The Molecular and Functional Characterization of the Durum Wheat Lipoxygenase TdLOX2 Suggests Its Role in Hyperosmotic Stress Response



Abstract
:1. Introduction
2. Results and Discussion
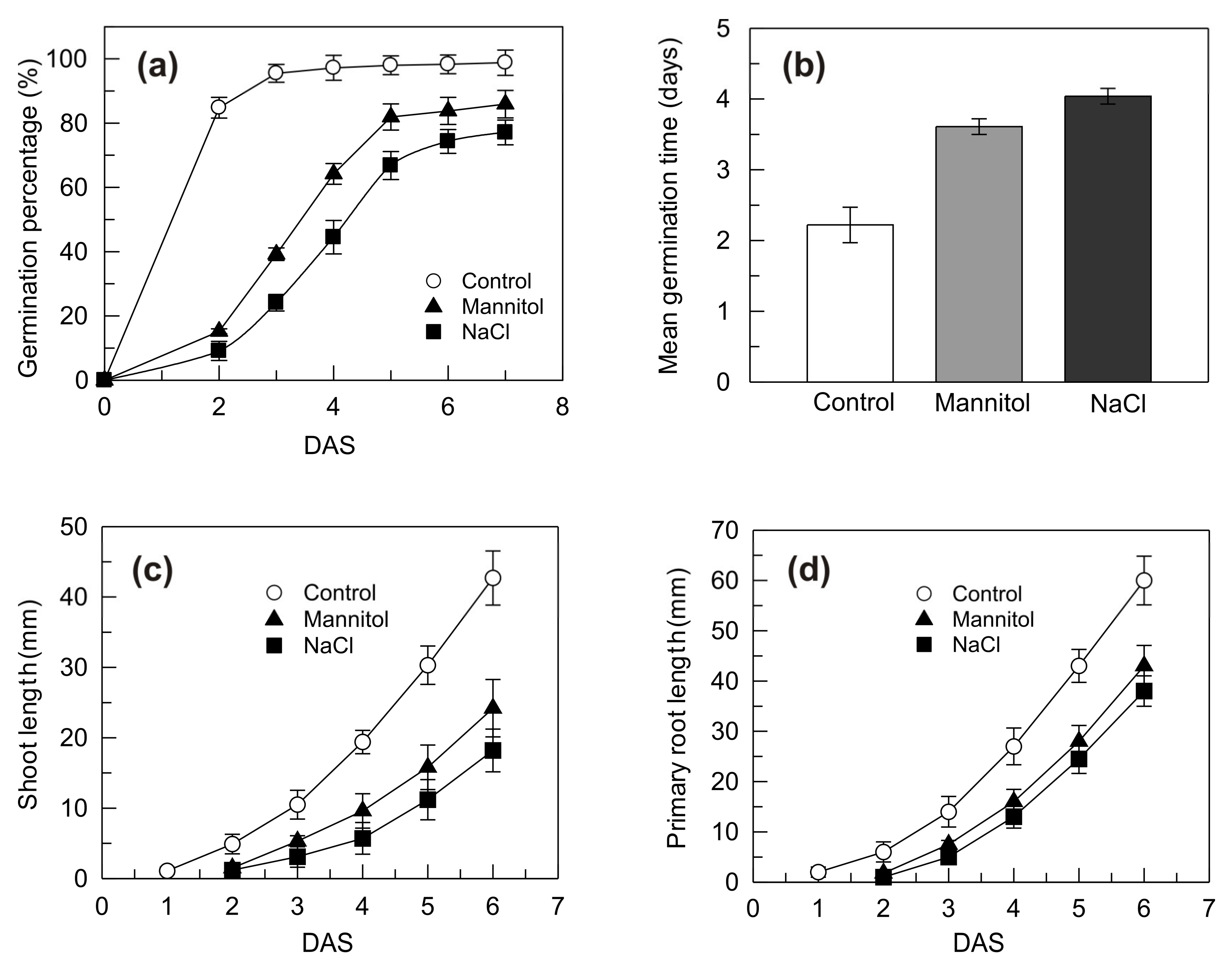
2.1. Growth Performances of Seedlings Grown under Control, Mannitol and NaCl Stress Conditions
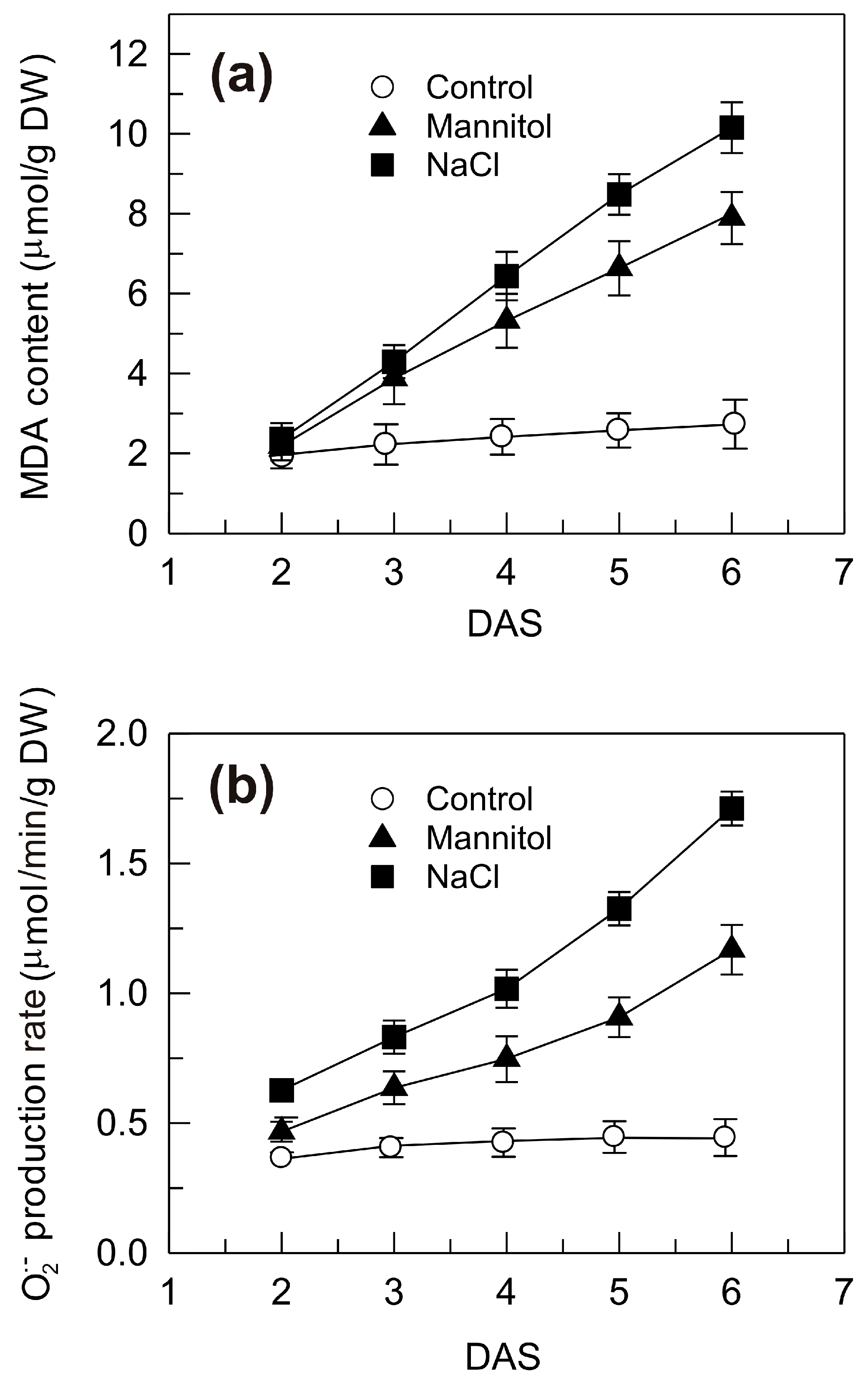
2.2. Malondialdehyde Accumulation and Superoxide Anion Generation Rate in the Shoots of Seedlings Grown under Control, Mannitol and NaCl Stress Conditions
2.3. Expression Levels of the TdLpx Genes in the Shoots of Seedlings Grown under Control, Mannitol and NaCl Stress Conditions
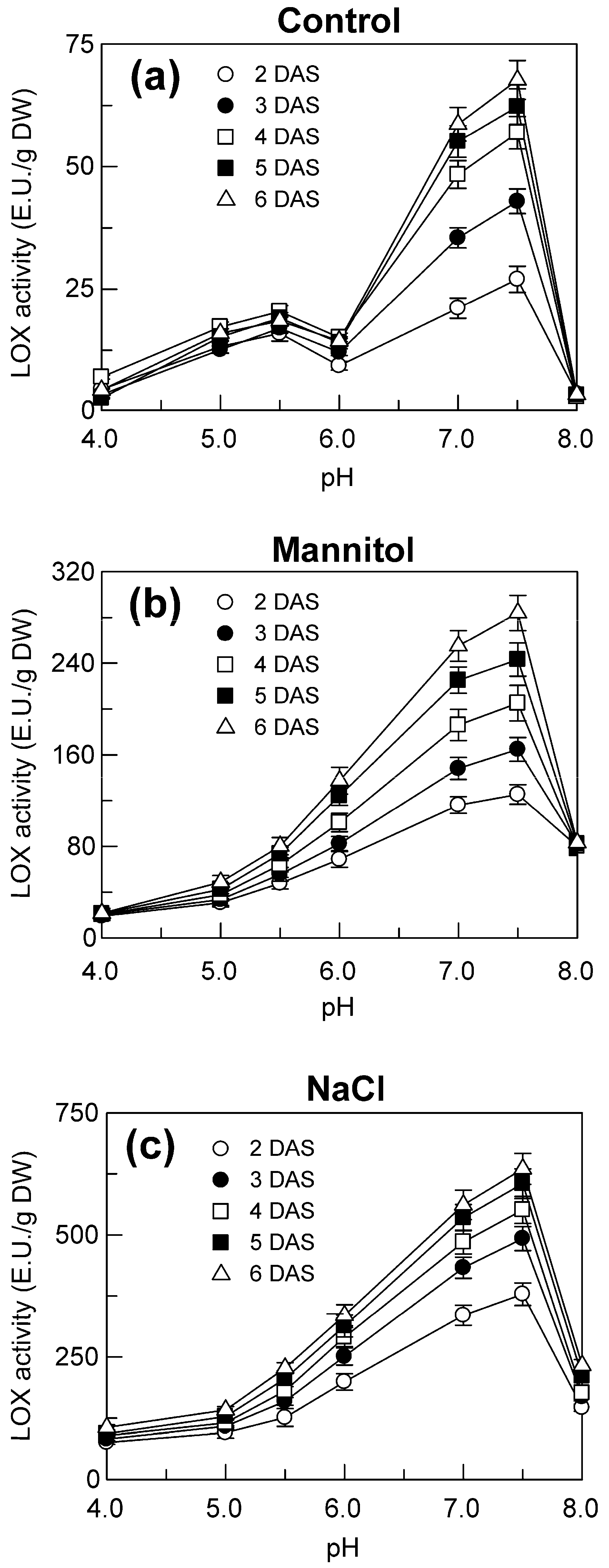
2.4. LOX Activity in the Shoots of Seedlings Grown under Control, Mannitol and NaCl Stress
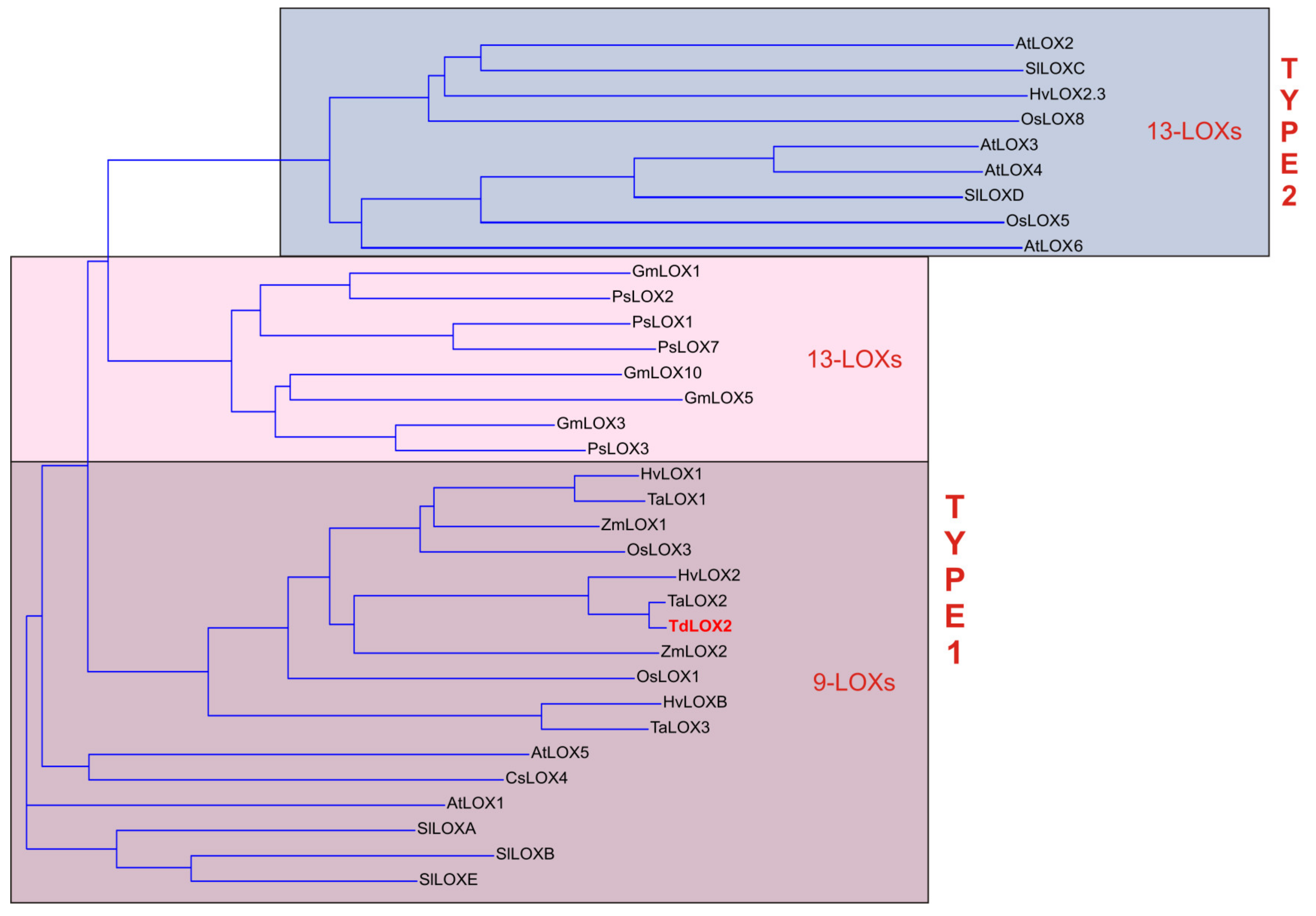
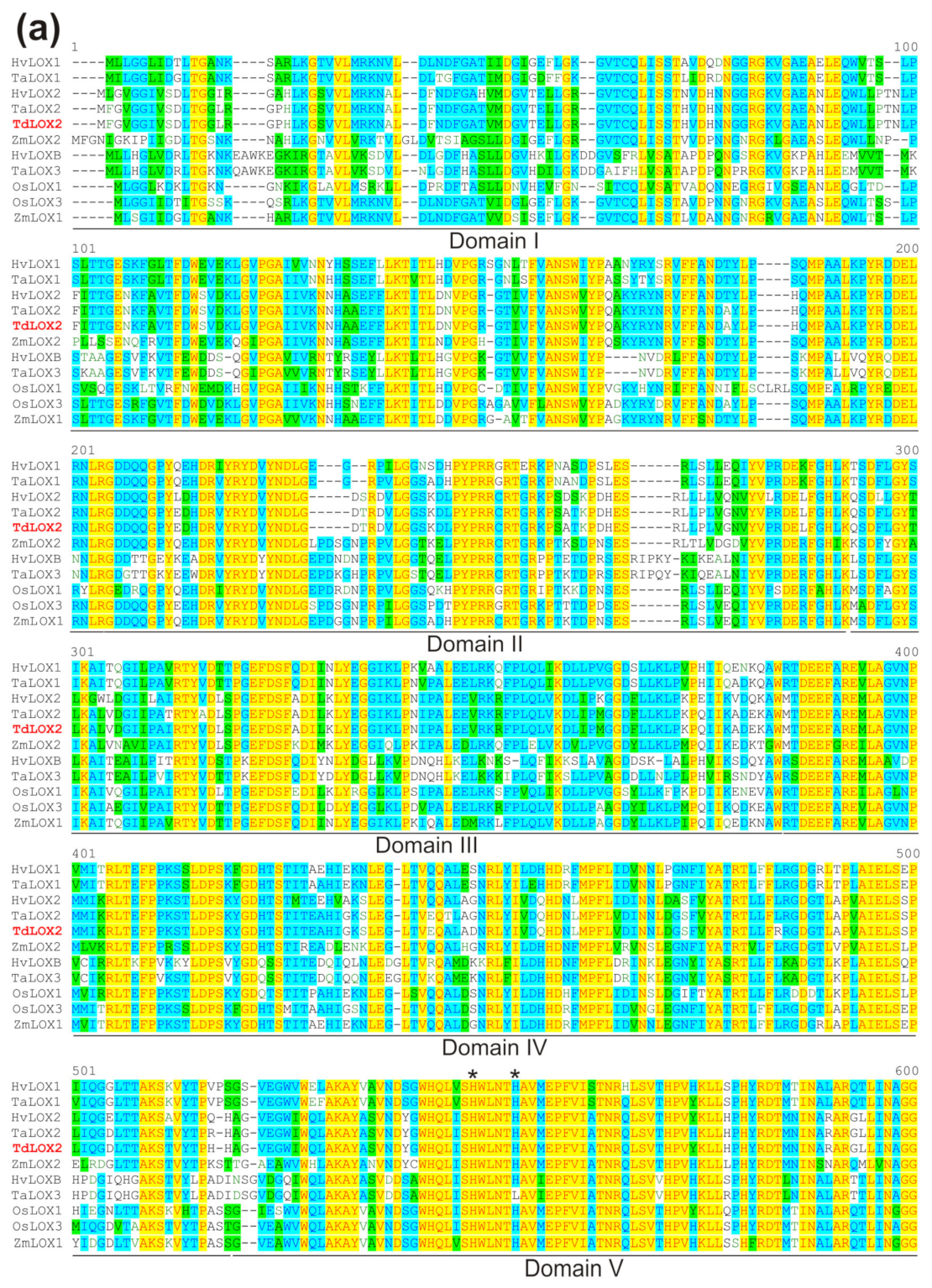
2.5. Isolation and Characterization of the Full-Length TdLpx-A2 Transcript
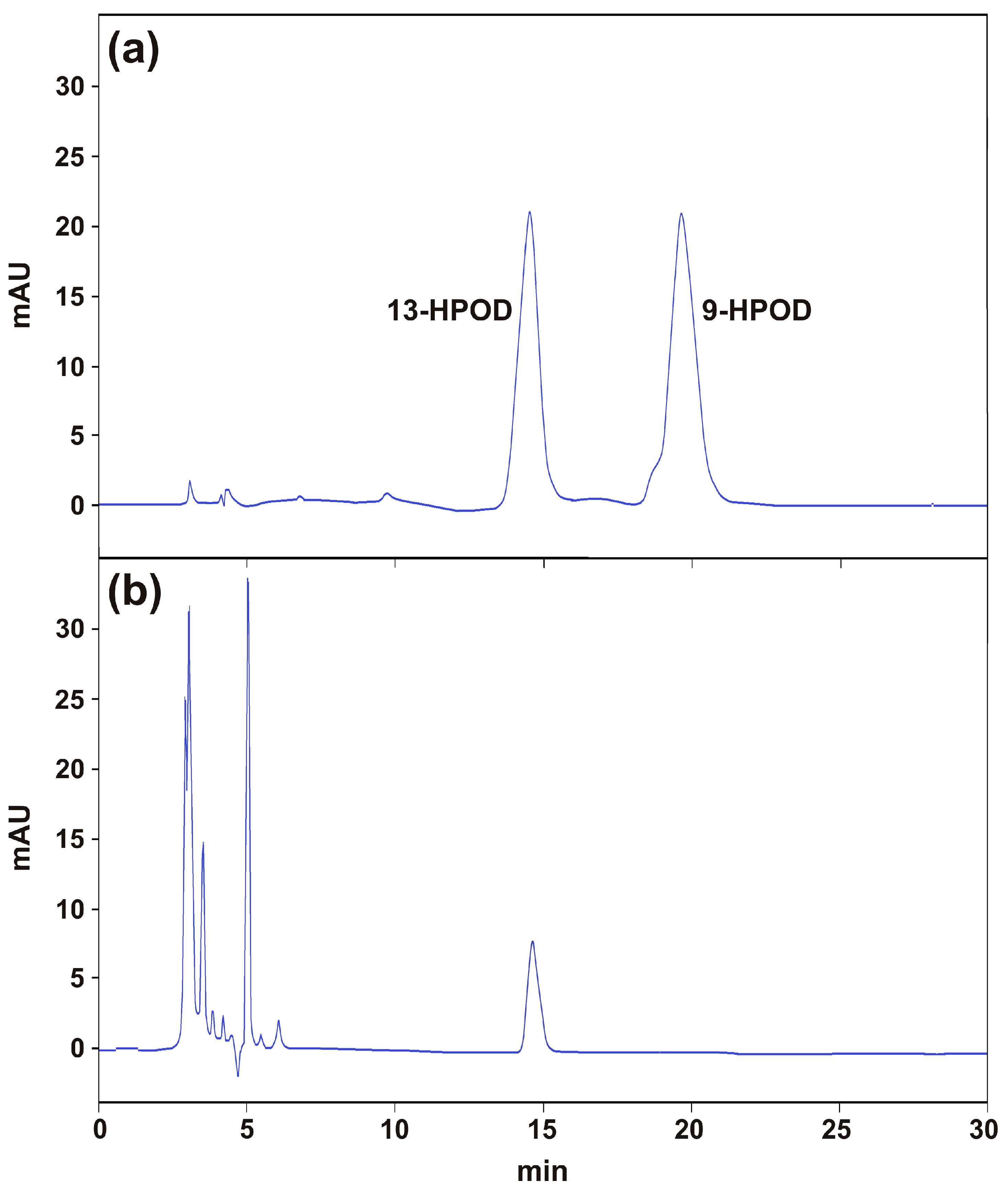
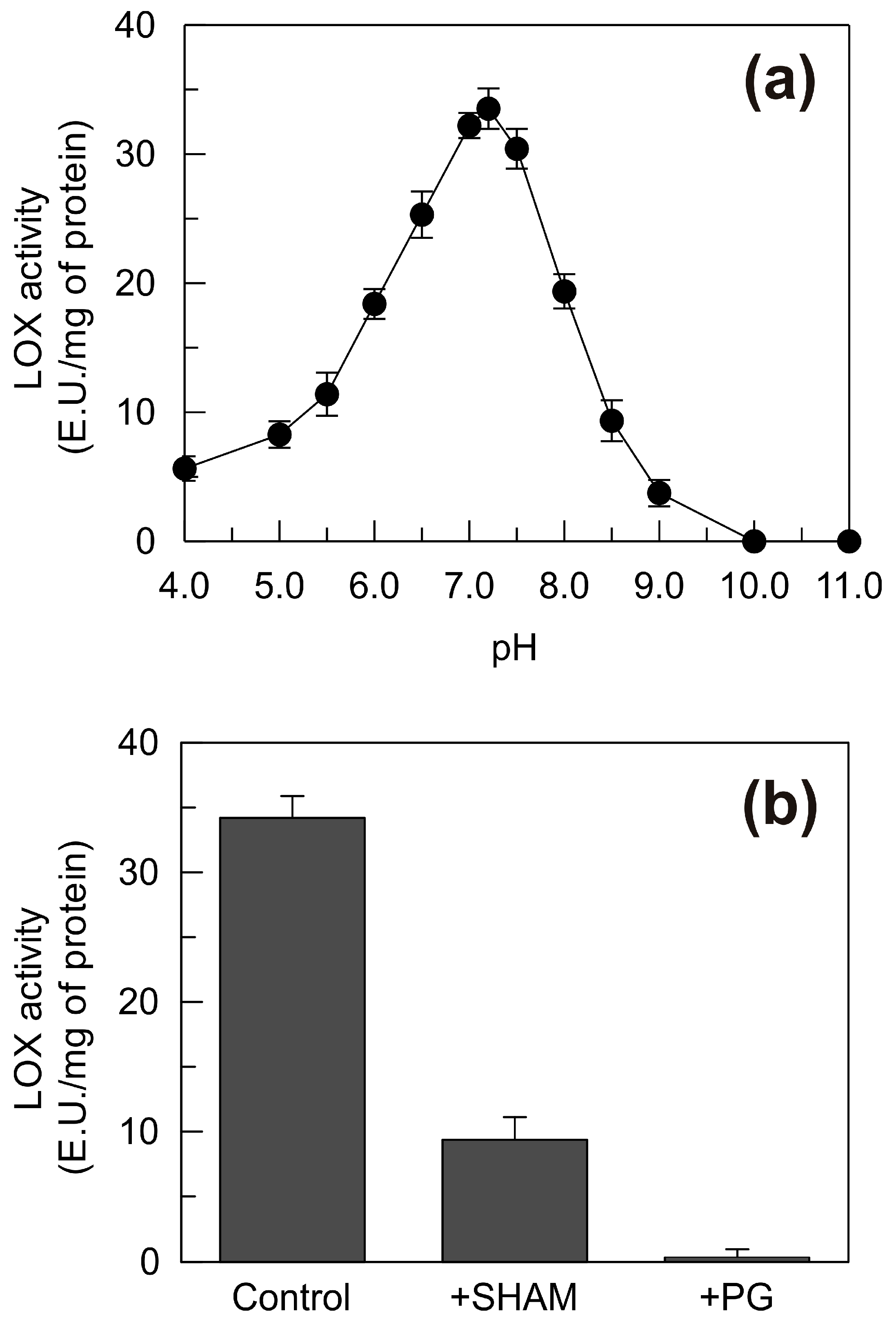
2.6. Functional Characterization of the Recombinant TdLOX2
2.7. Effect of PG on the LOX Activity, the MDA Content and the Rate of Superoxide Anion Generation in the Shoots of Seedlings Grown under Control, Mannitol and NaCl Stress Conditions
3. Materials and Methods
3.1. Growth of Control and Stressed Seedlings
3.2. Seed Germination and Seedling Growth Measurements
3.3. Measurement of the MDA Content
3.4. Measurement of the Superoxide Anion Generation Rate
3.5. Expression Analysis of the TdLpx Genes
3.6. Isolation of the Full-Length TdLpx-A2 Transcript
3.7. Heterologous Expression of the TdLOX2 in E. coli
3.8. Measurement of the LOX Activity
3.9. Analysis of the TdLOX2 Reaction Products
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pasternak, D.; Malach, Y.D. Crop irrigation with saline water. In Handbook of Plant and Crop Stress; Pessarakli, M., Ed.; Marcel Dekker: New York, NY, USA, 1994; pp. 599–622. [Google Scholar]
- Greenway, H.; Munns, R. Mechanisms of salt tolerance in nonhalophytes. Annu. Rev. Plant Physiol. 1980, 31, 149–190. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Hu, Y.; Fromm, J.; Schmidhalter, U. Effect of salinity on tissue architecture in expanding wheat leaves. Planta 2005, 220, 838–848. [Google Scholar] [CrossRef]
- Alscher, R.G.; Donahue, J.L.; Cramer, C.L. Reactive oxygen species and antioxidants: Relationships in green cells. Physiol. Plant. 1997, 1001, 224–233. [Google Scholar] [CrossRef]
- Siedow, J.N. Plant lipoxygenase: Structure and function. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 145–188. [Google Scholar] [CrossRef]
- Feussner, I.; Wasternack, C. The lipoxygenase pathway. Annu. Rev. Plant Biol. 2002, 53, 275–297. [Google Scholar] [CrossRef]
- Lee, D.-S.; Nioche, P.; Hamberg, M.; Raman, C. Structural insights into the evolutionary paths of oxylipin biosynthetic enzymes. Nature 2008, 455, 363–368. [Google Scholar] [CrossRef]
- Kolomiets, M.; Hannapel, D.; Chen, H.; Tymeson, M.; Gladon, R. Lipoxygenase is involved in the control of potato tuber development. Plant Cell 2001, 13, 613–626. [Google Scholar] [CrossRef]
- Santino, A.; Iannacone, R.; Hughes, R.; Casey, R.; Mita, G. Cloning and characterisation of an almond 9-lipoxygenase expressed early during seed development. Plant Sci. 2005, 168, 699–706. [Google Scholar] [CrossRef]
- Vellosillo, T.; Martinez, M.; Lopez, M.A.; Vicente, J.; Cascon, T.; Dolan, L.; Hamberg, M.; Castresana, C. Oxylipins produced by the 9-lipoxygenase pathway in Arabidopsis regulate lateral root development and defense responses through a specific signaling cascade. Plant Cell 2007, 19, 831–846. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Starr, J.; Göbel, C.; Engelberth, J.; Feussner, I.; Tumlinson, J.; Kolomiets, M. Maize 9-lipoxygenase ZmLOX3 controls development, root-specific expression of defense genes, and resistance to root-knot nematodes. Mol. Plant Microbe Interact. 2008, 21, 98–109. [Google Scholar] [CrossRef] [Green Version]
- León-Morcillo, R.J.; Ocampo, J.A.; García Garrido, J.M. Plant 9-lox oxylipin metabolism in response to arbuscular mycorrhiza. Plant Signal. Behav. 2012, 7, 1584–1588. [Google Scholar] [CrossRef] [Green Version]
- Porta, H.; Figueroa-Balderas, R.E.; Rocha-Sosa, M. Wounding and pathogen infection induce a chloroplast-targeted lipoxygenase in the common bean (Phaseolus vulgaris L.). Planta 2008, 227, 363–373. [Google Scholar] [CrossRef]
- Prasad, A.; Sedlářová, M.; Kale, R.S.; Pospíšil, P. Lipoxygenase in singlet oxygen generation as a response to wounding: In vivo imaging in Arabidopsis thaliana. Sci. Rep. 2017, 7, 9831. [Google Scholar] [CrossRef]
- Cacas, J.L.; Vailleau, F.; Devoine, C.; Ennar, N.; Agnel, J.P.; Tronchet, M.; Ponchet, M.; Blein, J.P.; Roby, D.; Triantaphylides, C.; et al. The combined action of 9 lipoxygenase and galactolipase is sufficient to bring about programmed cell death during tobacco hypersensitive response. Plant Cell Environ. 2005, 28, 1367–1378. [Google Scholar] [CrossRef]
- Hwang, I.S.; Hwang, B.K. The pepper 9-lipoxygenase gene CaLOX1 functions in defense and cell death responses to microbial pathogens. Plant Physiol. 2010, 152, 948–967. [Google Scholar] [CrossRef] [Green Version]
- Bae, K.S.; Rahimi, S.; Kim, Y.J.; Devi, B.S.R.; Khorolragchaa, A.; Sukweenadhi, J.; Silva, J.; Myagmarjav, D.; Yang, D.C. Molecular characterization of lipoxygenase genes and their expression analysis against biotic and abiotic stresses in Panax ginseng. Eur. J. Plant Pathol. 2016, 145, 331–343. [Google Scholar] [CrossRef]
- Lim, C.W.; Han, S.-W.; Hwang, I.S.; Kim, D.S.; Hwang, B.K.; Lee, S.C. The pepper lipoxygenase CaLOX1 plays a role in osmotic, drought and high salinity stress response. Plant Cell Physiol. 2015, 56, 930–942. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Meng, K.; Han, Y.; Ban, Q.; Wang, B.; Suo, J.; Lv, J.; Rao, J. The persimmon 9-lipoxygenase gene DkLOX3 plays positive roles in both promoting senescence and enhancing tolerance to abiotic stress. Front. Plant Sci. 2015, 6, 1073. [Google Scholar] [CrossRef] [Green Version]
- Shaban, M.; Ahmed, M.M.; Sun, H.; Ullah, A.; Zhu, L. Genome-wide identification of lipoxygenase gene family in cotton and functional characterization in response to abiotic stresses. BMC Genom. 2018, 19, 599. [Google Scholar] [CrossRef]
- Wang, C.; Gao, G.; Cao, S.; Xie, Q.; Qi, H. Isolation and functional validation of the CmLOX08 promoter associated with signalling molecule and abiotic stress responses in oriental melon, Cucumis melo var. makuwa Makino. BMC Plant Biol. 2019, 19, 75. [Google Scholar] [CrossRef] [Green Version]
- Maccarrone, M.; Veldinka, G.A.; Finazzi Agrò, A.; Vliegenthart, J.F.G. Modulation of soybean lipoxygenase expression and membrane oxidation by water deficit. FEBS Lett. 1995, 371, 223–226. [Google Scholar]
- Sofo, A.; Dichio, B.; Xiloyannis, C.; Masia, A. Lipoxygenase activity and proline accumulation in leaves and roots of olive trees in response to drought stress. Physiol. Plant. 2004, 121, 58–65. [Google Scholar] [CrossRef]
- Rodríguez-Rosales, M.P.; Kerkeb, L.; Bueno, P.; Donaire, J.P. Changes induced by NaCl in lipid content and composition, lipoxygenase, plasma membrane H+-ATPase and antioxidant enzyme activities of tomato (Lycopersicon esculentum. Mill) calli. Plant Sci. 1999, 143, 143–150. [Google Scholar]
- Mostofa, M.G.; Hossain, M.A.; Fujita, M. Trehalose pretreatment induces salt tolerance in rice (Oryza sativa L.) seedlings: Oxidative damage and co-induction of antioxidant defense and glyoxalase systems. Protoplasma 2015, 252, 461–475. [Google Scholar] [CrossRef]
- Delaplace, P.; Frettinger, P.; Ghanem, M.E.; Blondiaux, A.; Bauwens, J.; Cotton, S.; De Clerck, C.; Dewalque, A.; Guy, J.; Heuze, F. Lipoxygenase pathway and antioxidant system in salt stressed tomato seedlings (Lycopersicon esculentum Mill.). Biotechnol. Agron. Soc. Environ. 2009, 13, 529–536. [Google Scholar]
- Mittova, V.; Tal, M.; Volokita, M.; Guy, M. Salt stress induces up-regulation of an efficient chloroplast antioxidant system in the salt-tolerant wild tomato species Lycopersicon pennellii but not in the cultivated species. Physiol. Plant. 2002, 115, 393–400. [Google Scholar] [CrossRef]
- Rana, G.; Katerji, N. Measurement and estimation of actual evapotranspiration in the field under Mediterranean climate: A review. Eur. J. Agron. 2000, 13, 125–153. [Google Scholar] [CrossRef]
- Caliandro, A.; Cucci, G.; De Caro, A.; Cordella, S. Irrigation with brackish water: Influence of the irrigation regime on salt built-up in the soil and leaching effect of rainfall. In Proceedings of the European Mediterranean Conference on the Use of Saline Water in Irrigation, Bari, Italy, 25–26 July 1991. [Google Scholar]
- Munns, R.; James, R.A.; Lauchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef] [Green Version]
- Van Mechelen, J.R.; Smits, M.; Douma, A.C.; Rouster, J.; Cameron-Mills, V.; Heidekamp, F.; Valk, B.E. Primary structure of a lipoxygenase from barley grain as deduced from its cDNA sequence. Biochim. Biophys. Acta 1995, 1254, 221–225. [Google Scholar] [CrossRef]
- Van Mechelen, J.R.; Schuurink, R.C.; Smits, M.; Graner, A.; Douma, A.C.; Sedee, N.J.A.; Schmitt, N.F.; Valk, B.E. Molecular characterization of two lipoxygenases from barley. Plant Mol. Biol. 1999, 39, 1283–1289. [Google Scholar] [CrossRef] [PubMed]
- Holtman, W.L.; van Duijn, G.; Sedee, N.J.A.; Douma, A.C. Differential expression of lipoxygenase isoenzymes in embryos of germinating barley. Plant Physiol. 1996, 111, 569–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrera, A.; Echenique, V.; Zhang, W.; Helguera, M.; Manthey, F.; Schrager, A.; Picca, A.; Cervigni, G.; Dubcovsky, J. A deletion at the Lpx-B1 locus is associated with low lipoxygenase activity and improved pasta color in durum wheat, Triticum turgidum ssp. durum. J. Cereal Sci. 2007, 45, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Verlotta, A.; De Simone, V.; Mastrangelo, A.M.; Cattivelli, L.; Papa, R.; Trono, D. Insight durum wheat Lpx-B1: A small gene family coding for Lipoxygenase (LOX) responsible for carotenoid bleaching in mature grains. BMC Plant Biol. 2010, 10, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbus, I.; Soresi, D.; Romero, J.; Echenique, V. Identification, mapping and evolutionary course of wheat lipoxygenase-1 genes located on the A genome. J. Cereal Sci. 2013, 58, 298–304. [Google Scholar] [CrossRef]
- Pastore, D.; Trono, D.; Padalino, L.; Simone, S.; Valenti, D.; Di Fonzo, N.; Passerella, S. Inhibition by α-tocopherol and L-ascorbate of linoleate hydroperoxidation and β-carotene bleaching activities in durum wheat semolina. J. Cereal Sci. 2000, 31, 41–54. [Google Scholar] [CrossRef]
- Borrelli, G.M.; Fragasso, M.; Nigro, F.; Platani, C.; Papa, R.; Beleggia, R.; Trono, D. Analysis of metabolic and mineral changes in response to salt stress in durum wheat (Triticum turgidum ssp. durum) genotypes, which differ in salinity tolerance. Plant Physiol. Biochem. 2018, 133, 57–70. [Google Scholar] [CrossRef]
- Borrelli, G.M.; Ficco, D.B.M.; Giuzio, L.; Pompa, M.; Cattivelli, L.; Flagella, Z. Durum wheat salt tolerance in relation to physiological, yield and quality characters. Cereal Res. Commun. 2011, 39, 525–534. [Google Scholar] [CrossRef]
- Almansouri, M.; Kinet, J.-M.; Lutts, S. Effect of salt and osmotic stresses on germination in durum wheat (Triticum durum Desf.). Plant Soil 2001, 231, 243–254. [Google Scholar] [CrossRef]
- Flagella, Z.; Trono, D.; Pompa, M.; Di Fonzo, N.; Pastore, D. Seawater stress applied at germination affects mitochondrial function in durum wheat (Triticum durum) early seedlings. Funct. Plant Biol. 2006, 33, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Nagel, M.; Navakode, S.; Scheibal, V.; Baum, M.; Nachit, M.; Röder, M.S.; Börner, A. The genetic basis of durum wheat germination and seedling growth under osmotic stress. Biol. Plant. 2014, 58, 681–688. [Google Scholar] [CrossRef]
- Yildirim, M.; Kizilgexi, F.; Akinci, C.; Albayrak, O. Response of durum wheat seedlings to salinity. Not. Bot. Horti Agrobot. 2015, 43, 108–112. [Google Scholar] [CrossRef] [Green Version]
- Alom, R.; Hasan, M.A.; Islam, M.R.; Wang, Q.-F. Germination characters and early seedling growth of wheat (Triticum aestivum L.) genotypes under salt stress conditions. J. Crop Sci. Biotechnol. 2016, 19, 383–392. [Google Scholar] [CrossRef]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef]
- Alché, J.D. A concise appraisal of lipid oxidation and lipoxidation in higher plants. Redox Biol. 2019, 23, 101136. [Google Scholar] [CrossRef]
- Bali, A.S.; Sidhu, G.P.S. Abiotic stress-induced oxidative stress in wheat. In Wheat Production in Changing Environments; Hasanuzzaman, M., Nahar, K., Hossain, M.A., Eds.; Springer Nature: Singapore, 2019; pp. 225–239. [Google Scholar]
- Chen, Z.; Chen, X.; Yan, H.; Li, W.; Li, Y.; Cai, R.; Xiang, Y. The lipoxygenase gene family in poplar: Identification, classification, and expression in response to MeJA treatment. PLoS ONE 2015, 10, e0125526. [Google Scholar] [CrossRef] [Green Version]
- De Simone, V.; Menzo, V.; De Leonardis, A.M.; Ficco, D.B.M.; Trono, D.; Cattivelli, L.; De Vita, P. Different mechanisms control lipoxygenase activity in durum wheat kernels. J. Cereal Sci. 2010, 52, 121–128. [Google Scholar] [CrossRef]
- Doderer, A.; Kokkelink, I.; Van der Ween, S.; Valk, B.I.; Schram, A.W.; Douma, A.C. Purification and characterization of two lipoxygenase isoenzymes from germinating barley. Biochim. Biophys. Acta 1992, 1120, 97–104. [Google Scholar] [CrossRef]
- Pokotylo, I.V.; Kolesnikov, Y.S.; Derevyanchuk, M.V.; Kharitonenko, A.I.; Kravets, V.S. Lipoxygenases and plant cell metabolism regulation. Ukr. Biochem. J. 2015, 87, 41–55. [Google Scholar] [CrossRef] [Green Version]
- De Simone, V.; Soccio, M.; Borrelli, G.M.; Pastore, D.; Trono, D. Stay-green trait-antioxidant status interrelationship in durum wheat (Triticum durum) flag leaf during post-flowering. J. Plant Res. 2014, 127, 159–171. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in ExPASy server. In Methods in Molecular Biology, 2-D Proteome Analysis Protocols; Link, A.J., Ed.; Humana Press: Totowa, NJ, USA, 1998; Volume 112, pp. 531–552. [Google Scholar]
- Sarde, S.J.; Kumar, A.; Remme, R.N.; Dicke, M. Genome-wide identification, classification and expression of lipoxygenase gene family in pepper. Plant Mol. Biol. 2018, 98, 375–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minor, W.; Steczko, J.; Stec, B.; Otwinowski, Z.; Bolin, J.T.; Walter, R.; Axelrod, B. Crystal structure of soybean lipoxygenase L-1 at 1.4 Å resolution. Biochemistry 1996, 35, 10687–10701. [Google Scholar] [CrossRef] [PubMed]
- Hugues, M.; Boivin, P.; Gauillard, F.; Nicolas, J.; Thiry, J.M.; Richard-Forget, F. Two lipoxygenases from germinated barley-heat and kilning stability. J. Food Sci. 1994, 59, 885–889. [Google Scholar] [CrossRef]
- Yang, G.; Schwarz, P.B.; Brady, A.V. Purification and characterization of lipoxygenase isoenzymes in germinating barley. Cereal Chem. 1993, 70, 589–595. [Google Scholar]
- Christensen, S.A.; Nemchenko, A.; Borrego, E.; Murray, I.; Sobhy, I.S.; Bosak, L.; DeBlasio, S.; Erb, M.; Robert, C.A.M.; Vaughn, K.A.; et al. The maize lipoxygenase, ZmLOX10, mediates green leaf volatile, jasmonate and herbivore-induced plant volatile production for defense against insect attack. Plant J. 2013, 74, 59–73. [Google Scholar] [CrossRef]
- Cofer, T.M.; Engelberth, M.; Engelberth, J. Green leaf volatiles protect maize (Zea mays) seedlings against damage from cold stress. Plant Cell Environ. 2018, 41, 1673–1682. [Google Scholar] [CrossRef]
- Tian, S.; Guo, R.; Zou, X.; Zhang, X.; Yu, X.; Zhan, Y.; Ci, D.; Wang, M.; Wang, Y.; Si, T. Priming with the green leaf volatile (Z)-3-hexeny-1-yl acetate enhances salinity stress tolerance in peanut (Arachis hypogaea L.) seedlings. Front. Plant Sci. 2019, 10, 785. [Google Scholar] [CrossRef]
- Sanz, L.C.; Peréz, A.G.; Olías, J.M. The lipoxygenase in the plant kingdom. I. Property. Grasas y Aceites 1992, 43, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Babenko, L.M.; Shcherbatiuk, M.M.; Skaterna, T.D.; Kosakivska, I.V. Lipoxygenases and their metabolites in formation of plant stress tolerance. Ukr. Biochem. J. 2017, 89, 5–21. [Google Scholar] [CrossRef]
- Cho, K.; Kim, Y.C.; Woo, J.C.; Rakwald, R.; Agrawal, G.K.; Yoeun, S.; Hana, O. Transgenic expression of dual positional maize lipoxygenase-1 leads to the regulation of defense-related signaling molecules and activation of the antioxidative enzyme system in rice. Plant Sci. 2012, 185–186, 238–245. [Google Scholar] [CrossRef]
- Morales, M.; Munné-Bosch, S. Malondialdehyde: Facts and artifacts. Plant Physiol. 2019, 180, 1246–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francois, L.E.; Maas, E.V. Crop response and management of salt-affected soils. In Handbook of Plant and Crop Stress; Pessarakli, M., Ed.; CRC Press: New York, NY, USA, 1994; pp. 149–181. [Google Scholar]
- Ellis, R.H.; Roberts, E.H. Towards a rational basis for testing seed quality. In Seed Production; Hebblethwaite, P.D., Ed.; Butterworths: London, UK, 1980; pp. 605–635. [Google Scholar]
- Zhang, H.Y.; Jiang, Y.N.; He, Z.Y.; Ma, M. Cadmium accumulation and oxidative burst in garlic (Allium sativum). J. Plant Physiol. 2005, 162, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar]
- Harris, D.A. Spectrophotometric assays. In Spectrophotometry and Spectrofluorimetry: A Practical Approach; Bashford, C.L., Harris, D.A., Eds.; IRL Press: Oxford, UK, 1987; pp. 59–61. [Google Scholar]
- Feng, B.; Dong, Z.; Xu, Z.; An, X.; Qin, H.; Wu, N.; Wang, D.; Wang, T. Molecular analysis of lipoxygenase (LOX) genes in common wheat and phylogenetic investigation of LOX proteins from model and crop plants. J. Cereal Sci. 2010, 52, 387–394. [Google Scholar] [CrossRef]
- Zhang, C.; Cao, S.; Jin, Y.; Ju, L.; Chen, Q.; Xing, Q.; Qi, H. Melon13-lipoxygenase CmLOX18 may be involved in C6 volatiles biosynthesis in fruit. Sci. Rep. 2017, 7, 2816. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Km (µM) | Vmax (E.U./mg of Protein) |
|---|---|---|
| Linoleic acid | 73.69 ± 7.33 | 35.42 ± 3.16 |
| Linolenic acid | 300.10 ± 21.36 | 7.41 ± 0.95 |
| Arachidonic acid | 741.00 ± 58.35 | 2.45 ± 0.51 |
| Gene Fragment | Forward Primer (5′→3′) | Reverse Primer (5′→3′) | Annealing Temperature (°C) | Product Size (bp) |
|---|---|---|---|---|
| TdLpx-B1.1 | CCAAGATGATACTGGGCGGGC | CGCCGCCTTGCCGTGGTTGG | 62 | 1154 |
| TdLpx-B1.2 | TACACGCCGGTGCCGAGCGGCAG | CGTGTCACGCTGCCCGAGGTAGAG | 72 | 912 |
| TdLpx-A2 | GACCTGACCACGGCGAAGAGCACC | TGCGGGCTCGATGGGTCCTCCACC | 70 | 425 |
| TdLpx-B2 | GACCTGACCACCGCGAAGAGCACG | TTGTACACCGGGTGGGTCACGCTC | 68 | 206 |
| TdLpx-A3 | AATACAGCACGGCGCGAAGAGCAC | GTTCACCCACCGCTCGATCGCCC | 70 | 514 |
| TdLpx-B3 | CCTGCCGCACCCCCAGGGGATG | GTTCACCCACCGCTCGATCGCCC | 70 | 532 |
| Actin1 | CTTCGGACCCAAGAAAGAAAGCC | CACCGCCCGTATTTCTCTAGTAGCC | 62 | 280 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menga, V.; Trono, D. The Molecular and Functional Characterization of the Durum Wheat Lipoxygenase TdLOX2 Suggests Its Role in Hyperosmotic Stress Response. Plants 2020, 9, 1233. https://doi.org/10.3390/plants9091233
Menga V, Trono D. The Molecular and Functional Characterization of the Durum Wheat Lipoxygenase TdLOX2 Suggests Its Role in Hyperosmotic Stress Response. Plants. 2020; 9(9):1233. https://doi.org/10.3390/plants9091233
Chicago/Turabian StyleMenga, Valeria, and Daniela Trono. 2020. "The Molecular and Functional Characterization of the Durum Wheat Lipoxygenase TdLOX2 Suggests Its Role in Hyperosmotic Stress Response" Plants 9, no. 9: 1233. https://doi.org/10.3390/plants9091233
APA StyleMenga, V., & Trono, D. (2020). The Molecular and Functional Characterization of the Durum Wheat Lipoxygenase TdLOX2 Suggests Its Role in Hyperosmotic Stress Response. Plants, 9(9), 1233. https://doi.org/10.3390/plants9091233