
Modeling Study of Factors Determining Efficacy of Biological Control of Adventive Weeds



Abstract
:1. Introduction
2. The Model
3. Results
3.1. Homogeneous Case
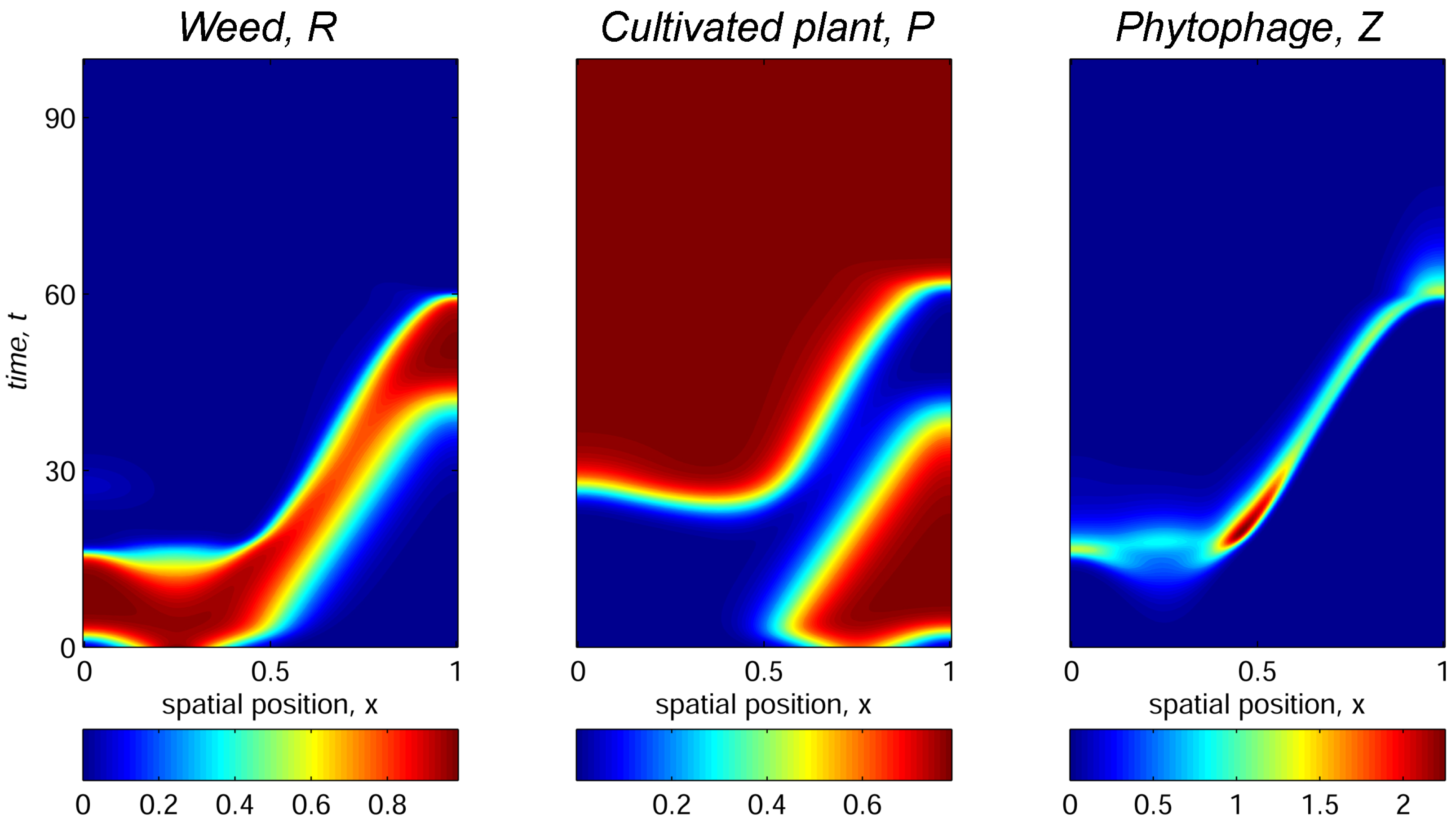
3.2. Case of 1D Spatial Domain
3.3. Case of 2D Spatial Domain
4. Discussion and Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| PDE | Partial differential equation |
| ODE | Ordinary differential equation |
| SPW | Solitary population wave |
References
- McFadyen, R.E.C. Successes in biological control of weeds. In Proceedings of the X International Symposium on Biological Control of Weeds, Bozeman, MT, USA, 4–14 July 1999; Spencer, N.R., Ed.; Montana State University: Bozeman, MT, USA, 2000; pp. 3–14. Available online: https://www.invasive.org/publications/xsymposium/proceed/01apg03.pdf (accessed on 29 November 2023).
- Cuda, J.P.; Charudattan, R.; Grodowitz, M.J.; Newman, R.M.; Shearer, J.F.; Tamayo, M.L.; Vilegas, B. Recent advances in biological control of submersed aquatic weeds. J. Aquat. Plant Manag. 2008, 46, 15–32. [Google Scholar]
- Kovalev, O.V.; Tyutyunov, Y.V. The role of solitary population waves in efficient suppression of adventive weeds by introduced phytophagous insects. Entmol. Rev. 2014, 94, 310–319. [Google Scholar] [CrossRef]
- Mason, P. (Ed.) Biological Control: Global Impacts, Challenges and Future Directions of Pest Management; CSIRO Publishing: Canberra, Australia, 2021; 626p. [Google Scholar]
- López-Núñez, F.A.; Marchante, E.; Heleno, R.; Duarte, L.N.; Palhas, J.; Impson, F.; Freitas, H.; Marchante, H. Establishment, spread and early impacts of the first biocontrol agent against an invasive plant in continental Europe. J. Environ. Manag. 2021, 290, 112545. [Google Scholar] [CrossRef] [PubMed]
- Shaw, R.H.; Ellison, C.A.; Marchante, H.; Pratt, C.F.; Schaffner, U.; Sforza, R.F.H.; Deltoro, V. Weed biological control in the European Union: From serendipity to strategy. BioControl 2018, 63, 333–347. [Google Scholar] [CrossRef]
- Luck, R.F. Evaluation of natural enemies for biological control: A behavioral approach. Trends Ecol. Evol. 1990, 5, 196–199. [Google Scholar] [CrossRef] [PubMed]
- Arditi, R.; Berryman, A.A. The biological control paradox. Trends Ecol. Evol. 1991, 6, 32. [Google Scholar] [CrossRef] [PubMed]
- Berryman, A. The theoretical foundations of biological control. In Theoretical Approaches to Biological Control; Hawkins, B., Cornell, H.V., Eds.; Cambridge University Press: Cambridge, UK, 1999; pp. 3–21. [Google Scholar]
- Sapoukhina, N.; Tyutyunov, Y.; Arditi, R. The role of prey-taxis in biological control: A spatial theoretical model. Am. Nat. 2003, 162, 61–76. [Google Scholar] [CrossRef]
- Kovalev, O.V. The solitary population wave, a physical phenomenon accompanying the introduction of a chrysomelid. In New Developments in the Biology of Chrysomelidae; Jolivet, P., Ed.; SPB Academic Publishing BV: The Hague, The Netherlands, 2004; pp. 591–601. [Google Scholar] [CrossRef]
- Kovalev, O.V. Spread of adventive plants of Ambrosieae tribe in Eurasia and methods of bilogical control of Ambrosia L. (Asteraceae). In Theoretical Principles of Biological Control of the Common Ragweed, Proceedings of the Zoological Institute; Kovalev, O.V., Belokobylsky, S.A., Eds.; “Nauka” Publishing House, Leningrad Branch: Leningrad, Russia, 1989; Volume 189, pp. 7–23. (In Russian) [Google Scholar]
- Kovalev, O.V.; Tyutyunov, Y.V.; Iljina, L.P.; Berdnikov, S.V. On the efficiency of introduction of american insects feeding on the common ragweed (Ambrosia artemisiifolia L.) in the South of Russia. Entomol. Rev. 2013, 92, 251–264. [Google Scholar] [CrossRef]
- Vechernin, V.V.; Kovalev, O.V. New mathematical model of a solitary wave for describing a population wave. Biophysics 1988, 33, 701–707. [Google Scholar]
- Tyutyunov, Y.V.; Kovalev, O.V.; Titova, L.I. Spatial demogenetic model for studying phenomena observed upon introduction of the ragweed leaf beetle in the south of Russia. Math. Model. Nat. Phenom. 2013, 8, 80–95. [Google Scholar] [CrossRef]
- Tyutyunov, Y.V. Spatial demo-genetic predator–prey model for studying natural selection of traits enhancing consumer motility. Mathematics 2023, 11, 3378. [Google Scholar] [CrossRef]
- Kovalev, O.V.; Tyutyunov, Y.V.; Arkhipova, O.E.; Kachalina, N.A.; Iljina, L.P.; Titova, L.I. On assessment of the large-scale effect of introduction of the ragweed leaf beetle Zygogramma suturalis F. (Coleoptera, Chrysomelidae) Phytocenoses South Russia. Entomol. Rev. 2015, 95, 1–14. [Google Scholar] [CrossRef]
- Allee, W. Animal Aggregations: A Study in General Sociology; Chicago University Press: Chicago, IL, USA, 1931; 431p. [Google Scholar]
- Dennis, B. Allee effects: Population growth, critical density, and the chance of extinction. Nat Resour Model. 1989, 3, 481–538. [Google Scholar] [CrossRef]
- Bazykin, A.D. Nonlinear Dynamics of Interacting Populations; World Scientific Publishing: Singapore, 1998; 193p. [Google Scholar]
- Courchamp, F.; Berec, J.; Gascoigne, J. Allee Effects in Ecology and Conservation; Oxford University Press: Oxford, NY, USA, 2008; 256p. [Google Scholar]
- Tyutyunov, Y.; Sen, D.; Titova, L.I.; Banerjee, M. Predator overcomes the Allee effect due to indirect prey-taxis. Ecol. Complex. 2019, 39, 100772. [Google Scholar] [CrossRef]
- Hairer, E.; Nørsett, S.; Wanner, G. Solving Ordinary Differential Equations I. Nonstiff Problems; Springer: Berlin/Heidelberg, Germany, 2009; 528p. [Google Scholar]
- Budyansky, A.V.; Frischmuth, K.; Tsybulin, V.G. Cosymmetry approach and mathematical modeling of species coexistence in a heterogeneous habitat. Discrete Contin. Dyn. Syst.-Ser. B 2019, 24, 547–561. [Google Scholar] [CrossRef]
- Govorukhin, V.N.; Zagrebneva, A.D. Population waves and their bifurcations in a model “active predator–passive prey”. Comput. Res. Model. 2020, 12, 831–843. (In Russian) [Google Scholar] [CrossRef]
- Gause, G.F.; Witt, A.A. Behavior of mixed populations and the problem of natural selection. Am. Nat. 1935, 69, 596–609. [Google Scholar] [CrossRef]
- Mallet, J. The struggle for existence: How the notion of carrying capacity, K, obscures the links between demography, Darwinian evolution, and speciation. Evol. Ecol. Res. 2012, 14, 627–665. [Google Scholar]
- Myers, J.H. How many insect species are necessary for successful biocontrol of weeds? In Proceedings of the VI International Symposium on Biological Control of Weeds, Vancouver, BC, Canada, 19–25 August 1984; Delfosse, E.S., Ed.; Agriculture: Vancouver, BC, Canada, 1985; pp. 77–82. [Google Scholar]
- McEvoy, P.B.; Coombs, E.M. Biological control of plant invaders: Regional patterns, field experiments, and structured population models. Ecol. Appl. 1999, 9, 387–401. [Google Scholar] [CrossRef]
- Briese, D.T. Classical biological control. In Australian Weed Management Systems; Sindel, B.M., Ed.; R.G. & F.J. Richardson: Melbourne, Australia, 2000; pp. 161–192. [Google Scholar]
- Van Klinken, R.D.; Raghu, S. A scientific approach to agent selection. Aust. J. Entomol. 2006, 45, 253–258. [Google Scholar] [CrossRef]
- Müller-Schärer, H.; Schäffner, U. Classical biological control: Exploiting enemy escape to manage plant invasions. Biol. Invasions 2006, 10, 859–874. [Google Scholar] [CrossRef]
- Schwarzländer, M.; Hinz, H.L.; Winston, R.L.; Day, M.D. Biological control of weeds: An analysis of introductions, rates of establishment and estimates of success, worldwide. BioControl 2018, 63, 319–331. [Google Scholar] [CrossRef]
- Julien, M.N.; Griffiths, M.W. Biological Control of Weeds: A World Catalogue of Agents and Their Target Weeds, 4th ed.; CABI Publishing: Wallingford, UK, 1998; 223p. [Google Scholar]
- Kovalev, O.V.; Medvedev, L.N. Theoretical basis of introduction of ragweed leaf beetles of the genus Zygogramma Chevr. (Coleoptera: Chrysomelidae) in the USSR for biological control of ragweed. Entomol. Obozr. 1983, 62, 17–32. (In Russian) [Google Scholar]
- Kovalev, O.V. New factor of efficiency of phytophages: A solitary population wave and succession process. In Proceedings of the VII International Symposium on Biological Control of Weeds, Rome, Italy, 6–11 March 1988; Delfosse, E.S., Ed.; Ministero dell’Agricoltura e delle Foreste: Rome, Italy; CSIRO: Melbourne, Australia, 1990; pp. 51–53. [Google Scholar]
- Arditi, R.; Tyutyunov, Y.; Morgulis, A.; Govorukhin, V.; Senina, I. Directed movement of predators and the emergence of density-dependence in predator-prey models. Theor. Popul. Biol. 2001, 59, 207–221. [Google Scholar] [CrossRef]
- Ginzburg, L.R.; D’Andrea, R. Trophic Levels. In Encyclopedia of Biodiversity, 3rd ed.; Scheiner, S.M., Ed.; Academic Press: Oxford, UK, 2024; Volume 5, pp. 252–258. [Google Scholar] [CrossRef]
- White, T.C.R. The Inadequate Environment: Nitrogen and the Abundance of Animals; Springer: Berlin, Germany, 1993; 425p. [Google Scholar]
- White, T.C.R. Why Does the World Stay Green?: Nutrition and Survival of Plant-Eaters; CSIRO Publishing: Collingwood, Australia, 2005; 120p. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Meaning | Set 1 | Set 2 | Set 3 | Set 4 [Dimension Units] * |
|---|---|---|---|---|---|
| Growth rate of weed | 1 | 1 | 1 | 0.0117 | |
| Growth rate of cultivated plant | 0.8 | 0.8 | 0.5 | 0.00337 | |
| Intraspecific competition coefficient of weed | 1 | 1 | 1 | 0.01 | |
| Intraspecific competition coefficient of cultivated plant | 1 | 1 | 1 | 0.045 | |
| Interspecific competition coefficient representing the effect of cultivated plant on weed | 1.5 | 1.5 | 1.5 | 0.07 | |
| Interspecific competition coefficient representing the effect of weed on cultivated plant | 1.5 | 1.5 | 1.5 | 0.005 | |
| a | Searching efficiency of phytophage | 3 | 3 | 3 | |
| h | Handling time of phytophage | 1 | 1 | 1/15 | 35,700 |
| Conversion efficiency of phytophage | 0.8 | 0.8 | 0.9 | 2625 | |
| Mortality coefficient of phytophage | 0.2 | 0.2 | 0.2 | 0.0114 | |
| Allee coefficient | 0.001 | 0.001 | 0.001 | 0.001 | |
| Diffusion coefficient of weed | – | 0.001 | 0.001 | 0.5 | |
| Diffusion coefficient of cultivated plant | – | 0.001 | 0.001 | 0.2 | |
| Diffusion coefficient of phytophage | – | 0.05 | 0.02 | 15 | |
| Diffusion coefficient of taxis stimulus | – | 0 | 0 | 0.05 | |
| Emission rate of taxis stimulus | – | 1 | 1 | 1 | |
| Trophotaxis coefficient | – | 0…0.17 | 0…0.7 | 0.01 | |
| Decay coefficients of stimulus | – | 0.001 | 0.001 | 0.01 | |
| Length of spatial domain | – | 1 | 1 | 4000 [m] | |
| Width of spatial domain | – | – | – | 3000 [m] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tyutyunov, Y.V.; Govorukhin, V.N.; Tsybulin, V.G. Modeling Study of Factors Determining Efficacy of Biological Control of Adventive Weeds. Mathematics 2024, 12, 160. https://doi.org/10.3390/math12010160
Tyutyunov YV, Govorukhin VN, Tsybulin VG. Modeling Study of Factors Determining Efficacy of Biological Control of Adventive Weeds. Mathematics. 2024; 12(1):160. https://doi.org/10.3390/math12010160
Chicago/Turabian StyleTyutyunov, Yuri V., Vasily N. Govorukhin, and Vyacheslav G. Tsybulin. 2024. "Modeling Study of Factors Determining Efficacy of Biological Control of Adventive Weeds" Mathematics 12, no. 1: 160. https://doi.org/10.3390/math12010160
APA StyleTyutyunov, Y. V., Govorukhin, V. N., & Tsybulin, V. G. (2024). Modeling Study of Factors Determining Efficacy of Biological Control of Adventive Weeds. Mathematics, 12(1), 160. https://doi.org/10.3390/math12010160