Analysis of the Incidence of Poxvirus on the Dynamics between Red and Grey Squirrels

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Model
2.2. Model Reparametrization
2.3. Study of the Ecosystem Behavior
3. Results
3.1. Equilibria
3.1.1. Feasibility
3.1.2. Stability
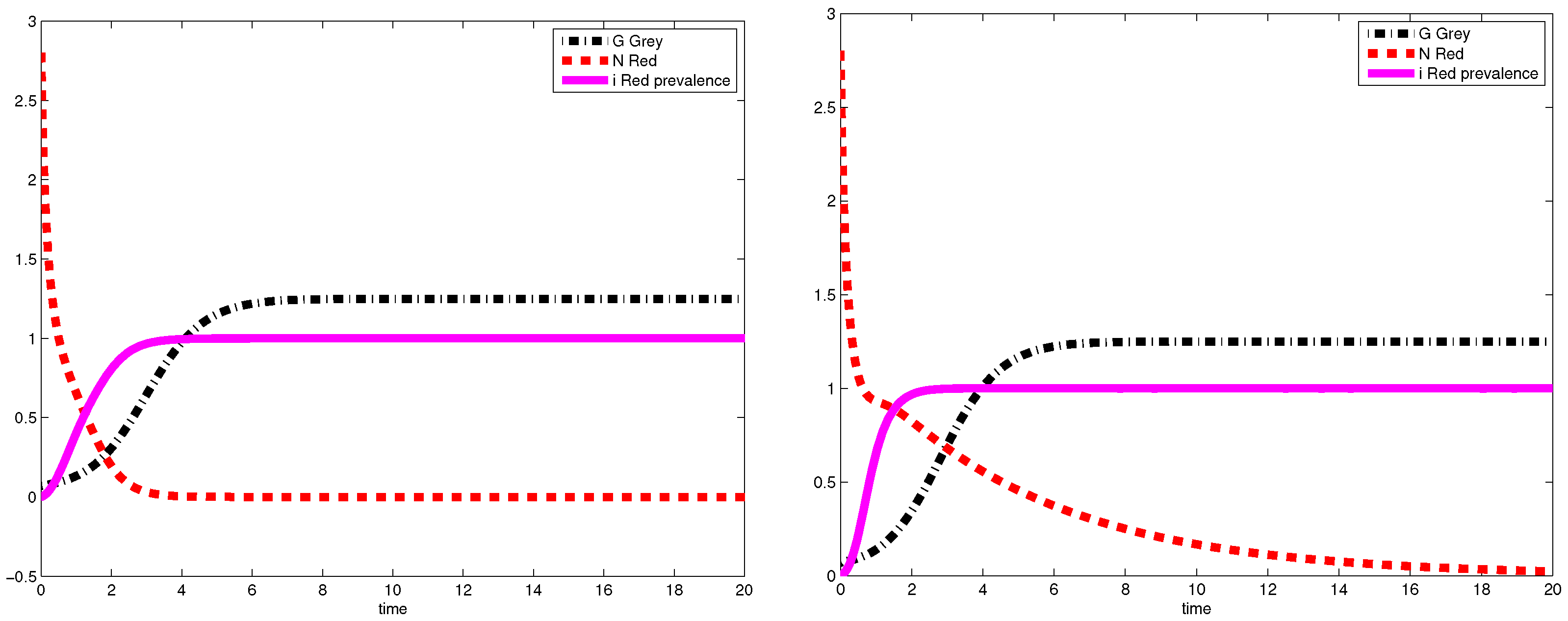
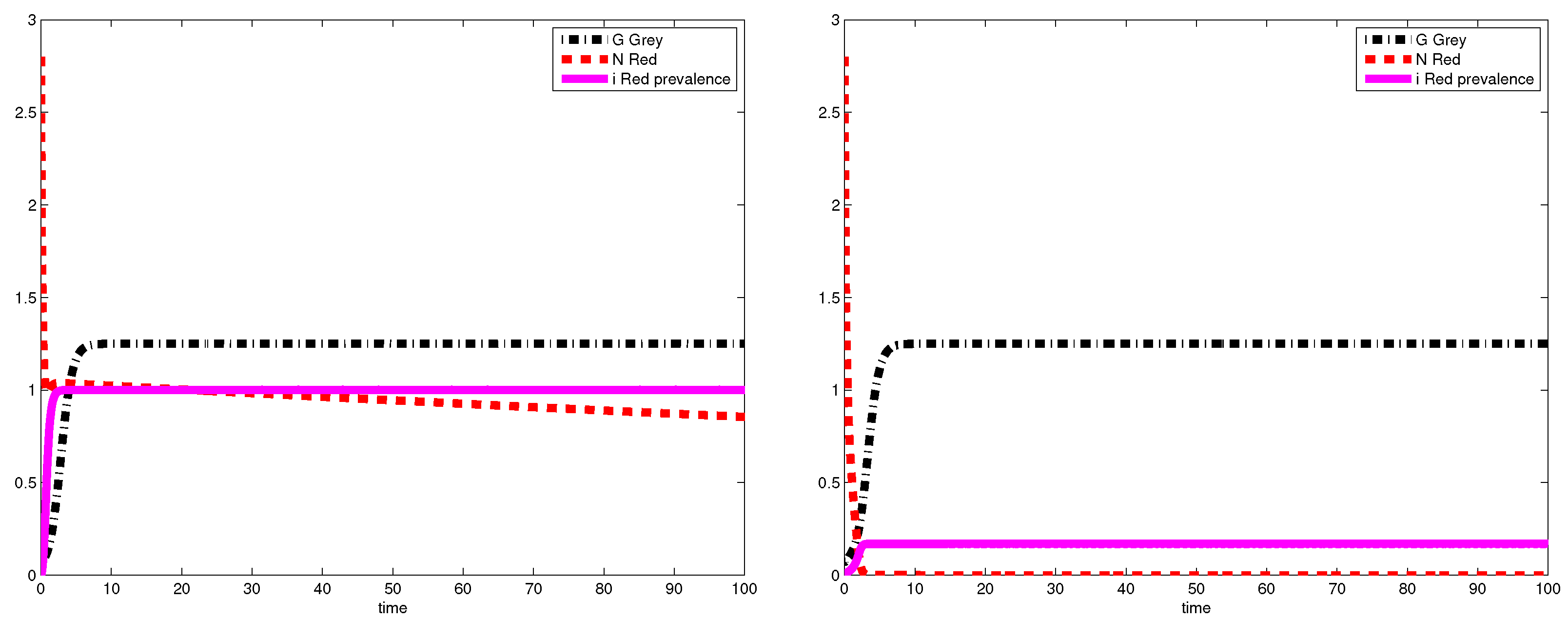
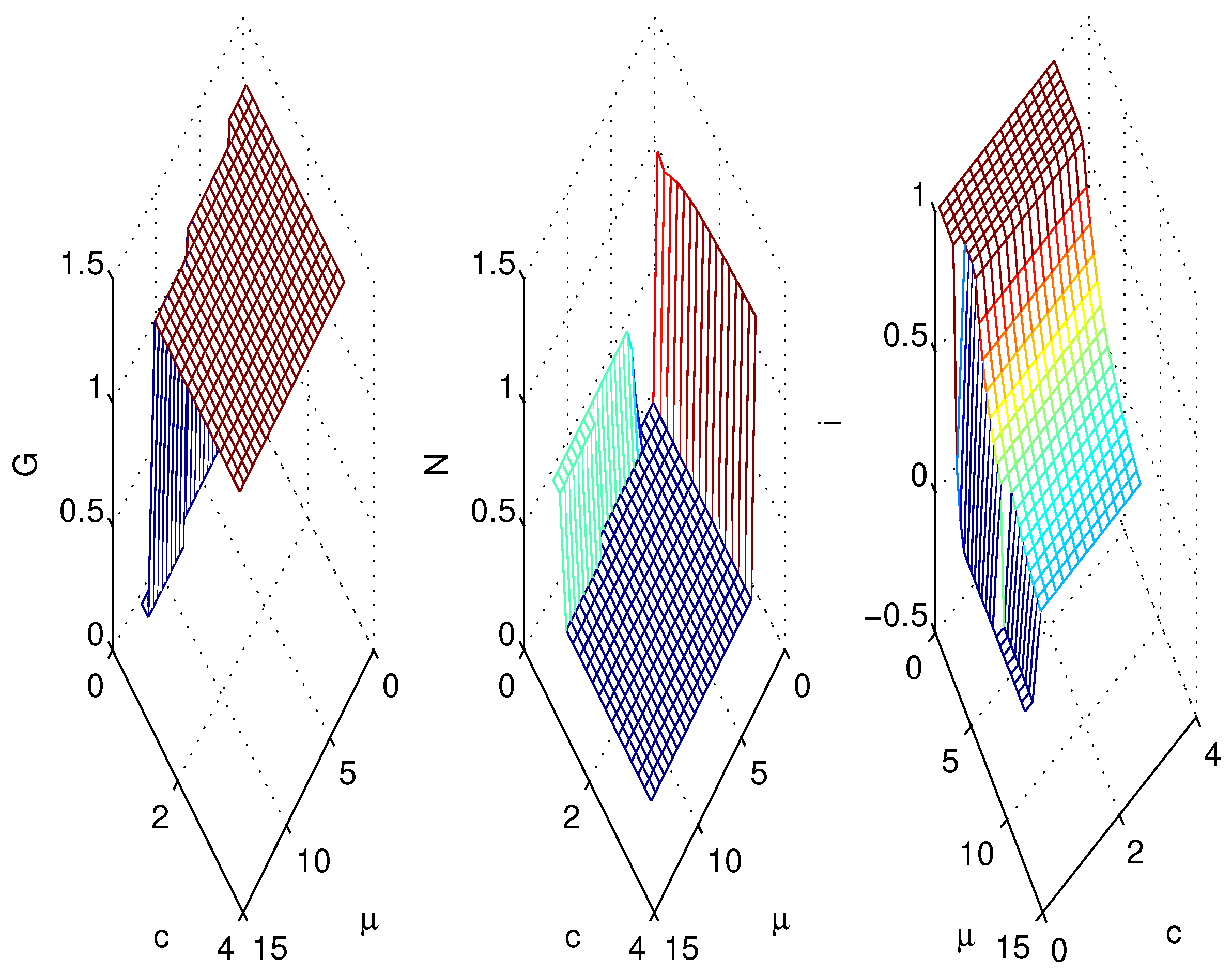
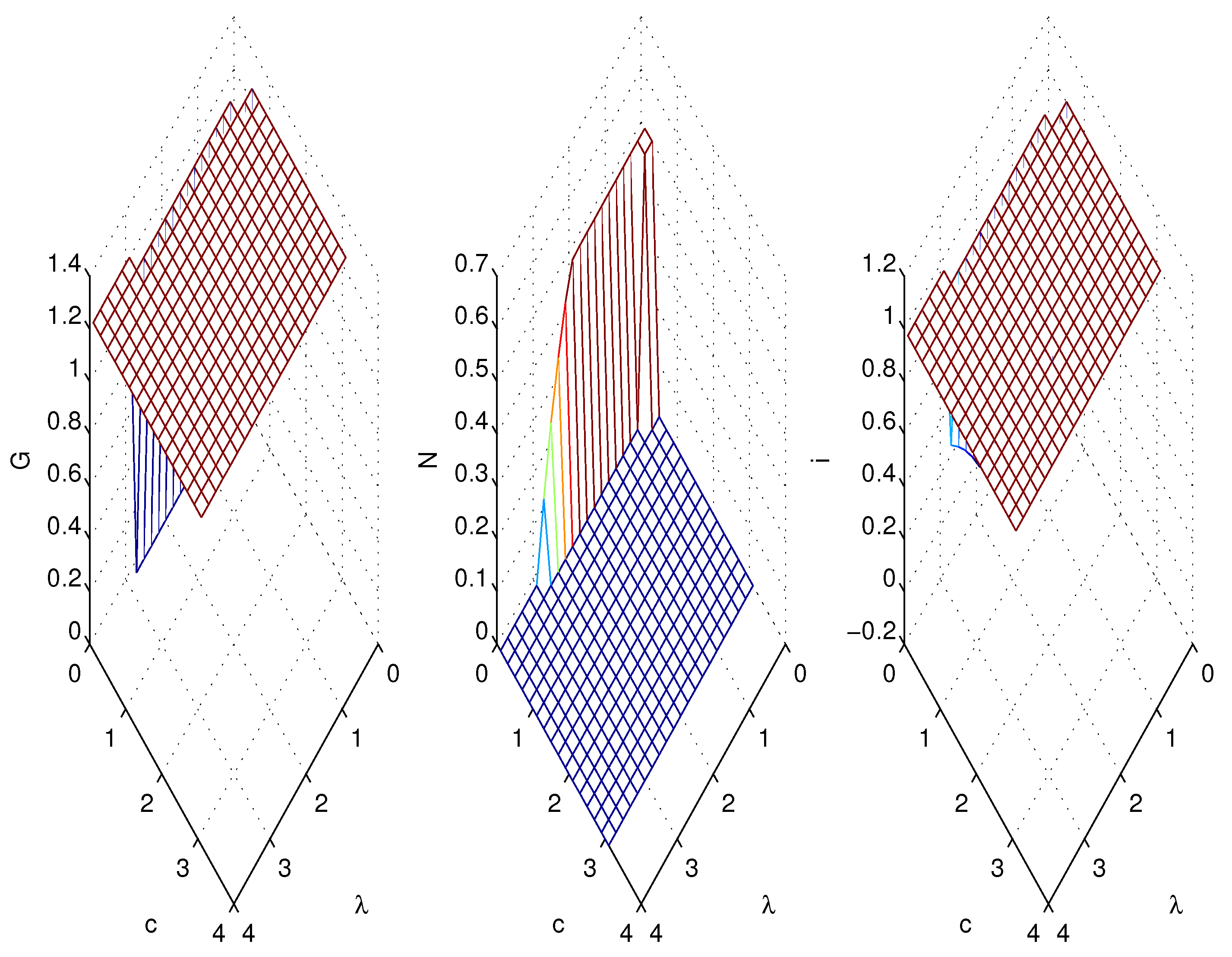
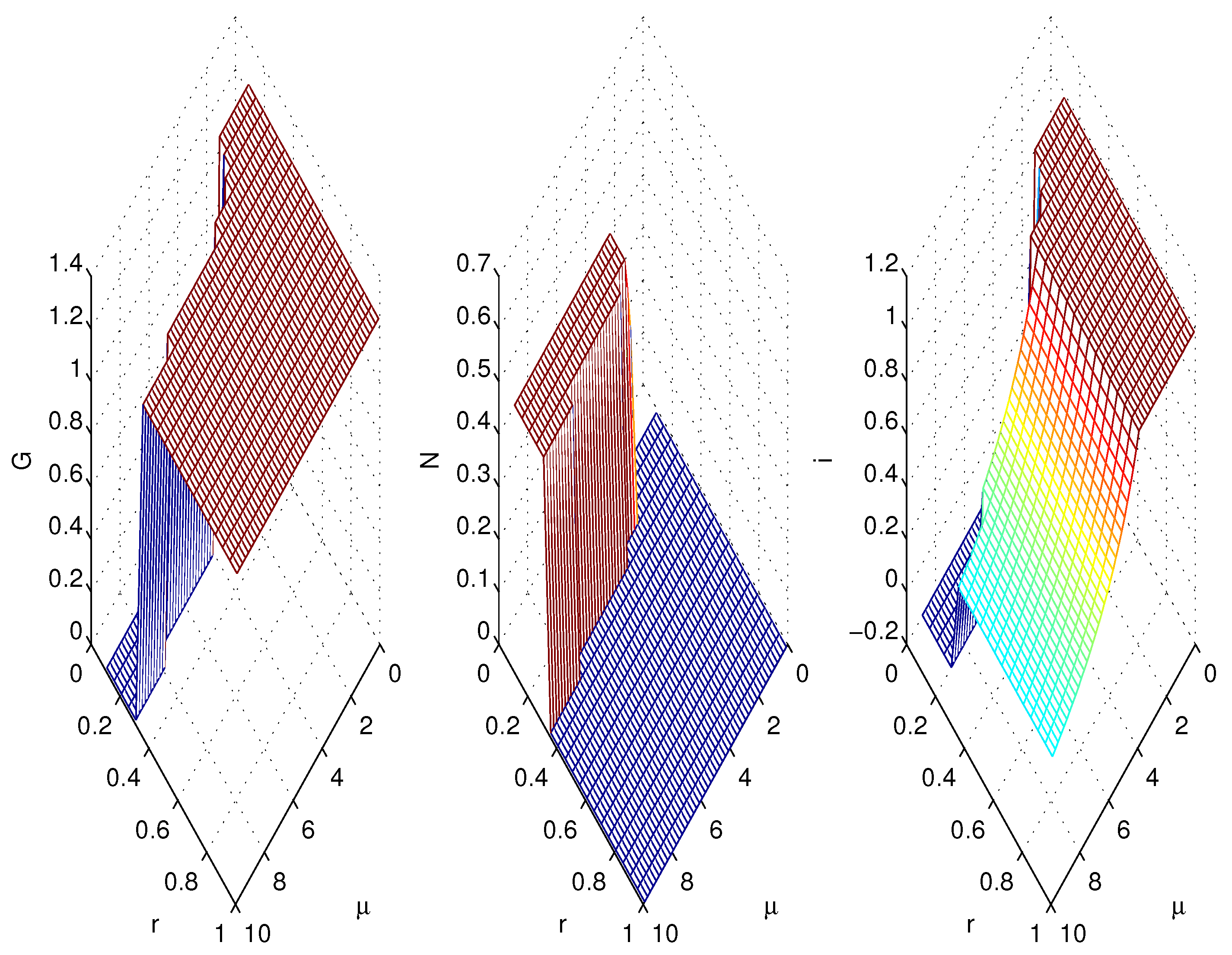
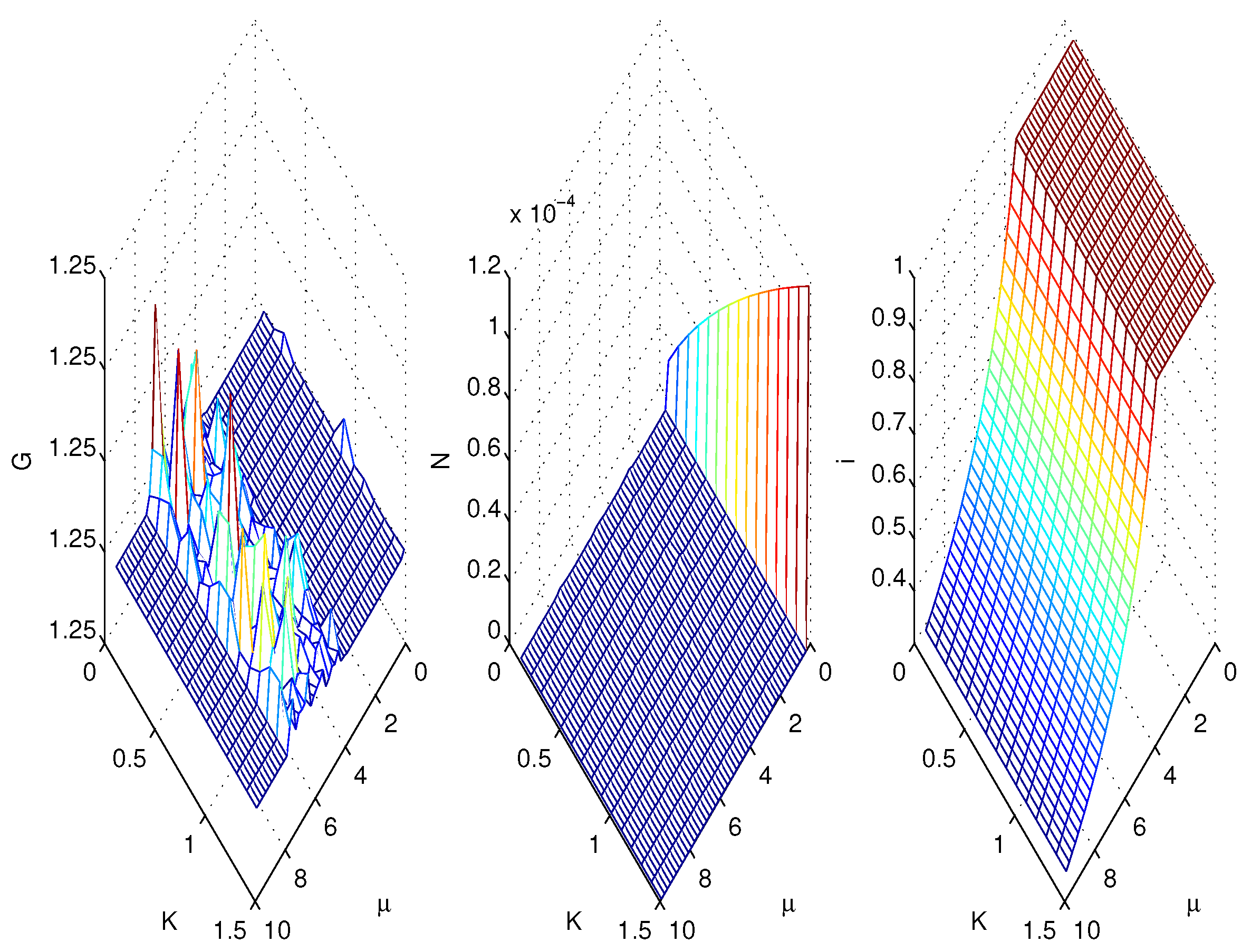
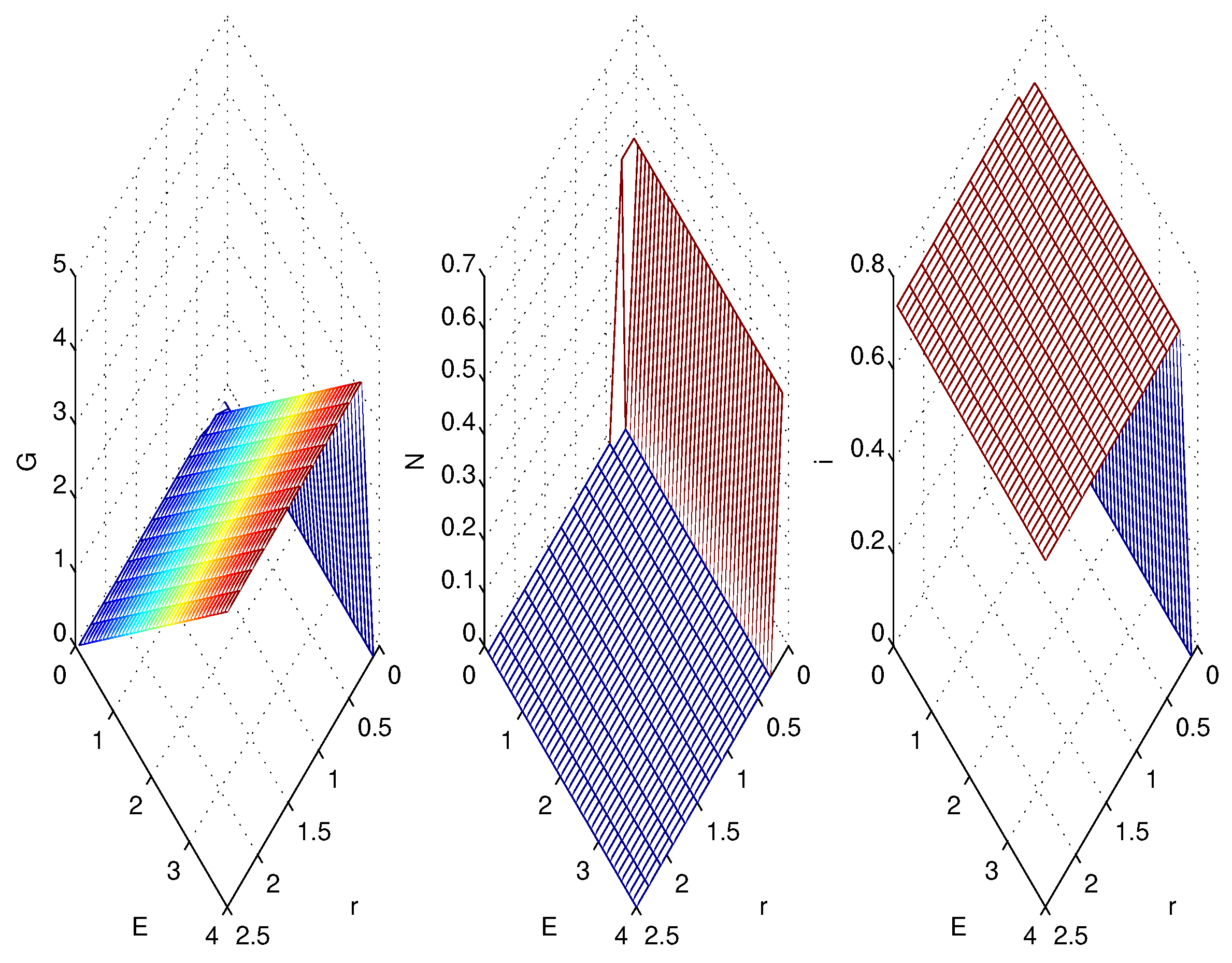
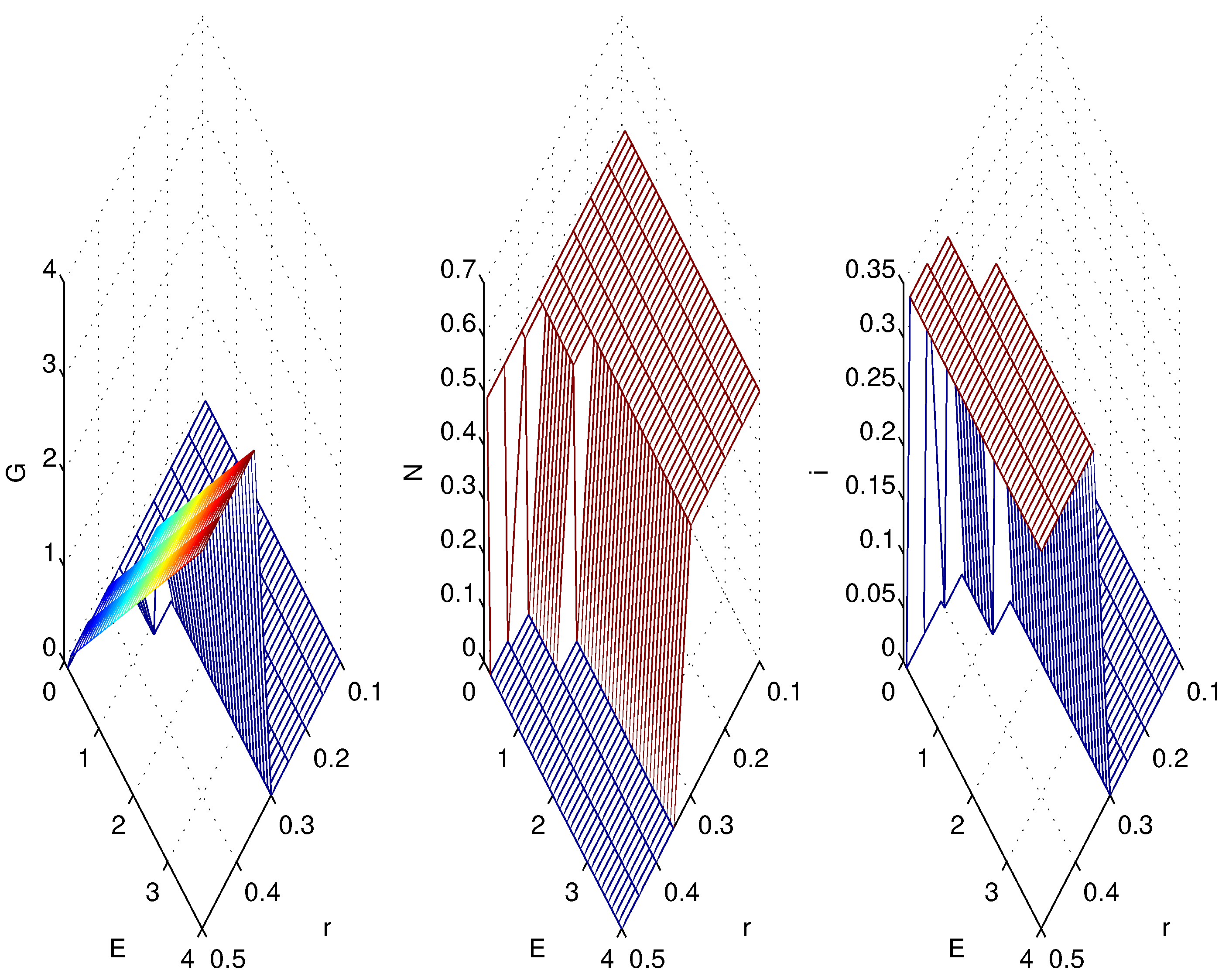
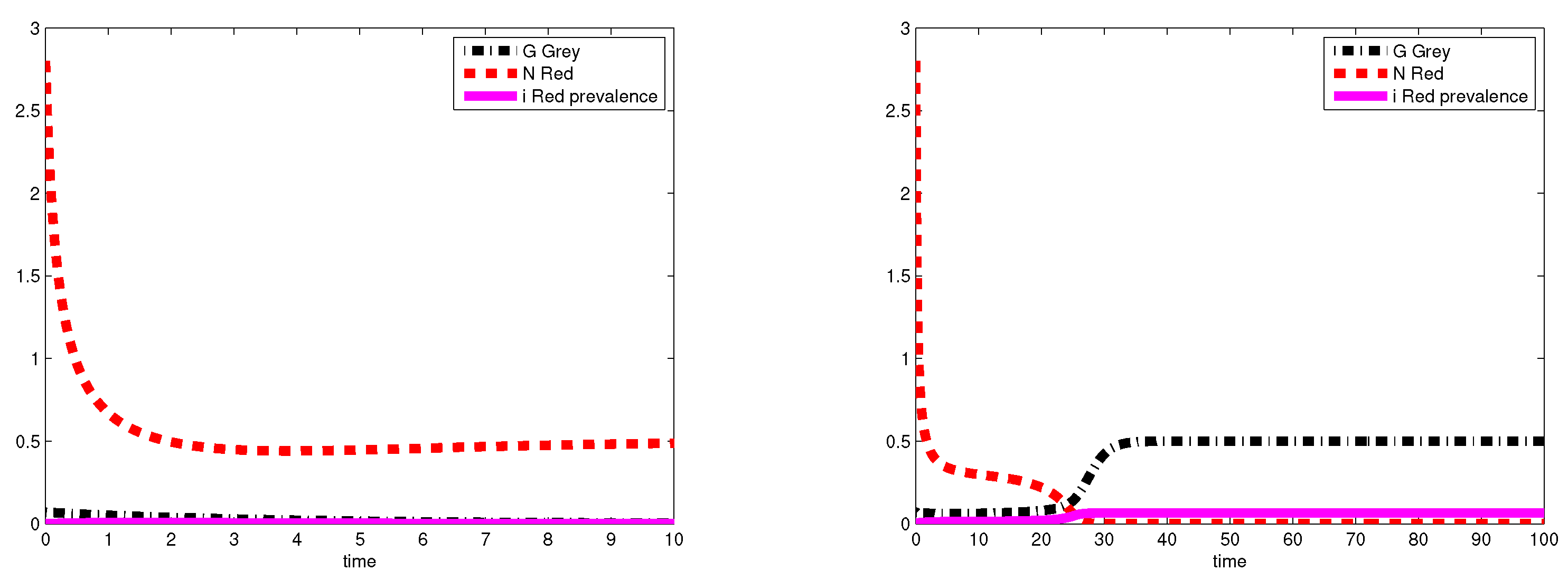
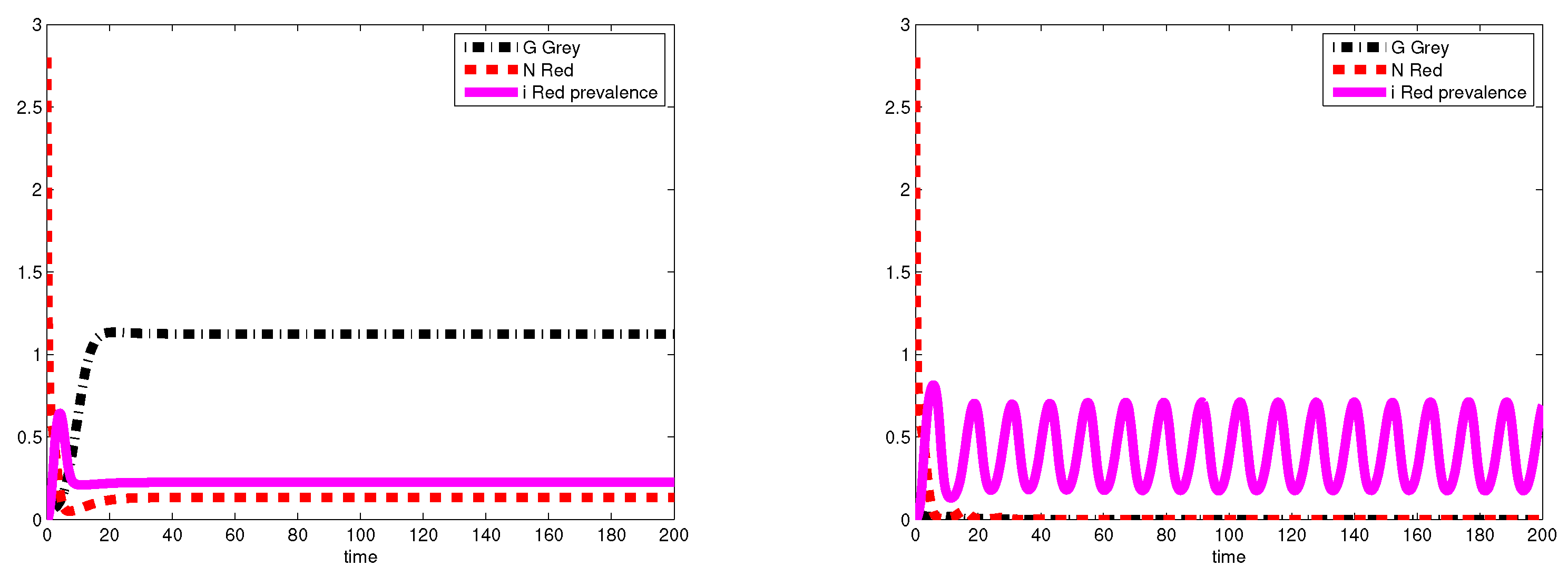
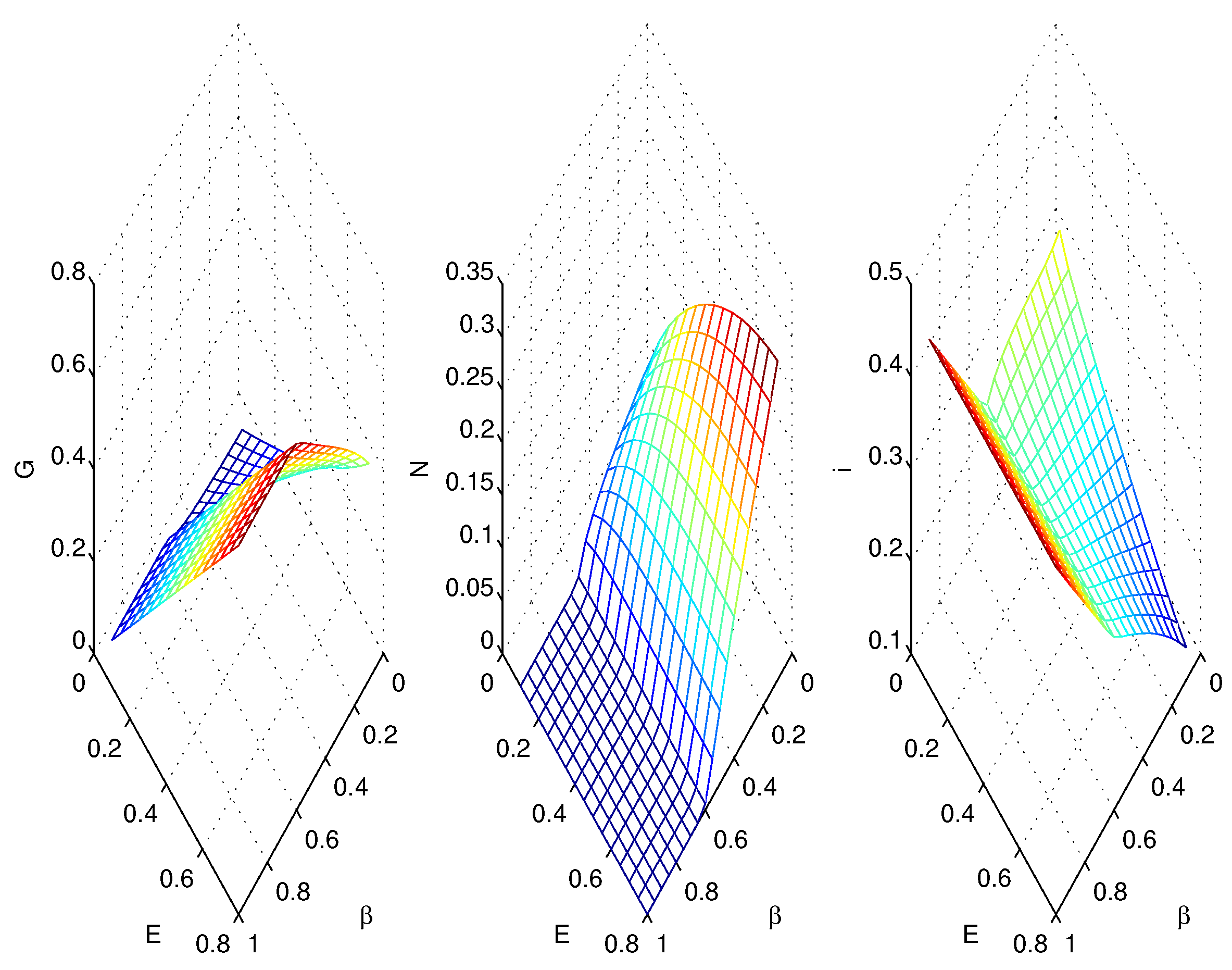
3.2. Simulations
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Shar, S.; Lkhagvasuren, D.; Bertolino, S.; Henttonen, H.; Kryštufek, B.; Meinig, H. Sciurus vulgaris (errata version published in 2017). The IUCN Red List of Threatened Species. 2017. Available online: http://www.iucnredlist.org/details/20025/0 (accessed on 27 June 2018).
- Middleton, A.D. 38. The ecology of the American grey squirrel (Sciurus carolinensis Gmelin) in the British Isles. Proc. Zool. Soc. Lond. 1930, 100, 809–843. [Google Scholar] [CrossRef]
- Shorten, M.; Elton, C. Some aspects of the biology of the grey squirrel (Sciurus carolinensis) in Great Britain. Proc. Zool. Soc. Lond. 1951, 121, 427–459. [Google Scholar] [CrossRef]
- Martinoli, A.; Bertolino, S.; Preatoni, D.G.; Balduzzi, A.; Marsan, A.; Genovesi, P.; Tosi, G.; Wauters, L.A. Headcount 2010: The multiplication of the grey squirrel populations introduced to Italy. Hystrix Ital. J. Mammal. 2010, 21, 127–136. [Google Scholar]
- Bertolino, S.; Martinoli, A.; Wauters, L.A. Risk Assessment for Sciurus carolinensis (grey squirrel). In Invasive Alien Species—Framework for the Identification of Invasive Species of EU Concern; Roy, H., Ed.; Natural Environment Research Council: Swindon, UK, 2014; p. 298. [Google Scholar]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species; The Invasive Species Specialist Group (ISSG), a Specialist Group of the Species Survival Commission (SSC) of the World Consevation Union (IUCN): Gland, Switzerland, 2000. [Google Scholar]
- Gurnell, J. Squirrel numbers and the abundance of tree seeds. Mamm. Rev. 1983, 13, 133–148. [Google Scholar] [CrossRef]
- Wauters, L.A.; Gurnell, J. The mechanism of replacement of red squirrels by grey squirrels: A test of the interference competition hypothesis. Ethology 1999, 29, 1053–1071. [Google Scholar] [CrossRef]
- Wauters, L.A.; Gurnell, J.; Martinoli, A.; Tosi, G. Interspecific competition between native Eurasian red squirrels and alien grey squirrels: Does resource partitioning occur? Behav. Ecol. Sociobiol. 2002, 52, 332–341. [Google Scholar] [CrossRef]
- Wauters, L.; Tosi, G.; Gurnell, J. Interspecific competition in tree squirrels: Do introduced grey squirrels (Sciurus carolinensis) deplete tree seeds hoarded by red squirrels (S. vulgaris)? Behav. Ecol. Sociobiol. 2002, 51, 360–367. [Google Scholar] [CrossRef]
- Wauters, L.A.; Lurz, P.W.W.; Gurnell, J. Interspecific effects of grey squirrels (Sciurus carolinensis) on the space use and population demography of red squirrels (Sciurus vulgaris) in conifer plantations. Ecol. Res. 2000, 15, 271–284. [Google Scholar] [CrossRef]
- Thomas, K.; Tompkins, D.M.; Sainsbury, A.W.; Wood, A.R.; Dalziel, R.; Nettleton, P.F.; McInnes, C.J. A novel poxvirus lethal to red squirrels (Sciurus vulgaris). J. Gener. Virol. 2003, 84, 3337–3341. [Google Scholar] [CrossRef] [PubMed]
- McInnes, C.J.; Wood, A.R.; Thomas, K.; Sainsbury, A.W.; Gurnell, J.; Dein, F.J.; Nettleton, P.F. Genomic characterization of a novel poxvirus contributing to the decline of the red squirrel (Sciurus vulgaris) in the UK. J. Gener. Virol. 2006, 87, 2115–2125. [Google Scholar] [CrossRef] [PubMed]
- Sainsbury, A.W.; Nettleton, P.; Gurnell, J. Recent developments in the study of parapoxvirus in red and grey squirrels. In The Conservation of Red Squirrels, Sciurus vulgaris L.; Gurnell, J., Lurz, P.W.W., Eds.; PTES (People’s Trust for Endangered Species): London, UK, 1997; pp. 105–108. [Google Scholar]
- Edwards, F.B. Red squirrel disease. Vet. Rec. 1962, 74, 739–741. [Google Scholar]
- Sainsbury, A.; Gurnell, J. An investigation into the health and welfare of red squirrels, Sciurus vulgaris, involved in reintroduction studies. Vet. Rec. 1995, 137, 367–370. [Google Scholar] [CrossRef] [PubMed]
- McInnes, C.J.; Coulter, L.; Dagleish, M.P.; Fiegna, C.; Gilray, J.; Willoughby, K.; Cole, M.; Milne, E.; Meredith, A.; Everest, D.J.; et al. First cases of squirrelpox in red squirrels (Sciurus vulgaris) in Scotland. Vet. Rec. 2009, 164, 528–531. [Google Scholar] [CrossRef] [PubMed]
- Tompkins, D.M.; Sainsbury, A.W.; Nettleton, P.; Buxton, D.; Gurnell, J. Parapoxvirus causes a deleterious disease in red squirrels associated with UK population declines. Proc. R. Soc. B Biol. Sci. 2002, 269, 529–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sainsbury, A.W.; Nettleton, P.; Gilray, J.; Gurnell, J. Grey squirrels have high seroprevalence to a parapoxvirus associated with deaths in red squirrels. Anim. Conserv. 2000, 3, 229–233. [Google Scholar] [CrossRef]
- Sainsbury, A.W.; Deaville, R.; Lawson, B.; Cooley, W.A.; Farelly, S.S.J.; Stack, M.J.; Duff, P.; McInnes, C.J.; Gurnell, J.; Russell, P.H.; et al. Poxviral disease in red squirrels Sciurus vulgaris in the UK: Spatial and temporal trends of an emerging threat. EcoHealth 2008, 5, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Sainsbury, T.; Ward, L. Parapoxvirus infection in red squirrels. Vet. Rec. 1996, 138, 400. [Google Scholar] [PubMed]
- Scott, A.; Keymer, I.; Labram, J. Parapoxvirus infection of the red squirrel (Sciurus vulgaris). Vet. Rec. 1981, 109, 202. [Google Scholar] [CrossRef] [PubMed]
- Duff, J.P.; Scott, A.; Keymer, I.F. Parapoxvirus infection of the grey squirrel. Vet. Rec. 1996, 138, 527. [Google Scholar] [PubMed]
- Reynolds, J.C. Details of the geographic replacement of the red squirrel (Sciurus vulgaris) by the grey squirrel (Sciurus carolinensis) in Eastern England. J. Anim. Ecol. 1985, 54, 149–162. [Google Scholar] [CrossRef]
- Rushton, S.P.; Lurz, P.W.W.; Gurnell, J.; Fuller, R. Modelling the spatial dynamics of parapoxvirus disease in red and grey squirrels: A possible cause of the decline in the red squirrel in the UK? J. Appl. Ecol. 2000, 37, 997–1012. [Google Scholar] [CrossRef]
- Tompkins, D.M.; White, A.R.; Boots, M. Ecological replacement of native red squirrels by invasive greys driven by disease. Ecol. Lett. 2003, 6, 189–196. [Google Scholar] [CrossRef]
- Jones, H.; White, A.; Lurz, P.; Shuttleworth, C. Mathematical models for invasive species management: Grey squirrel control on Anglesey. Ecol. Model. 2017, 359, 276–284. [Google Scholar] [CrossRef]
- Rushton, S.P.; Lurz, P.W.W.; Gurnell, J.; Nettleton, P.; Bruemmer, C.; Shirley, M.D.F.; Sainsbury, A.W. Disease threats posed by alien species: The role of a poxvirus in the decline of the native red squirrel in Britain. Epidemiol. Infect. 2006, 134, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, E.C. Red squirrel predation by pine martens in Scotland. In The Conservation of Red Squirrels, Sciurus vulgaris L.; PTES (People’s Trust for Endangered Species): London, UK, 1997; pp. 39–48. [Google Scholar]
- Lurz, P.W.W.; Garson, P.J.; Wauters, L.A. Effects of temporal and spatial variations in food supply on the space and habitat use of red squirrels (Sciurus vulgaris L.). J. Zool. 2000, 251, 167–178. [Google Scholar] [CrossRef]
- Wauters, L.A.; Gurnell, J.; Preatoni, D.; Tosi, G. Effects of spatial variation in food availability on spacing behavior and demography of Eurasian red squirrels. Ecography 2001, 24, 525–538. [Google Scholar] [CrossRef]
- Thompson, D.C. The social system of the grey squirrel. Behaviour 1978, 64, 305–328. [Google Scholar] [CrossRef]
- Koprowski, J.L. Sciurus carolinensis. Mamm. Species 1994, 480, 1. [Google Scholar] [CrossRef]
- Mosby, H.S. The influence of hunting on the population dynamics of a woodlot gray squirrel population. J. Wildl. Manag. 1969, 33, 709–717. [Google Scholar] [CrossRef]
- Montgomery, S.D.; Whelan, J.B.; Mosby, H.S. Bioenergetics of a woodlot gray squirrel population. J. Wildl. Manag. 1975, 39, 709. [Google Scholar] [CrossRef]
- Gurnell, J. The effects of food availability and winter weather on the dynamics of a grey squirrel population in Southern England. J. Appl. Ecol. 1996, 33, 325–338. [Google Scholar] [CrossRef]
- Bertolino, S. Attivazione di un Progetto di Monitoraggio Estansivo ed Intensivo dello Scoiattolo Grigio (Sciurus carolinensis) in Piemonte; Technical Report; Regione Piemonte: Torino, Italy, 2004. [Google Scholar]
- Lurz, P.W.W. The ecology of squirrels in spruce dominated plantations: Implications for forest management. For. Ecol. Manag. 1995, 79, 79–90. [Google Scholar] [CrossRef]
- Wauters, L.A.; Lens, L. Effects of food availability and density on red squirrel (Sciurus vulgaris) reproduction. Ecology 1995, 76, 2460–2469. [Google Scholar] [CrossRef]
- Lurz, P.; Garson, P.; Wauters, L. Effects of temporal and spatial variation in habitat quality on red squirrel dispersal behavior. Anim. Behav. 1997, 54, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Lurz, P.W.W.; Garson, P.J.; Ogilvie, J.F. Conifer species mixtures, cone crops and red squirrel conservation. Forestry 1998, 71, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Wauters, L.A.; Matthysen, E.; Adriaensen, F.; Tosi, G. Within-sex density dependence and population dynamics of red squirrels Sciurus vulgaris. J. Anim. Ecol. 2004, 73, 11–25. [Google Scholar] [CrossRef]
- Okubo, A.; Maini, P.K.; Williamson, M.H.; Murray, J.D. On the spatial spread of the grey squirrel in Britain. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1989, 238, 113–125. [Google Scholar] [CrossRef]
- Bryce, J.M.; Speakman, J.R.; Johnson, P.J.; Macdonald, D.W. Competition between Eurasian red and introduced Eastern grey squirrels: The energetic significance of body-mass differences. Proc. R. Soc. B Biol. Sci. 2001, 268, 1731–1736. [Google Scholar] [CrossRef] [PubMed]
- Atkin, J.W.; Radford, A.D.; Coyne, K.P.; Stavisky, J.; Chantrey, J. Detection of squirrel poxvirus by nested and real-time PCR from red (Sciurus vulgaris) and grey (Sciurus carolinensis) squirrels. BCM Vet. Res. 2010, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Schuchert, P.; Shuttleworth, C.M.; McInnes, C.J.; Everest, D.J.; Rushton, S.P. Landscape scale impacts of culling upon a European grey squirrel population: Can trapping reduce population size and decrease the threat of squirrelpox virus infection for the native red squirrel? Biol. Invasions 2014, 16, 2381–2391. [Google Scholar] [CrossRef]
- Gurnell, J. The Natural History of Squirrels; Helm: London, UK, 1987. [Google Scholar]
- Roberts, M.G.; Heesterbeek, J.A.P. Cpidemiology. J. Math. Biol. 2013, 66, 1045–1064. [Google Scholar] [CrossRef] [PubMed]
- Hilker, F.M.; Malchow, H. Strange Periodic Attractors in a Prey-Predator System with Infected Prey. Math. Popul. Stud. 2006, 13, 119–134. [Google Scholar] [CrossRef]
- Oliveira, N.M.; Hilker, F.M. Modelling Disease Introduction as Biological Control of Invasive Predators to Preserve Endangered Prey. Bull. Math. Biol. 2010, 72, 444–468. [Google Scholar] [CrossRef] [PubMed]
- White, A.; Lurz, P.W.W.; Jones, H.E.; Boots, M.; Bryce, J.; Tonkin, M.; Ramoo, K.; Bamforth, L.; Jarrott, A. The use of mathematical models in red squirrel conservation: Assessing the threat from grey invasion and disease to the Fleet basin stronghold. In Red Squirrels Ecology, Conservation Management in Europe; Shuttleworth, C., Lurz, P.W.W., Hayward, M.W., Eds.; European Squirrel Initiative (ESI): Woodbridge, UK, 2015; pp. 265–279. [Google Scholar]
- White, A.; Bell, S.S.; Lurz, P.W.W.; Boots, M. Conservation management within strongholds in the face of disease-mediated invasions: Red and grey squirrels as a case study. J. Appl. Ecol. 2014, 51, 1631–1642. [Google Scholar] [CrossRef]
- Massei, G.; Cowan, D.P.; Coats, J.; Gladwell, F.; Lane, J.E.; Miller, L.A. Effect of the GnRH vaccine GonaCon on the fertility, physiology and behavior of wild boar. Wildl. Res. 2008, 35, 540–547. [Google Scholar] [CrossRef]
- Krause, S.K.; Kelt, D.A.; Van Vuren, D.H.; Gionfriddo, J.P. Regulation of tree squirrel populations with immunocontraception: A fox squirrel example. Hum. Wildl. Interact. 2014, 8, 1–12. [Google Scholar]
- Moore, H.; Jenkins, N.M.; Wong, C. Immunocontraception in rodents: A review of the development of a sperm-based immunocontraceptive vaccine for the grey squirrel (Sciurus carolinensis). Reprod. Fertil. Dev. 1997, 9, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Cowan, D.P.; Massei, G. Wildlife contraception, individuals, and populations: How much fertility control is enough? In Proceedings of the 23rd Vertebrate Pest Conference, San Diego, CA, USA, 17–20 March 2008; pp. 220–228. [Google Scholar]
- Massei, G.; Cowan, D. Fertility control to mitigate human–wildlife conflicts: A review. Wildl. Res. 2014, 41, 1–21. [Google Scholar] [CrossRef]
- Pai, M. Field Evaluation of the Immunocontraceptive GonaConTM in Reducing Eastern Gray Squirrel Fecundity in Urban Areas. Ph.D. Thesis, Clemson University, Clemson, SC, USA, 2009. [Google Scholar]
- Pai, M.; Bruner, R.; Schlafer, D.H.; Yarrow, G.K.; Yoder, C.A.; Miller, L.A. Immunocontraception in Eastern gray squirrels (Sciurus carolinensis): Morphologic changes in reproductive organs. J. Zoo Wildl. Med. 2011, 42, 718–722. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, H.S.; Monteiro, M.T.T.; Torres, D.F. Dynamics of dengue epidemics when using optimal control. Math. Comput. Model. 2010, 52, 1667–1673. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, H.S.; Monteiro, M.T.T.; Torres, D.F.; Zinober, A. Dengue disease, basic reproduction number and control. Int. J. Comput. Math. 2012, 89, 334–346. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, H.S.; Monteiro, M.T.T.; Torres, D.F. Vaccination models and optimal control strategies to dengue. Math. Biosci. 2014, 247, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Interpretation | Unit |
|---|---|---|
| r | grey squirrels’ net growth rate | |
| E | grey squirrels’ carrying capacity | pure number |
| s | red squirrels’ net growth rate | |
| k | grey squirrels’ damage due to interspecific competition | |
| a | red squirrels’ damage due to interspecific competition | |
| K | red squirrels’ carrying capacity | pure number |
| intraspecific disease transmission rate for red squirrels | ||
| interspecific disease transmission rate for red squirrels | ||
| red squirrels’ natural plus disease-related mortality | ||
| c | competitivity weight among different squirrel species |
| Equilibrium | Feasibility | Stability |
|---|---|---|
| always | , | |
| , | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbara, F.; La Morgia, V.; Parodi, V.; Toscano, G.; Venturino, E. Analysis of the Incidence of Poxvirus on the Dynamics between Red and Grey Squirrels. Mathematics 2018, 6, 113. https://doi.org/10.3390/math6070113
Barbara F, La Morgia V, Parodi V, Toscano G, Venturino E. Analysis of the Incidence of Poxvirus on the Dynamics between Red and Grey Squirrels. Mathematics. 2018; 6(7):113. https://doi.org/10.3390/math6070113
Chicago/Turabian StyleBarbara, Fadi, Valentina La Morgia, Valerio Parodi, Giuseppe Toscano, and Ezio Venturino. 2018. "Analysis of the Incidence of Poxvirus on the Dynamics between Red and Grey Squirrels" Mathematics 6, no. 7: 113. https://doi.org/10.3390/math6070113
APA StyleBarbara, F., La Morgia, V., Parodi, V., Toscano, G., & Venturino, E. (2018). Analysis of the Incidence of Poxvirus on the Dynamics between Red and Grey Squirrels. Mathematics, 6(7), 113. https://doi.org/10.3390/math6070113