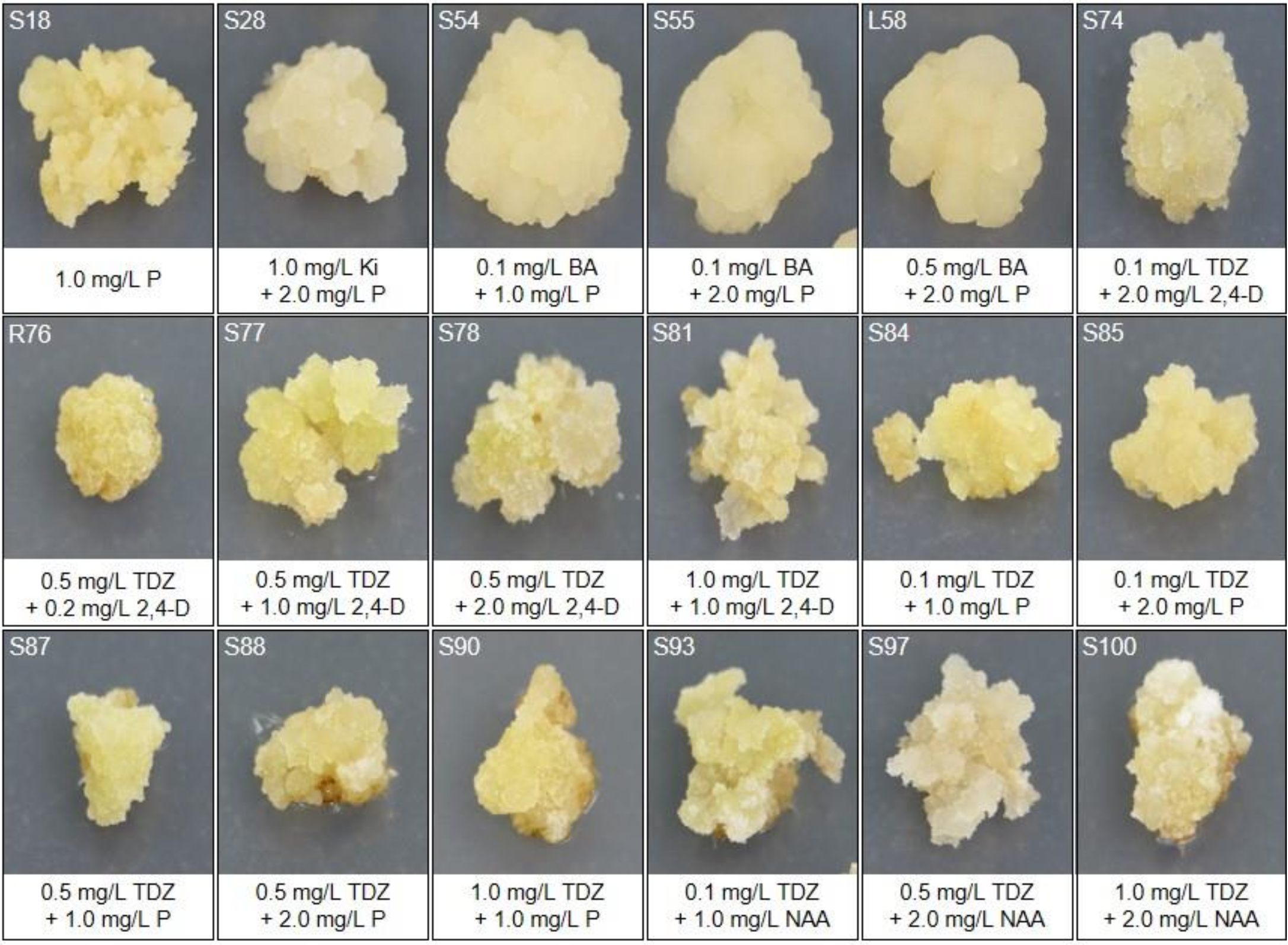
2.2. Callus Induction and Selection
2.3. Total Phenolic and Flavonoid Contents
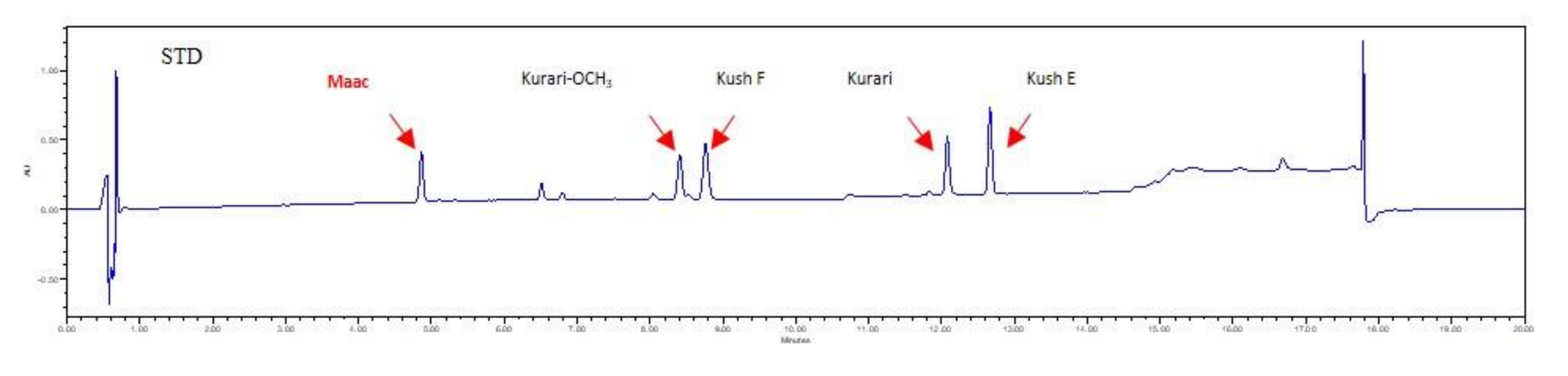
2.4. Analysis of Flavonoid Metabolites, Including Maackiain, in Calli by UHPLC
2.5. DPPH Radical Scavenging Activity
2.6. POD and SOD Activities
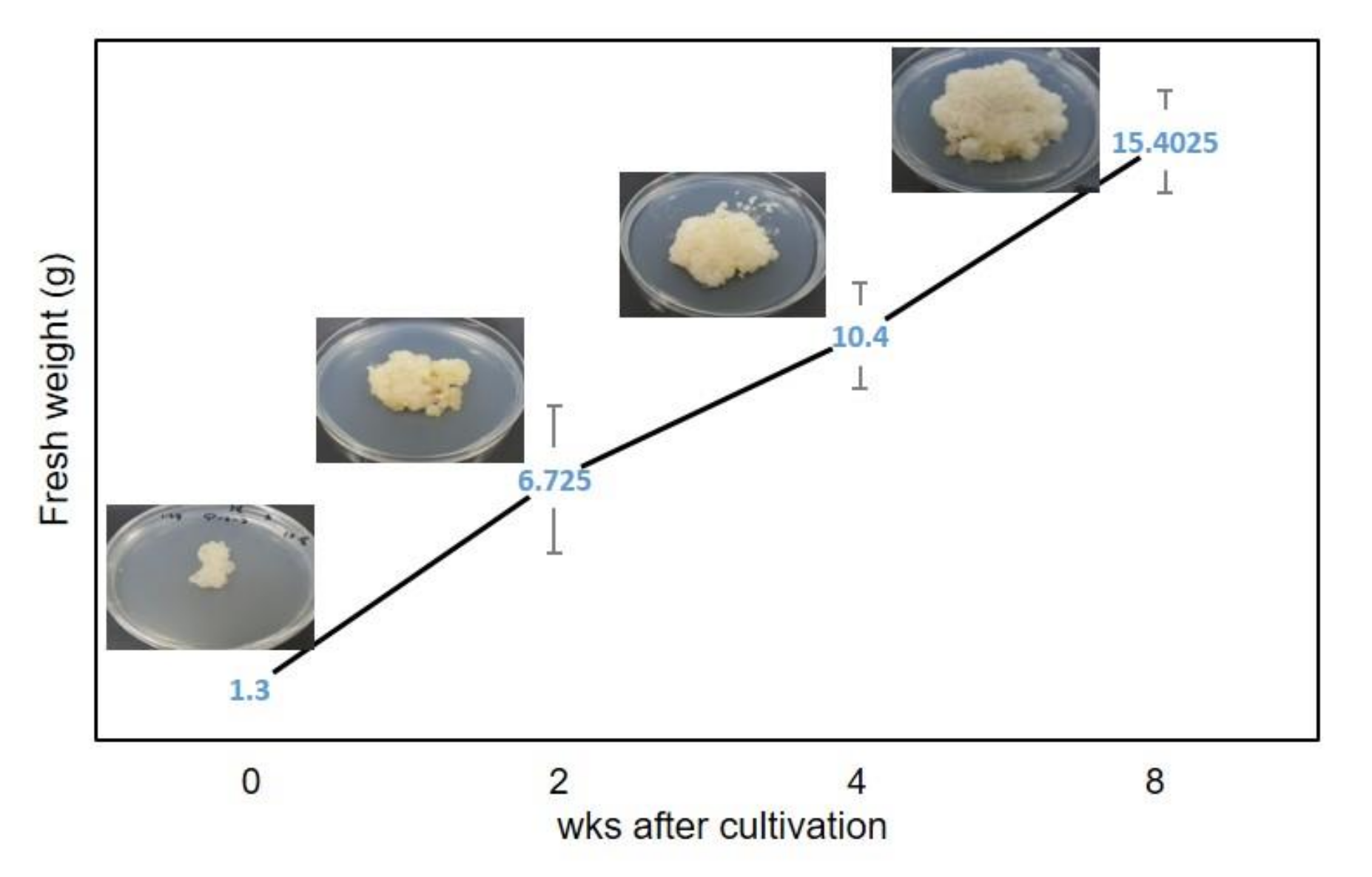
2.7. Biomass Production and Growth Kinetics
2.8. In Vitro Shoot Regeneration from the Callus
3. Discussion
4. Materials and Methods
4.1. Study Materials
4.2. In Vitro Seed Germination
4.3. Callus Induction
4.4. Analysis of Flavonoid Metabolites by Ultra-High-Performance Liquid Chromatography (UHPLC)
4.5. Total Phenolic Content
4.6. Total Flavonoid Content
4.7. DPPH Radical Scavenging Activity Determination
4.8. Peroxidase Activity
4.9. Superoxide Dismutase Activity
4.10. In Vitro Shoot Regeneration
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Phondani, P.C.; Bhatt, I.D.; Negi, V.S.; Kothyari, B.P.; Bhatt, A.; Maikhuri, R.K. Promoting medicinal plants cultivation as a tool for biodiversity conservation and livelihood enhancement in indian himalaya. J. Asia Pac. Biodivers. 2016, 9, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Fabricant, D.S.; Farnsworth, N.R. The value of plants used in traditional medicine for drug discovery. Environ. Health Perspect. 2001, 109, 69–75. [Google Scholar] [PubMed]
- Akula, R.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.-H.; Jeong, S.-J.; Ko, W.-G.; Kim, N.-Y.; Lee, B.-H.; Inagaki, M.; Miyamoto, T.; Higuchi, R.; Kim, Y.-C. Cytotoxic lavandulyl flavanones from Sophora flavescens. J. Nat. Prod. 2000, 63, 680–681. [Google Scholar] [CrossRef]
- He, X.; Fang, J.; Huang, L.; Wang, J.; Huang, X. Sophora flavescens ait.: Traditional usage, phytochemistry and pharmacology of an important traditional chinese medicine. J. Ethnopharmacol. 2015, 172, 10–29. [Google Scholar] [CrossRef]
- Han, J.-S.; Shin, D.-H. Antimicrobial effect of each solvent fraction of morus alba linne, Sophora flavescens aiton on Listeria monocytogenes. Korean J. Food Sci. Technol. 1994, 26, 539–544. [Google Scholar]
- Chen, X.; Yi, C.; Yang, X.; Wang, X. Liquid chromatography of active principles in sophora flavescens root. J. Chromatogr. B 2004, 812, 149–163. [Google Scholar] [CrossRef]
- Yamamoto, O.; Yamada, Y. Production of reserpine and its optimization in cultured Rauwolfia serpentina Benth. Cells. Plant Cell Rep. 1986, 5, 50–53. [Google Scholar] [CrossRef]
- Dias, A.C.P.; Seabra, R.M.; Andrade, P.B.; Ferreres, F.; Fernandes-Ferreira, M. Xanthone biosynthesis and accumulation in calli and suspended cells of Hypericum androsaemum. Plant Sci. 2000, 150, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Meratan, A.; Ghaffari, S.; Niknam, V. In vitro organogenesis and antioxidant enzymes activity in Acanthophyllum sordidum. Biol. Plant. 2009, 53, 5–10. [Google Scholar] [CrossRef]
- Ali, M.; Abbasi, B.H. Light-induced fluctuations in biomass accumulation, secondary metabolites production and antioxidant activity in cell suspension cultures of Artemisia absinthium L. J. Photochem. Photobiol. B Biol. 2014, 140, 223–227. [Google Scholar] [CrossRef]
- Maneechai, S.; De-Eknamkul, W.; Umehara, K.; Noguchi, H.; Likhitwitayawuid, K. Flavonoid and stilbenoid production in callus cultures of Artocarpus lakoocha. Phytochemistry 2012, 81, 42–49. [Google Scholar] [CrossRef]
- Del Pilar Nicasio-Torres, M.; Meckes-Fischer, M.; Aguilar-Santamaría, L.; Garduño-Ramírez, M.L.; Chávez-Ávila, V.M.; Cruz-Sosa, F. Production of chlorogenic acid and isoorientin hypoglycemic compounds in Cecropia obtusifolia calli and in cell suspension cultures with nitrate deficiency. Acta Physiol. Plant. 2012, 34, 307–316. [Google Scholar] [CrossRef]
- Cheng, S.; Zhang, W.; Nannan, S.; Feng, X.; Linling, L.; Yongling, L.; Cheng, H. Production of flavonoids and terpene lactones from optimized Ginkgo biloba tissue culture. Not. Bot. Horti Agrobot. Cluj Napoca 2014, 42, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Panda, B.M.; Mehta, U.J.; Hazra, S. Optimizing culture conditions for establishment of hairy root culture of Semecarpus anacardium L. 3 Biotech 2017, 7, 21. [Google Scholar] [CrossRef] [PubMed]
- Furuya, T.; Ikuta, A. The presence of l-maackiain and pterocarpin in callus tissue of sophora angustifolia. Chem. Pharm. Bull. 1968, 16, 771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, H.; Kawai, S.; Mayumi, J.; Tanaka, T.; Iinuma, M.; Mizuno, M. Prenylated flavanone production in callus cultures of Sophora flavescens var. angustifolia. Z. Nat. C 1991, 46, 172–176. [Google Scholar]
- Yamamoto, H.; Zhao, P.; Inoue, K. Origin of two isoprenoid units in a lavandulyl moiety of sophoraflavanone g from Sophora flavescens cultured cells. Phytochemistry 2002, 60, 263–267. [Google Scholar] [CrossRef]
- Thengane, S.; Kulkarni, D.; Shrikhande, V.; Joshi, S.; Sonawane, K.; Krishnamurthy, K. Influence of medium composition on callus induction and camptothecin (s) accumulation in Nothapodytes foetida. Plant Cell Tissue Organ Cult. 2003, 72, 247–251. [Google Scholar] [CrossRef]
- Luciani, G.F.; Mary, A.K.; Pellegrini, C.; Curvetto, N. Effects of explants and growth regulators in garlic callus formation and plant regeneration. Plant Cell Tissue Organ Cult. 2006, 87, 139–143. [Google Scholar] [CrossRef]
- Şener, O.; Can, E.; Arslan, M.; Çeliş, N. Effects of genotype and picloram concentrations on callus induction and plant regeneration from immature inflorescence of spring barley cultivars (Hordeum vulgare L.). Biotechnol. Biotechnol. Equip. 2008, 22, 915–920. [Google Scholar]
- Landi, L.; Mezzetti, B. Tdz, auxin and genotype effects on leaf organogenesis in Fragaria. Plant Cell Rep. 2006, 25, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Yasmin, S.; Nasiruddin, K.; Begum, R.; Talukder, S. Regeneration and establishment of potato plantlets through callus formation with bap and naa. Asian J. Plant. Sci. 2003, 2, 936–940. [Google Scholar]
- Tao, H.; Shaolin, P.; Gaofeng, D.; Lanying, Z.; Gengguang, L. Plant regeneration from leaf-derived callus in Citrus grandis (pummelo): Effects of auxins in callus induction medium. Plant Cell Tissue Organ Cult. 2002, 69, 141–146. [Google Scholar] [CrossRef]
- De Queiroz Braga, K.; Coimbra, M.C.; Castro, A.H.F. In vitro germination, callus induction and phenolic compounds contents from Pyrostegia venusta (ker gawl.) miers. Acta Sci. Biol. Sci. 2015, 37, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, A.; Chiruvella, K.K.; Rao, Y.K.; Geethangili, M.; Raghavan, S.C.; Ghanta, R.G. In vitro production of echioidinin, 7-o-methywogonin from callus cultures of Andrographis lineata and their cytotoxicity on cancer cells. PloS ONE 2015, 10, e0141154. [Google Scholar] [CrossRef] [Green Version]
- Norton, D.; Godley, E.; Heenan, P.; Ladley, J. Germination of sophora seeds after prolonged storage. N. Z. J. Bot. 2002, 40, 389–396. [Google Scholar] [CrossRef]
- Karabourniotis, G.; Liakopoulos, G.; Nikolopoulos, D.; Bresta, P.; Stavroulaki, V.; Sumbele, S. “Carbon gain vs. Water saving, growth vs. Defence”: Two dilemmas with soluble phenolics as a joker. Plant Sci. 2014, 227, 21–27. [Google Scholar] [CrossRef]
- Ahmad, N.; Fazal, H.; Abbasi, B.H.; Farooq, S. Efficient free radical scavenging activity of Ginkgo biloba, Stevia rebaudiana and Parthenium hysterophorous leaves through dpph (2, 2-diphenyl-1-picrylhydrazyl). Int. J. Phytomed. 2010, 2. [Google Scholar] [CrossRef]
- Ryu, J.; Doo, H.; Kwon, T. Induction of haploid plants by anther culture in sesame (Sesamum indicum L.)-(1)-effects of growth regulators and difference between genotypes on callus induction. Korean J. Plant Tissue Cult. 1992, 19, 171–177. [Google Scholar]
- Palacio, L.; Cantero, J.J.; Cusidó, R.M.; Goleniowski, M.E. Phenolic compound production in relation to differentiation in cell and tissue cultures of Larrea divaricata (cav.). Plant Sci. 2012, 193, 1–7. [Google Scholar] [CrossRef]
- Castro, A.H.F.; Braga, K.d.Q.; Sousa, F.M.d.; Coimbra, M.C.; Chagas, R.C.R. Callus induction and bioactive phenolic compounds production from Byrsonima verbascifolia (L.) dc. (malpighiaceae). Rev. Cienc. Agron. 2016, 47, 143–151. [Google Scholar] [CrossRef] [Green Version]
- George, E.F.; Hall, M.A.; De Klerk, G.-J. Plant Propagation by Tissue Culture, 3rd ed.; Springer: Berlin, Germany, 2008; Volume 1, pp. 175–226. [Google Scholar]
- Al-Ajlouni, Z.I.; Abbas, S.; Shatnawi, M.; Al-Makhadmeh, I. In vitro propagation, callus induction, and evaluation of active compounds on Ruta graveolens. J. Food Agric. Environ. 2015, 13, 101–106. [Google Scholar]
- Farhadi, N.; Panahandeh, J.; Azar, A.M.; Salte, S.A. Effects of explant type, growth regulators and light intensity on callus induction and plant regeneration in four ecotypes of persian shallot (Allium hirtifolium). Sci. Hortic. 2017, 218, 80–86. [Google Scholar] [CrossRef]
- Farvardin, A.; Ebrahimi, A.; Hosseinpour, B.; Khosrowshahli, M. Effects of growth regulators on callus induction and secondary metabolite production in Cuminum cyminum. Nat. Prod. Res. 2017, 31, 1963–1970. [Google Scholar] [CrossRef]
- Anjusha, S.; Gangaprasad, A. Callus culture and in vitro production of anthraquinone in Gynochthodes umbellata (L.) Razafim. & B. Bremer (Rubiaceae). Ind. Crop. Prod. 2017, 95, 608–614. [Google Scholar]
- Karakas, F.P.; Turker, A.U. Improvement of shoot proliferation and comparison of secondary metabolites in shoot and callus cultures of phlomis armeniaca by lc-esi-ms/ms analysis. In Vitro Cell. Dev. Biol. Plant 2016, 52, 608–618. [Google Scholar] [CrossRef]
- Mahendran, G.; Bai, V.N. Micropropagation, antioxidant properties and phytochemical assessment of Swertia corymbosa (Griseb.) wight ex cb clarke: A medicinal plant. Acta Physiol. Plant. 2014, 36, 589–603. [Google Scholar] [CrossRef]
- Pasqua, G.; Avato, P.; Monacelli, B.; Santamaria, A.R.; Argentieri, M.P. Metabolites in cell suspension cultures, calli, and in vitro regenerated organs of Hypericum perforatum cv. Topas. Plant Sci. 2003, 165, 977–982. [Google Scholar] [CrossRef]
- Ohlsson, A.B. Effects of abscisic acid on cardenolide accumulation and growth in Digitalis lanata tissue cultures. J. Plant Physiol. 1990, 136, 510–512. [Google Scholar] [CrossRef]
- Hagimori, M.; Matsumoto, T.; Obi, Y. Studies on the production of Digitalis cardenolides by plant tissue culture: Ii. Effect of light and plant growth substances on digitoxin formation by undifferentiated cells and shoot-forming cultures of digitalis purpurea L. Grown in liquid media. Plant Physiol. 1982, 69, 653–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Chaturvedi, R. Sustainable production of azadirachtin from differentiated in vitro cell lines of neem (Azadirachta indica). AoB Plants 2013, 5. [Google Scholar] [CrossRef]
- Prakash, G.; Bhojwani, S.S.; Srivastava, A.K. Production of azadirachtin from plant tissue culture: State of the art and future prospects. Biotechnol. Bioprocess Eng. 2002, 7, 185–193. [Google Scholar] [CrossRef]
- Wewetzer, A. Callus cultures Ofazadirachta indica and their potential for the production of azadirachtin. Phytoparasitica 1998, 26, 47–52. [Google Scholar] [CrossRef]
- Loredo-Carrillo, S.E.; de Lourdes Santos-Díaz, M.; Leyva, E.; del Socorro Santos-Díaz, M. Establishment of callus from Pyrostegia venusta (Ker Gawl.) miers and effect of abiotic stress on flavonoids and sterols accumulation. J. Plant Biochem. Biotechnol. 2013, 22, 312–318. [Google Scholar] [CrossRef]
- Zhao, D.; Guo, G.; Wang, X.; Zheng, G. In vitro micropropagation of a medicinal plant species Sophora flavescens. Biol. Plant. 2003, 47, 117–120. [Google Scholar] [CrossRef]
- Jana, S.; Sivanesan, I.; Jeong, B.R. Effect of cytokinins on in vitro multiplication of Sophora tonkinensis. Asian Pac. J. Trop. Biomed. 2013, 3, 549–553. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Slinkard, K.; Singleton, V.L. Total phenol analysis: Automation and comparison with manual methods. Am. J. Enol. Vitic. 1977, 28, 49–55. [Google Scholar]
- Chang, C.-C.; Yang, M.-H.; Wen, H.-M.; Chern, J.-C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug. Anal. 2002, 10, 178–182. [Google Scholar]
- Tiwari, P.; Kumar, B.; Kaur, M.; Kaur, G.; Kaur, H. Phytochemical screening and extraction: A review. Int. Pharm. Sci. 2011, 1, 98–106. [Google Scholar]
- Abate, L.; Abebe, A.; Mekonnen, A. Studies on antioxidant and antibacterial activities of crude extracts of Plantago lanceolata leaves. Chem. Int. 2017, 3, 277–287. [Google Scholar]
- Nayyar, H.; Gupta, D. Differential sensitivity of c3 and c4 plants to water deficit stress: Association with oxidative stress and antioxidants. Environ. Exp. Bot. 2006, 58, 106–113. [Google Scholar] [CrossRef]
- Khan, T.; Khan, T.; Hano, C.; Abbasi, B.H. Effects of chitosan and salicylic acid on the production of pharmacologically attractive secondary metabolites in callus cultures of Fagonia indica. Ind. Crop. Prod. 2019, 129, 525–535. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Callus Type | TPC (mg GAE/g FW) | TFC (mg QE/g FW) | DPPH (%) | SOD (nM/min/mgFW) | POD (nM/min/mgFW) | |
|---|---|---|---|---|---|---|
| Root * | - | 7.82 ± 0.52a | 9.75 ± 0.18a | 84.00 ± 6.56a | 0.71 ± 0.04a | 5.60 ± 0.43a |
| Root (in vitro) ** | - | 5.09 ± 0.28bc | 5.80 ± 0.39b | 70.67 ± 8.02ab | 0.68 ± 0.04a | 5.07 ± 0.29a |
| L58 (0.5 mg/L BA + 2.0 mg/L picloram) | LY/JS | 6.00 ± 0.30b | 5.40 ± 0.56b | 62.67 ± 7.09b | 0.42 ± 0.04b | 4.10 ± 1.31ab |
| R76 (0.5 mg/L TDZ + 0.2 mg/L 2,4-D) | GY/F | 4.93 ± 0.59bc | 3.22 ± 0.67c | 57.00 ± 7.00b | 0.38 ± 0.06bc | 2.37 ± 0.23bc |
| S18 (1.0 mg/L Picloram) | Y/F | 3.99 ± 0.28c | 4.96 ± 0.33b | 32.00 ± 5.29c | 0.24 ± 0.04c | 1.22 ± 0.59c |
| S55 (0.1 mg/L BA + 2.0 mg/L picloram) | Y/JS | 5.93 ± 0.44b | 3.61 ± 0.17c | 55.00 ± 4.58b | 0.25 ± 0.07c | 3.13 ± 0.47b |
| S88 (0.5 mg/L TDZ + 2.0 mg/L picloram) | Y/JS | 5.31 ± 1.28bc | 3.91 ± 0.19c | 53.00 ± 8.54b | 0.33 ± 0.04bc | 2.67 ± 0.55bc |
| Maackiain | Kurari-Methoxy | Kush F | Kurari | Kush E | |
|---|---|---|---|---|---|
| Root * | 145.9a | 468.6b | 1621.4d | 299.1a | 0 |
| Root (in vitro) ** | 123.3a | 69.2a | 933.1c | 781.3b | 0 |
| L58 | 1580.6d | 0 | 229.1b | 0 | 0 |
| R76 | 653.7c | 0 | 0 | 0 | 0 |
| S18 | 428.3b | 0 | 0 | 0 | 0 |
| S55 | 592.4c | 0 | 182.6b | 0 | 0 |
| S88 | 395.9b | 0 | 14.8a | 0 | 0 |
| Plant Growth Regulators (mg/L) | Shoot Numbers Per Callus | Rooting Rates (%) | |
|---|---|---|---|
| TDZ | NAA | ||
| 0.1 | 0.2 | 4.0 ± 0.68a | 25.0 ± 4.52b |
| 1.0 | 3.1 ± 0.90a | 66.0 ± 14.6d | |
| 2.0 | 9.0 ± 1.08c | 11.0 ± 1.37a | |
| 0.5 | 0.2 | 6.0 ± 0.97b | 66.0 ± 9.88d |
| 1.0 | 4.1 ± 0.58a | 25.0 ± 7.72b | |
| 2.0 | 18.0 ± 1.53d | 16.0 ± 8.10a | |
| 1.0 | 0.2 | 6.0 ± 1.18b | 66.7 ± 5.06d |
| 1.0 | 3.0 ± 0.97a | 33.3 ± 12.48c | |
| 2.0 | 4.2 ± 0.64a | 75.0 ± 6.27e | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.-S.; Seong, Z.-K.; Kim, M.-S.; Ha, J.-H.; Moon, K.-B.; Lee, H.-J.; Lee, H.-K.; Jeon, J.-H.; Park, S.U.; Kim, H.-S. Production of Flavonoids in Callus Cultures of Sophora flavescens Aiton. Plants 2020, 9, 688. https://doi.org/10.3390/plants9060688
Park J-S, Seong Z-K, Kim M-S, Ha J-H, Moon K-B, Lee H-J, Lee H-K, Jeon J-H, Park SU, Kim H-S. Production of Flavonoids in Callus Cultures of Sophora flavescens Aiton. Plants. 2020; 9(6):688. https://doi.org/10.3390/plants9060688
Chicago/Turabian StylePark, Ji-Sun, Zuh-Kyung Seong, Mi-Sun Kim, Jang-Ho Ha, Ki-Beom Moon, Hyo-Jun Lee, Hyeong-Kyu Lee, Jae-Heung Jeon, Sang Un Park, and Hyun-Soon Kim. 2020. "Production of Flavonoids in Callus Cultures of Sophora flavescens Aiton" Plants 9, no. 6: 688. https://doi.org/10.3390/plants9060688
APA StylePark, J. -S., Seong, Z. -K., Kim, M. -S., Ha, J. -H., Moon, K. -B., Lee, H. -J., Lee, H. -K., Jeon, J. -H., Park, S. U., & Kim, H. -S. (2020). Production of Flavonoids in Callus Cultures of Sophora flavescens Aiton. Plants, 9(6), 688. https://doi.org/10.3390/plants9060688