On a New Discrete SEIADR Model with Mixed Controls: Study of Its Properties





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- The characterization of the relations between the stability of the disease free equilibrium (DFE) point and the reachability of the endemic (END) one for the discrete SEIADR model under positive conditions.
- The study of the stability and positivity properties and the equilibrium points and their properties.
- The study through numerical examples of the influence of the controller gain in the equilibrium points and in the rates of convergence even under a similar reproduction number.
2. The Continuous SEIADR Model
- is the recruitment/birth rate,
- is the natural average death rate,
- are the disease transmission coefficients from the susceptible to the symptomatic and asymptomatic infectious, and to the infective corpses subpopulations, respectively,
- is the average duration of the immunity period which reflects a transition state from the recovered to the susceptible,
- is the transition rate from the exposed to the symptomatic and asymptomatic infectious,
- is the extra average mortality being associated with the disease which affects to the symptomatic infectious subpopulation,
- is the natural recovery rate for the whole infectious subpopulation (i.e., A + I ),
- p is the exposed subpopulation fraction which becomes symptomatic infectious,
- 1−p is the exposed subpopulation fraction which becomes asymptomatic infectious,
- 1/ is the average time of infectiousness after death,
- V, and are the constant vaccination gain, the proportional vaccination control gain and the antiviral treatment control gain respectively. The constant vaccination is bounded such that , so a fraction of the new individuals of the system (newborn, immigrants) is vaccinated.
- A disease-free equilibrium point given by wherewith the total population at this point defined as .
- An endemic one given by where:with the total population at this point given by:
3. Discretization of the SEIADR
- Normally feedback control actions are exerted by discrete-time controllers, especially, if the volume of data to be processed is relevant since the computational load has to be supported by a computer. Therefore, it can be preferred to start with a discrete-time model of the process, which then generates discrete sequences of measurable data, than a continuous-time one since then discrete control sequences are directly generated by processing the available sequence of discrete measurable data. Note also that the discretization of a continuous-time model towards the use of a computer for taking actions is always an approximation of the continuous time-model. However the sampling period of a discrete-time model is a design parameter, which does not imply an approximation when running the model.
- It could be argued that in fact the use of a continuous-time model can be used for control generations through a computer but, in this case, the discretization period has to be very small in order to consider approximately valid the continuous-time control generation from continuous-time data. That is, there is no freedom to select the discretization sampling period. Note that if a discrete controller is accommodated to a discrete-time model then there is an important freedom in the choice of the sampling period, which takes the role of an extra control parameter which can be eventually time-varying, if it is compatible with the stability and bandwidth.
- There is an important saving in data memory storage needs when implementing control actions, since only a discretized sequence of measurements needs to be stored and the control actions can be exerted along a set of time instants while the computer can exert alternative monitoring or computation actions. Thus, the computing time for control implementation is reduced.
Positivity of the Solution
4. Equilibrium Points: Positivity and Stability
4.1. Local Asymptotic Stability of the DFE Point
4.2. Conditions of Positivity of the Equilibrium Points
4.2.1. DFE Point
4.2.2. END Point
- , and
- for so that ifthen , since is non-decreasing with respect to for all real .
4.3. Global Stability
- (i)
- The total population is positive and bounded for any given non-negative initial conditions.
- (ii)
- The discrete SEIADR epidemic model is globally Lyapunov’s stable for any given finite non-negative initial conditions irrespective of the value of the reproduction number.
- (iii)
- if then the DFE point is the unique reachable equilibrium which is globally asymptotically stable.
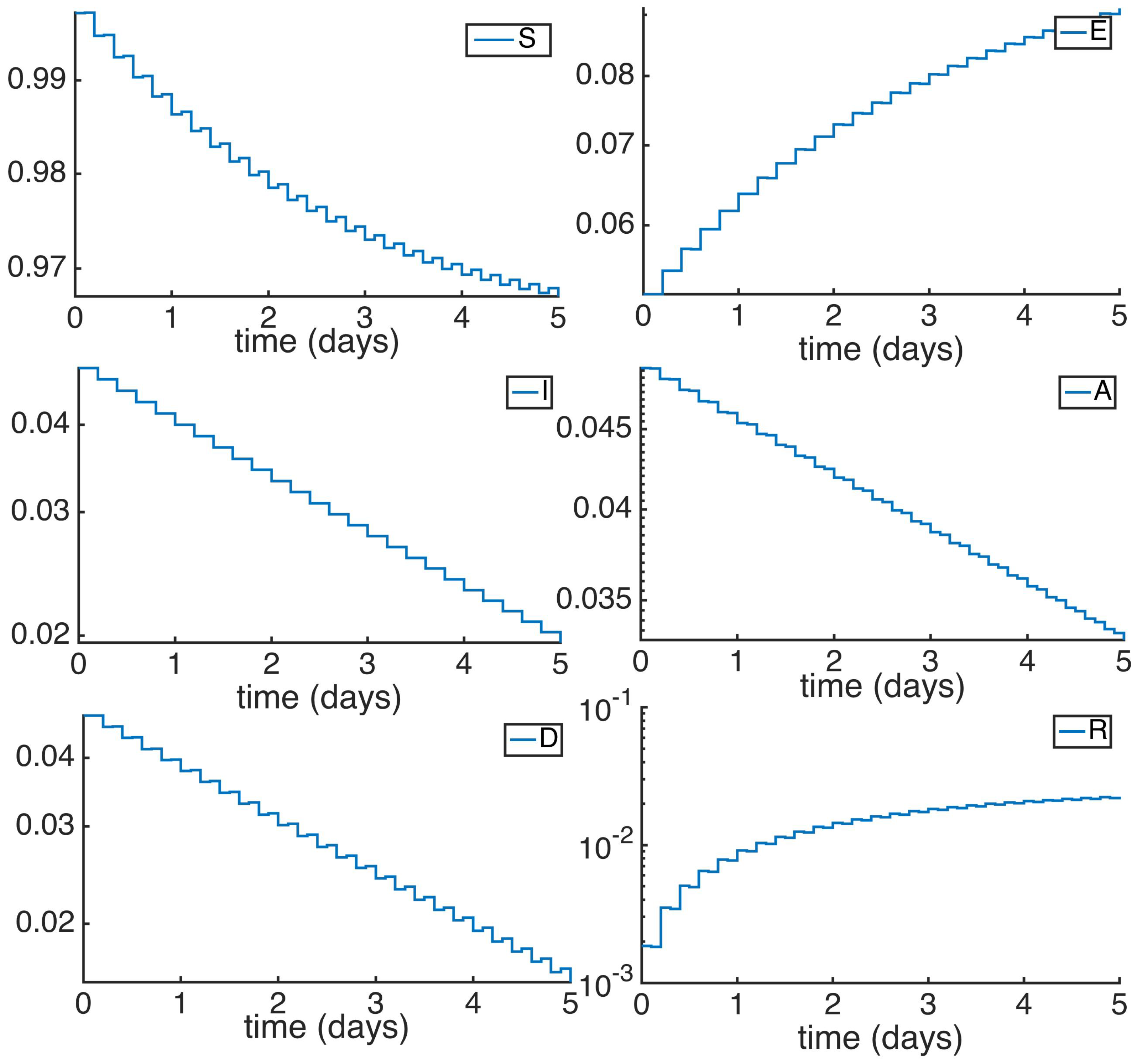
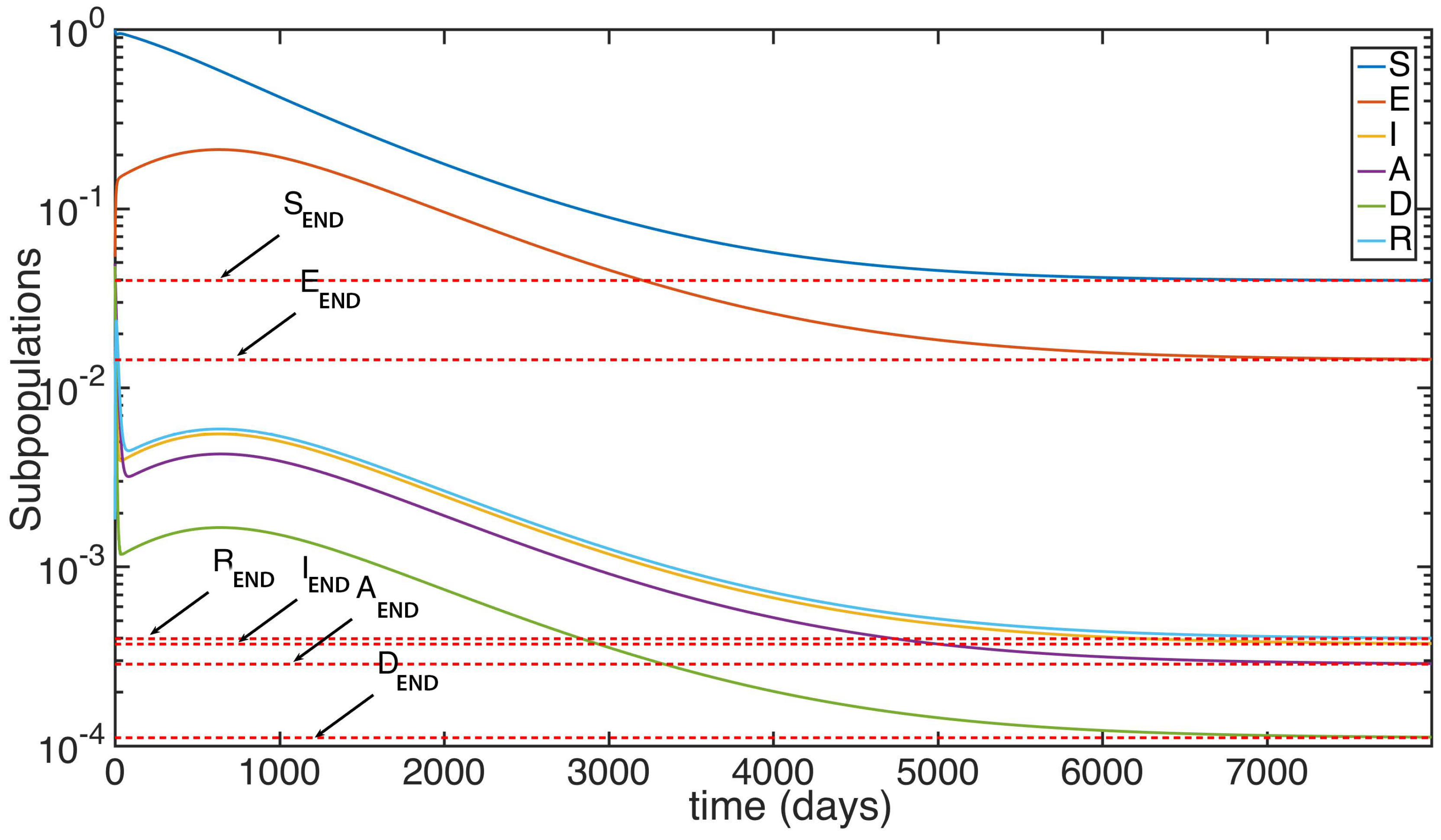
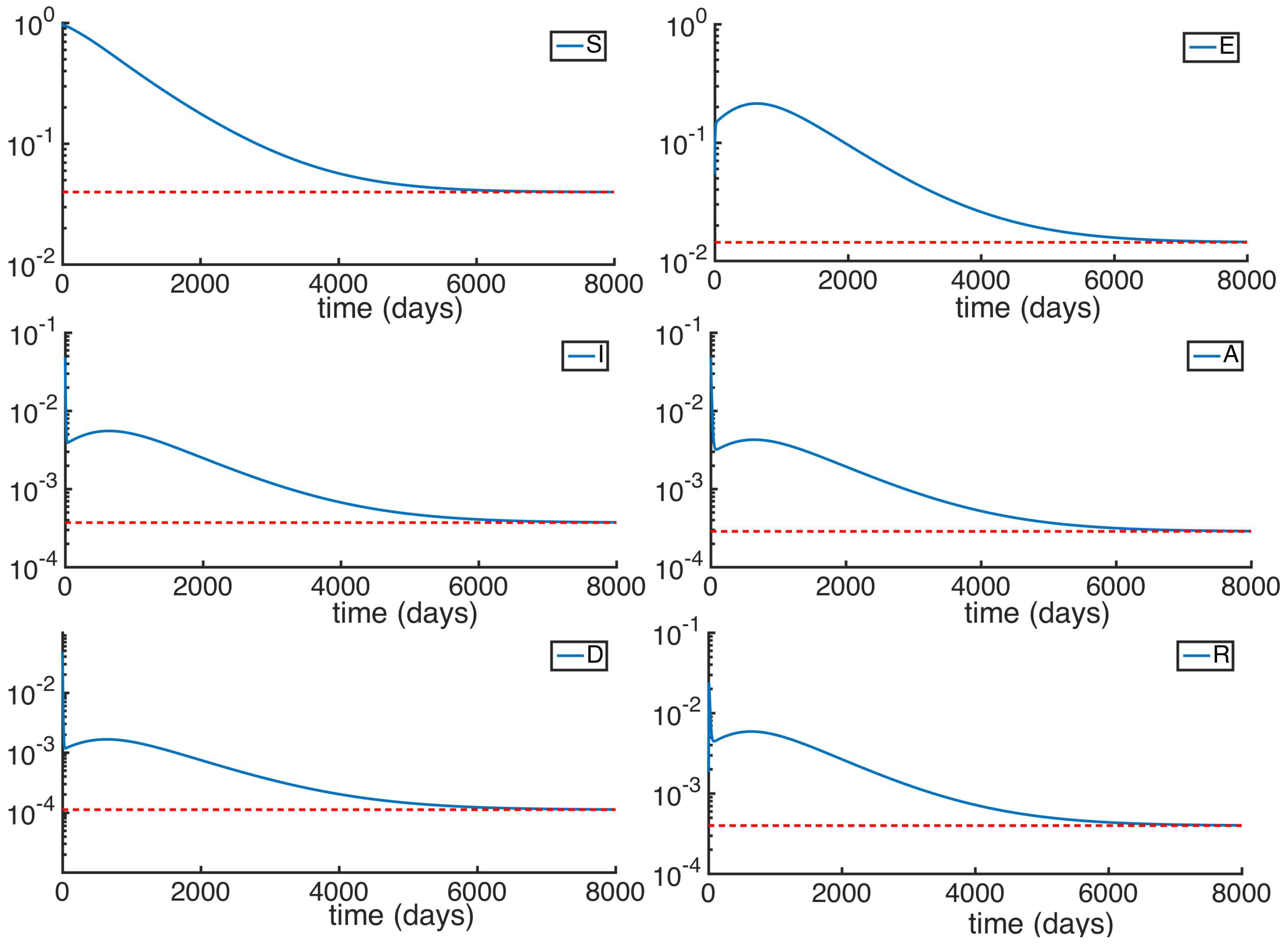
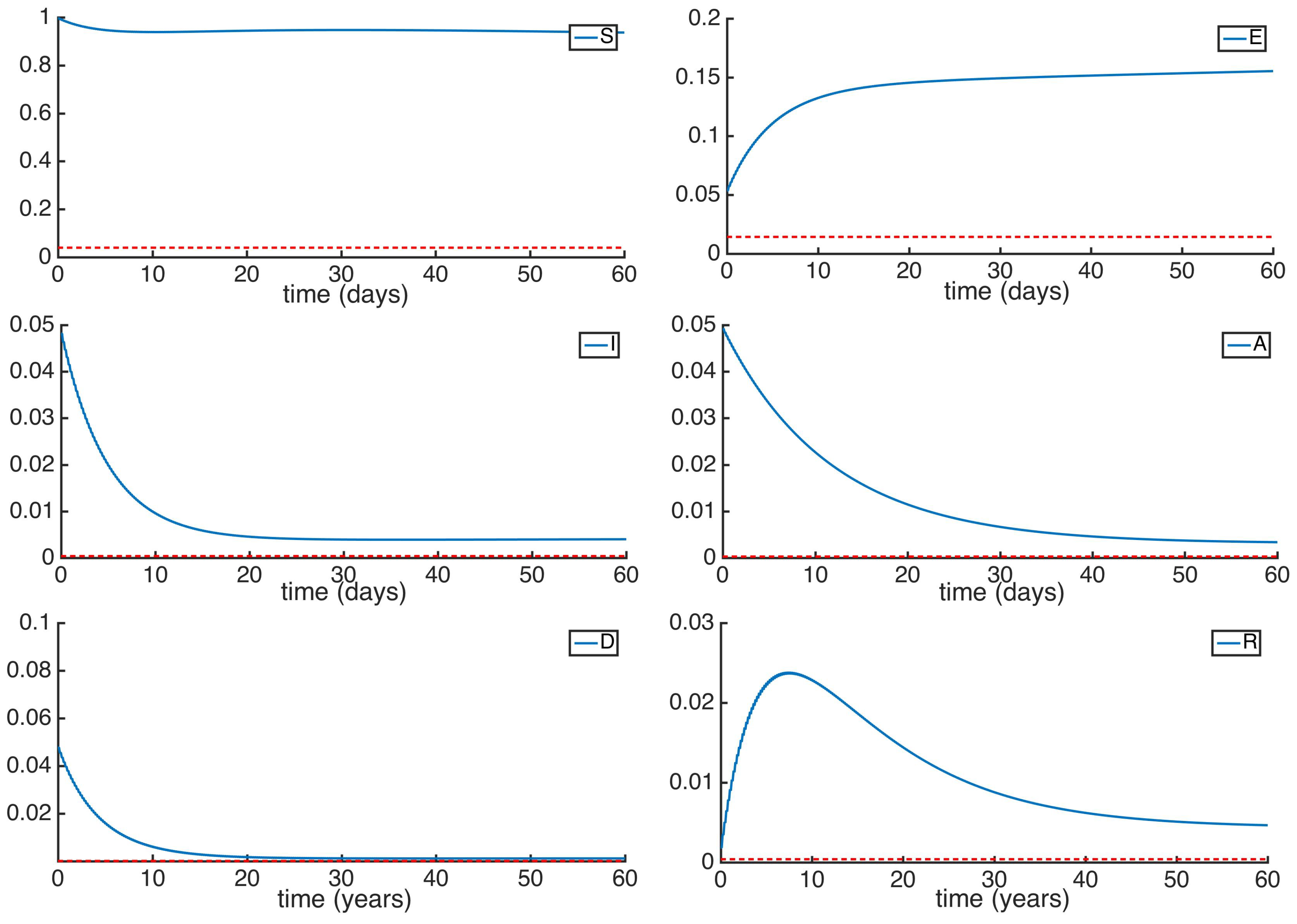
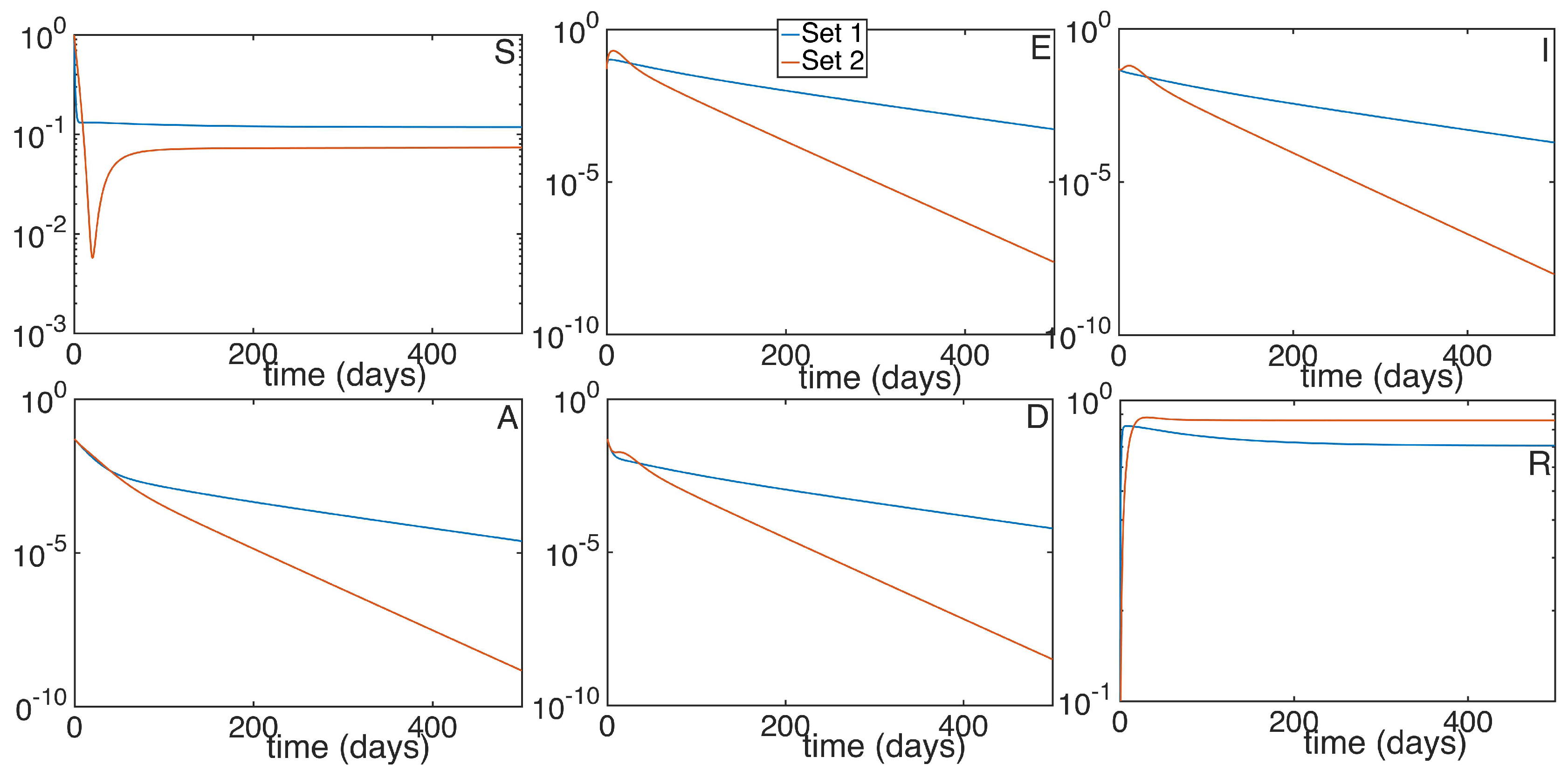
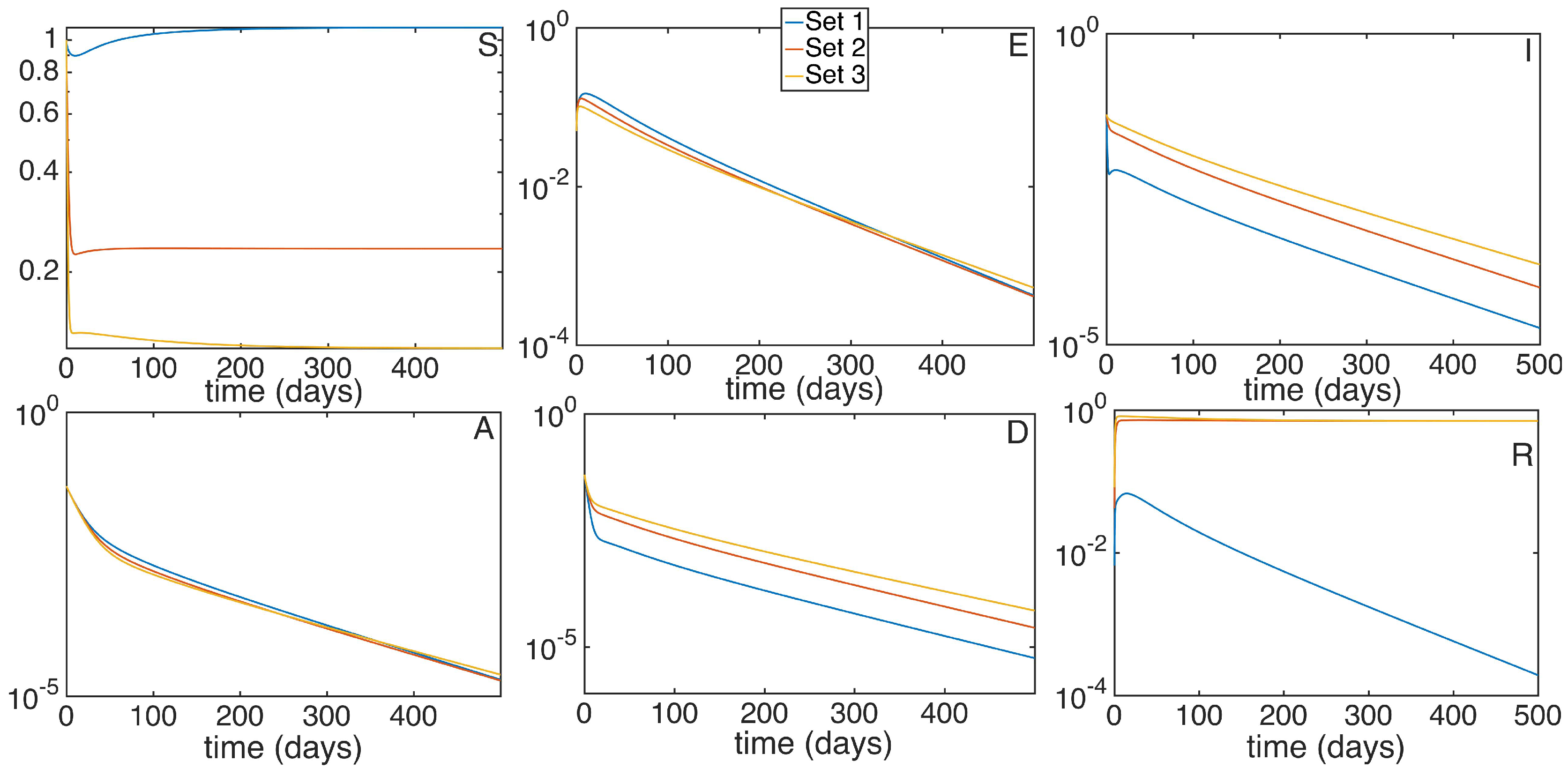
5. Numerical Simulations
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kermack, W.O.; McKendrick, A.G. A contribution to the mathematical theory of epidemics. Proc. R. Soc. Lond. Ser. A 1927, 115, 700–721. [Google Scholar] [CrossRef]
- Hethcote, H.W. The Mathematics of Infectious Diseases. SIAM Rev. 2000, 42, 599–653. [Google Scholar] [CrossRef] [Green Version]
- Keeling, M.J.; Rohani, P. Modeling Infectious Diseases in Humans and Animals; Princeton University Press: Princeton, NJ, USA; Oxford, UK, 2008. [Google Scholar]
- Hethcote, H.W. Three basic epidemiological models. Appl. Math. Ecol. 1989, 18, 119–144. [Google Scholar]
- Daley, D.; Gani, J. Epidemic Modeling: An Introduction; Cambridge Studies in Mathematical Biology: 15; Cambridge University Press: New York, NY, USA, 2005. [Google Scholar]
- Khan, H.; Mohapatra, R.; Vajravelu, K.; Liao, S.J. The explicit series solution of SIR and SIS epidemic models. Appl. Math. Comput. 2009, 215, 653–669. [Google Scholar] [CrossRef]
- Nistal, R.; De la Sen, M.; Alonso-Quesada, S.; Ibeas, A. A supervised multi-control for monitoring the antiviral treatment strategy for an SEIADR epidemic model. In Proceedings of the 2018 5th International Conference on Control, Decision and Information Technologies (CoDIT), Thessaloniki, Greece, 10–13 April 2018. [Google Scholar]
- Glynn, J.; Bower, H.; Johnson, S.; Houlihan, C.; Montesano, C.; Scott, J.T.; Semple, M.G.; Bangura, M.S.; Kamara, A.J.; Kamara, O.; et al. Asymptotic infection and unrecognised Ebola virus disease in Ebola-affected households in Sierra Leone: A cross-sectional study using new non-invasive assay for antibodies to Ebola virus. Lancet Infect. Dis. 2017, 6, 645–653. [Google Scholar] [CrossRef]
- de Pinho, M.; Maurer, H.; Kornienko, I. Optimal control of a SEIR model with mixed constraints and L1 cost. In Proceedings of the 11th Portuguese Conference on Automatic Control Lecture Notes in Electrical Engineering, Porto, Portugal, 21–23 July 2014. [Google Scholar]
- Leroy, E.; Baize, S.; Volchkov, V.; Fisher-Hoch, S.P.; Georges-Courbot, M.C.; Lansoud-Soukate, J.; Capron, M.; Debre, P.; Georges, A.J.; McCormick, J.B. Human asymptomatic Ebola infection and strong inflammatory response. Lancet 2000, 355, 2210–2215. [Google Scholar] [CrossRef]
- Bellan, S.; Pulliam, J.; Dushoff, J.; Meyers, L. Ebola control: Effect of asymptomatic infection and acquired immunity. Lancet 2014, 384, 1499–1500. [Google Scholar] [CrossRef]
- Santermans, E.; Robesyn, E.; Sudre, T.G.B.; Faes, C.; Quinten, C.; Van Bortel, W.; Haber, T.; Kovac, T.; Reeth, F.V.; Testa, M.; et al. Spatiotemporal evolution of Ebola disease at sub-national level during the 2014 West Africa epidemic: Model scrutiny and data meagerness. PLoS ONE 2016, 11, e0147172. [Google Scholar] [CrossRef]
- Al-Darabsah, I.; Yuan, Y. A time-delayed epidemic model for Ebola disease transmission. Appl. Math. Comput. 2016, 290, 307–325. [Google Scholar] [CrossRef]
- De la Sen, M.; Agarwal, R.P.; Ibeas, A.; Alonso-Quesada, S. On the existence of equilibrium points, boundedness, oscillating behaviour and positivity of a SVEIRS epidemic model under constant and impulsive vaccination. Adv. Differ. Equ. 2011, 2011, 748608. [Google Scholar] [CrossRef]
- De la Sen, M.; Alonso-Quesada, S. Vaccination strategies based on feedback control techniques for a SEIR-epidemic model. Appl. Math. Comput. 2011, 218, 3888–3904. [Google Scholar] [CrossRef]
- Wei, Z.; Le, M. Existence and Convergence of the Positive Solutions of a Discrete Epidemic Model. Discret. Dyn. Nat. Soc. 2015, 2015, 434537. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Z.; Zhang, X. Global dynamics of an SVEIR epidemic model with distributed delay and nonlinear incidence. Appl. Math. Comput. 2016, 2016, 47–65. [Google Scholar] [CrossRef]
- Wang, X. An SIRS Epidemic Model with Vital Dynamics and a Ratio-Dependent Saturation Incidence Rate. Discret. Dyn. Nat. Soc. 2015, 2015, 720682. [Google Scholar] [CrossRef]
- Fengying, W.; Chen, F. Stochastic permanence of an SIQS epidemic model with saturated incidence and independent random perturbations. Phys. A Stat. Mech. Its Appl. 2016, 453, 99–107. [Google Scholar]
- De la Sen, M.; Alonso-Quesada, S.; Ibeas, A. On the stability of an SEIR epidemic model with distributed time-delay and a general class of feedback vaccination rules. Appl. Math. Comput. 2015, 270, 953–976. [Google Scholar] [CrossRef]
- Shaikhet, L. Stability of equilibrium states for a stochastically perturbed exponential type system of differential equations. J. Comput. Appl. Math. 2015, 290, 92–103. [Google Scholar] [CrossRef]
- Shaikhet, L.; Korobeinikov, A. Stability of a stochastic model for HIV-1 dynamics within a host. Appl. Anal. 2016, 95, 1228–1238. [Google Scholar] [CrossRef]
- De la Sen, M.; Agarwal, R.P.; Ibeas, A.; Alonso-Quesada, S. On a generalized time-varying SEIR epidemic model with mixed point and distributed time-varying delays and combined regular and impulsive vaccination. Adv. Diff. Equ. 2010, 2010, 281612. [Google Scholar] [CrossRef]
- Tripathi, J.; Abbas, S. Global dynamics of autonomous and nonautonomous SI epidemic models with nonlinear incidence rate and feedback controls. Nonlinear Dyn. 2016, 86, 337–351. [Google Scholar] [CrossRef]
- Buonomo, B.; Lacitignola, D.; de Leon, C.V. Qualitative analysis and optimal control of an epidemic model with vaccination and treatment. Math. Comput. Simul. 2014, 100, 88–102. [Google Scholar] [CrossRef]
- Ling, L.; Jiang, G.; Long, T. The dynamics of an SIS epidemic model with fixed-time birth pulses and state feedback pulse treatments. Appl. Math. Model. 2015, 39, 5579–5591. [Google Scholar] [CrossRef]
- He, Y.; Gao, S.; Xie, D. An SIR epidemic model with time-varying pulse control schemes and saturated infectious force. Appl. Math. Model. 2013, 37, 8131–8140. [Google Scholar] [CrossRef]
- Sharma, S.; Samanta, G. Stability analysis and optimal control of an epidemic model with vaccination. Int. J. Biomath. 2015, 8, 1550030. [Google Scholar] [CrossRef]
- Samanta, G. A delayed hand-foot-mouth disease model with pulse vaccination strategy. Comput. Appl. Math. 2015, 34, 1131–1152. [Google Scholar] [CrossRef]
- De la Sen, M.; Ibeas, A. On the global asymptotic stability of switched linear time-varying systems with constant point delays. Discret. Dyn. Nat. Soc. 2008, 2008, 231710. [Google Scholar] [CrossRef]
- Bilbao-Guillerna, A.; De la Sen, M.; Ibeas, A.; Alonso-Quesada, S. Robustly stable multiestimation scheme for adaptive control and identification. Discret. Dyn. Nat. Soc. 2005, 2015, 31–67. [Google Scholar] [CrossRef]
- Bilbao-Guillerna, A.; De la Sen, M.; Alonso-Quesada, S. Multimodel discrete control with online updating of the fractional order hold gains. Cybern. Syst. 2007, 38, 249–274. [Google Scholar] [CrossRef]
- Herrera, J.; Ibeas, A.; Alcantara, S.; De la Sen, M. Multimodel-based techniques for the identification and adaptive control of delayed multi-input multi-output systems. IET Control Theory Appl. 2011, 5, 188–202. [Google Scholar] [CrossRef]
- Jang, S. On a discrete west Nile epidemic model. Comput. Appl. Math. 2007, 6, 397–414. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, L.; Han, Z. Stability analysis of two new SIRS models with two viruses. Int. J. Comput. Math. 2018, 95, 2026–2035. [Google Scholar] [CrossRef]
- Degue, K.; Ny, J.L. An interval observer for discrete-time SEIR epidemic model. In Proceedings of the 2018 American Control Conference (ACC), Milwaukee, WI, USA, 27–29 June 2018; pp. 5934–5939. [Google Scholar]
- Chang, C.; Jing, Y.; Zhu, B. Modeling and control for a descriptor epidemic system with nonlinear incidence rate. In Proceedings of the 2018 Chinese Control Conference (CCDC), Shenyang, China, 27–29 June 2018; pp. 2183–2188. [Google Scholar]
- Abdelaziz, M.; Ismail, A.I.; Abdullah, F.; Mohd, M.H. Bifurcations and chaos in a discrete SI epidemic model with fractional order. Adv. Diff. Equ. 2018, 44, 1–19. [Google Scholar] [CrossRef]
- Chiranjeevi, T.; Biswas, B. Discrete-time fractional optimal control. Mathematics 2017, 5, 25. [Google Scholar] [CrossRef]
- De la Sen, M. Preserving positive realness through discretization. Positivity 2002, 6, 31–45. [Google Scholar] [CrossRef]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nistal, R.; De la Sen, M.; Alonso-Quesada, S.; Ibeas, A. On a New Discrete SEIADR Model with Mixed Controls: Study of Its Properties. Mathematics 2019, 7, 18. https://doi.org/10.3390/math7010018
Nistal R, De la Sen M, Alonso-Quesada S, Ibeas A. On a New Discrete SEIADR Model with Mixed Controls: Study of Its Properties. Mathematics. 2019; 7(1):18. https://doi.org/10.3390/math7010018
Chicago/Turabian StyleNistal, Raul, Manuel De la Sen, Santiago Alonso-Quesada, and Asier Ibeas. 2019. "On a New Discrete SEIADR Model with Mixed Controls: Study of Its Properties" Mathematics 7, no. 1: 18. https://doi.org/10.3390/math7010018
APA StyleNistal, R., De la Sen, M., Alonso-Quesada, S., & Ibeas, A. (2019). On a New Discrete SEIADR Model with Mixed Controls: Study of Its Properties. Mathematics, 7(1), 18. https://doi.org/10.3390/math7010018