Mathematical Modelling of the Structure and Function of the Lymphatic System


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
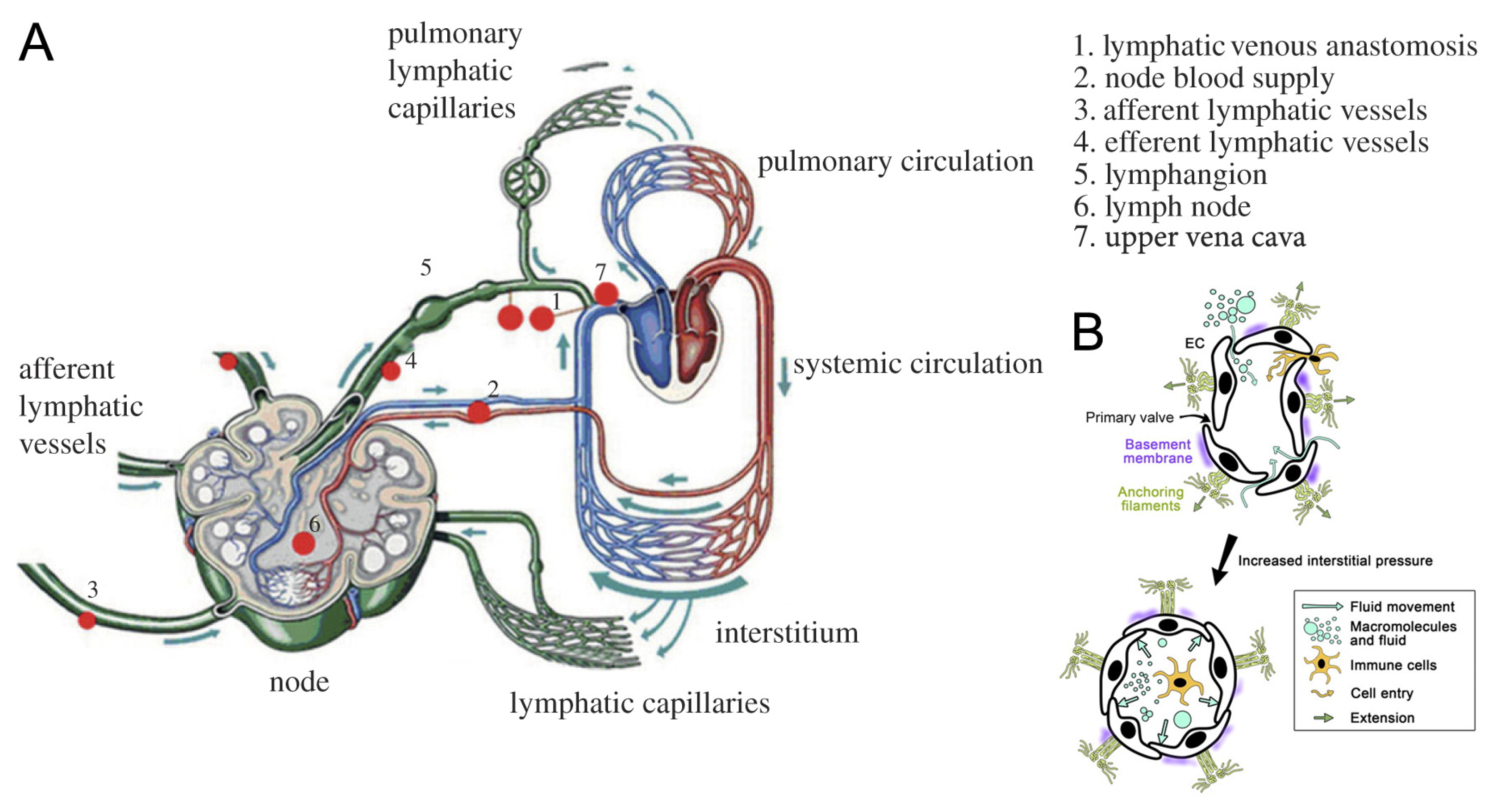
2. Transport Function of the LS
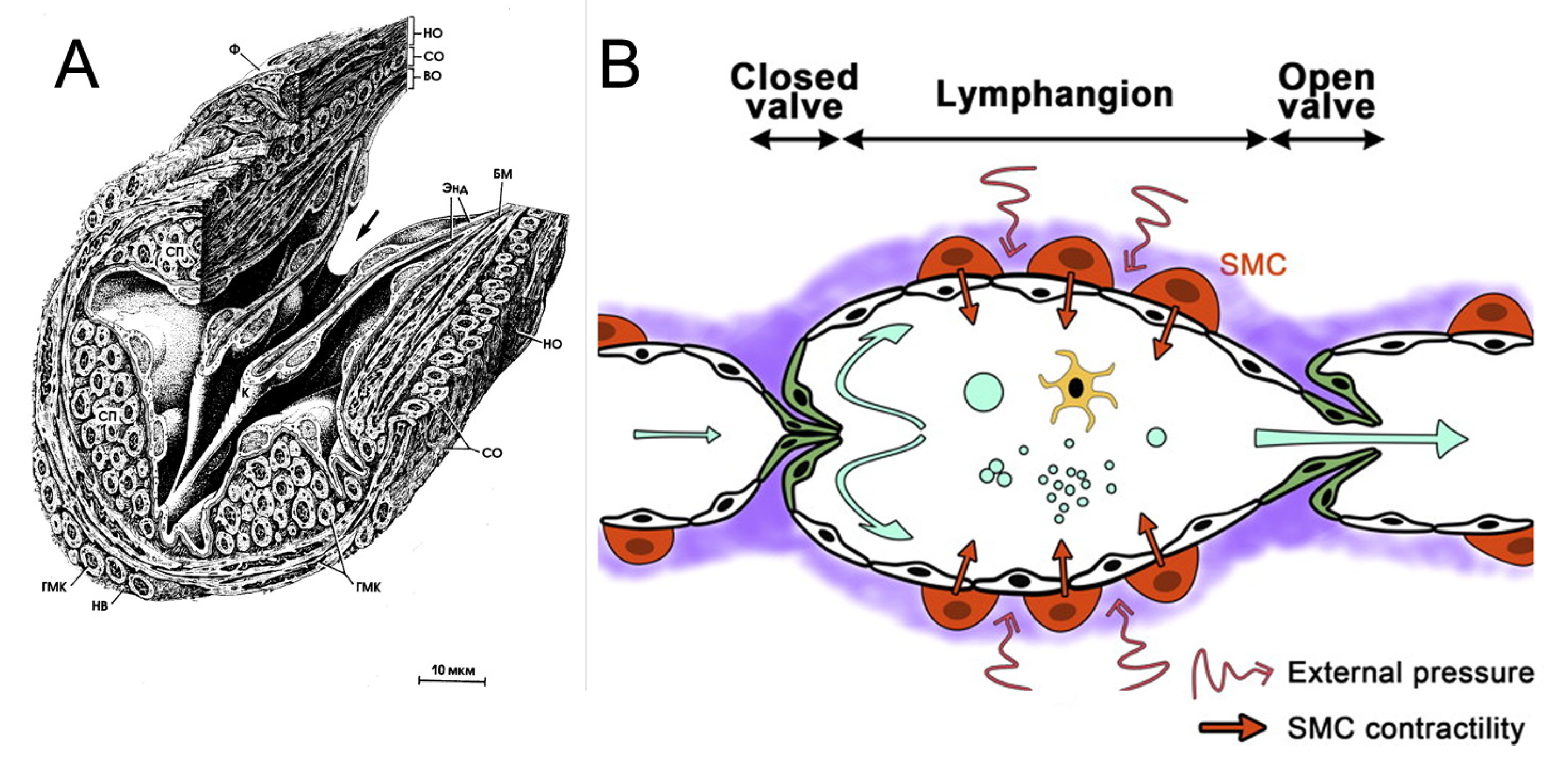
3. Flow in Lymphangions
3.1. Influence of Valves
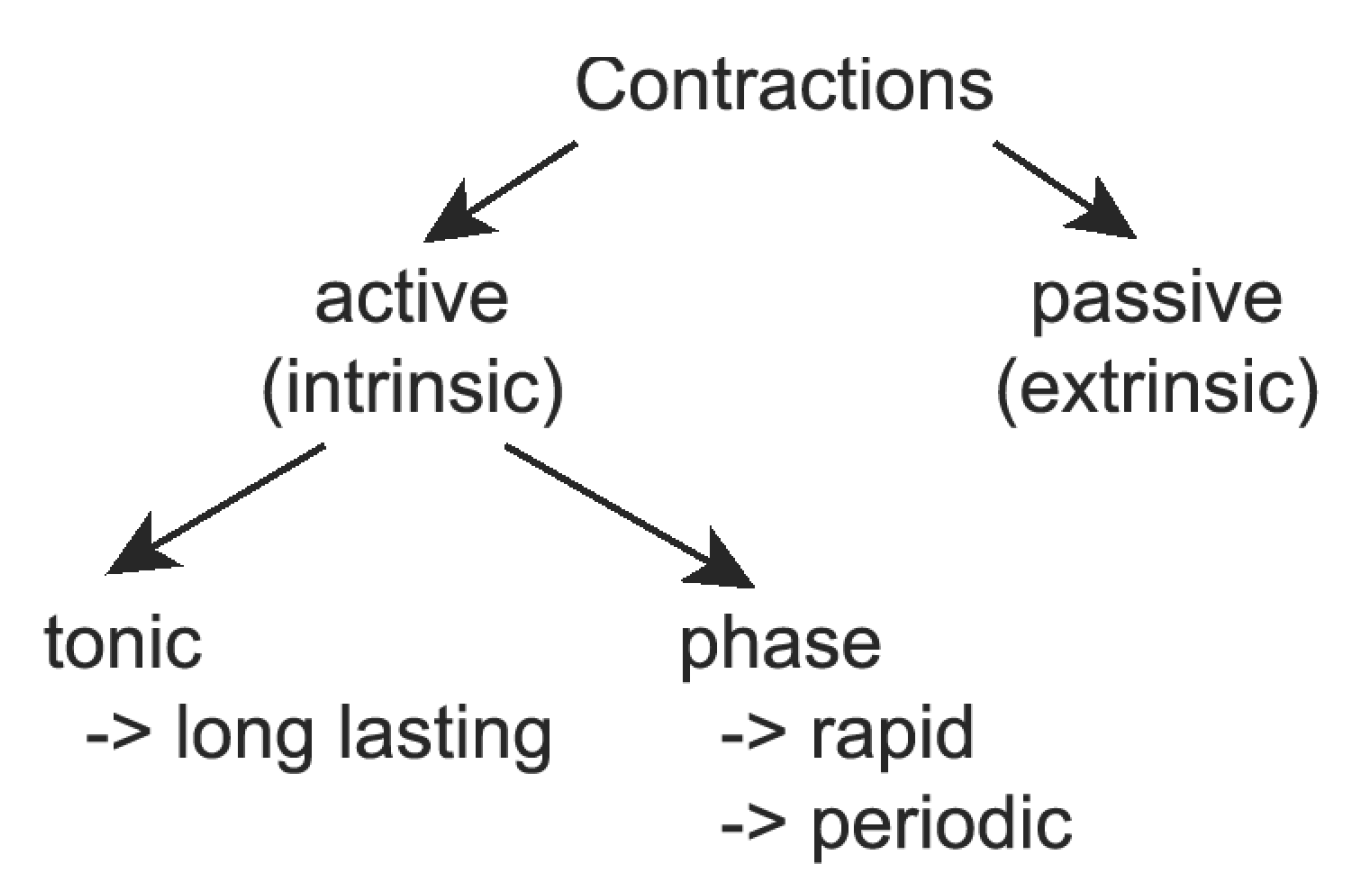
3.2. Influence of Contractions
3.2.1. Modelling of Tonic Response
3.2.2. Phase Contractions
3.2.3. Passive Contractions
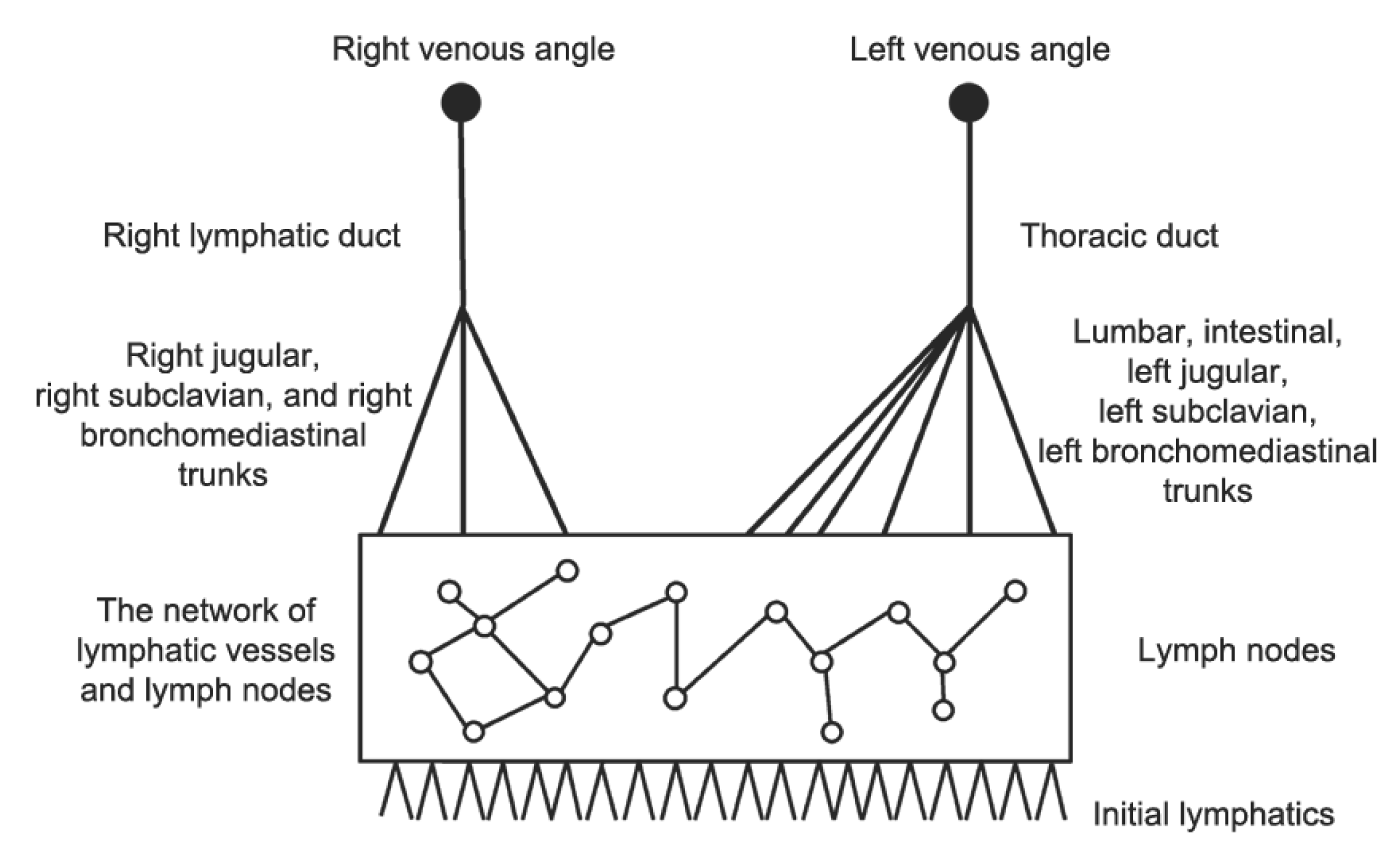
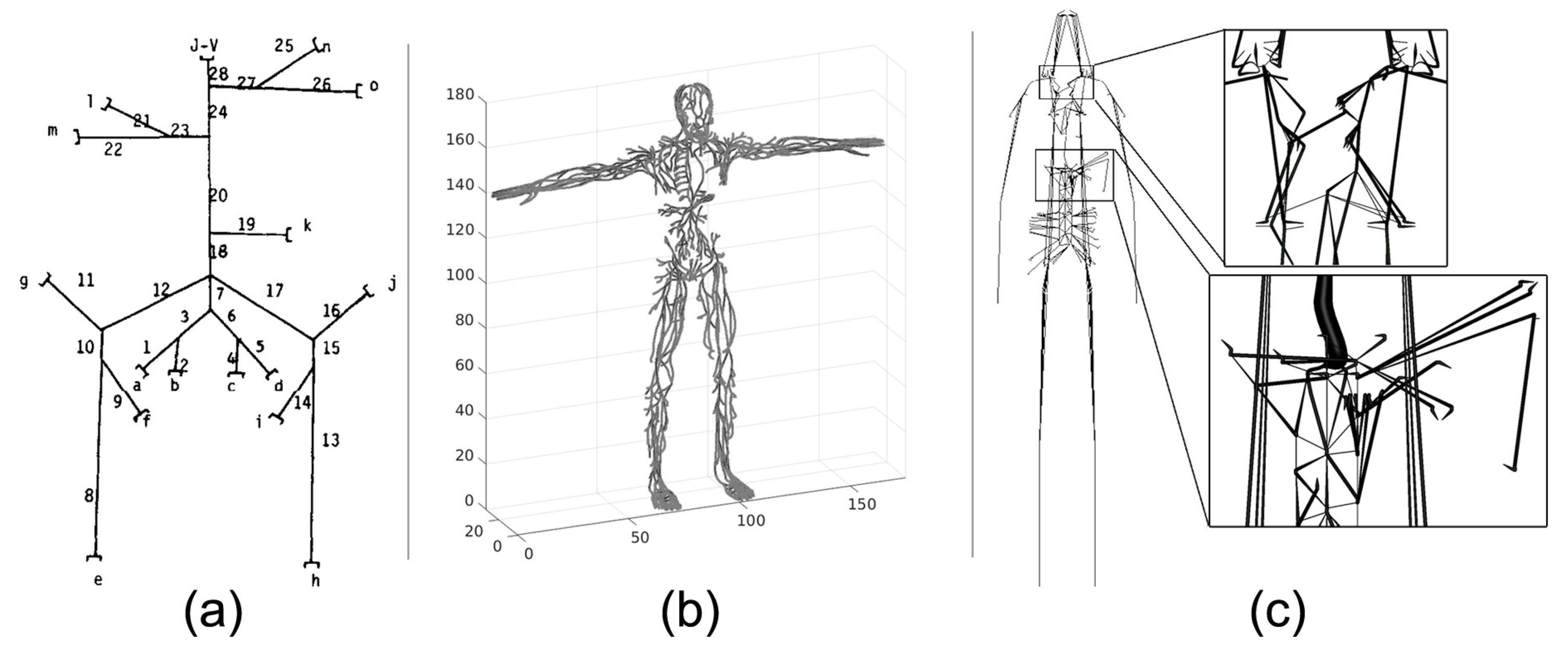
4. Systemic Lymph Flow and Graphs of the LS
5. Lymph Drainage and Flow in Initial Lymphatics
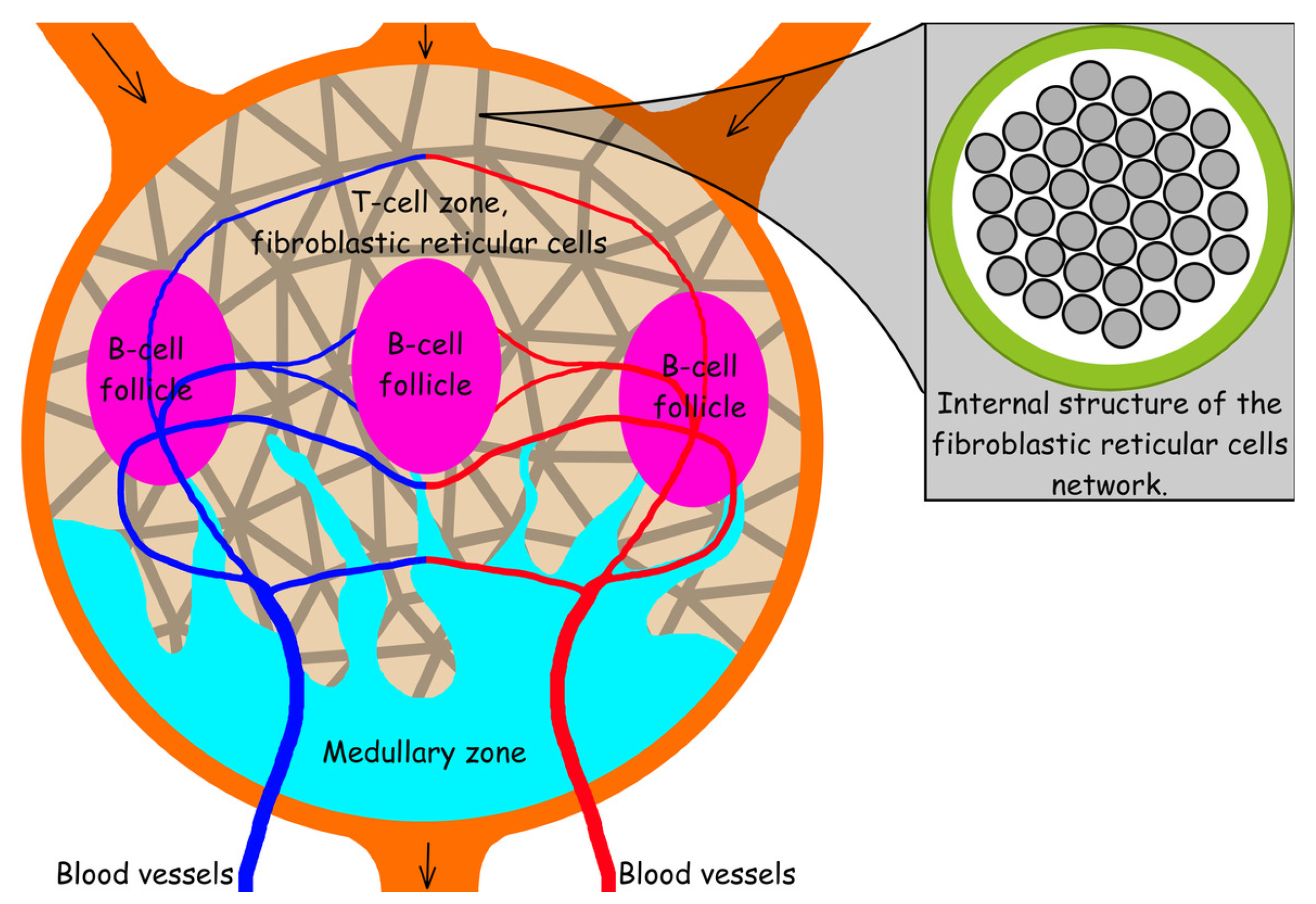
6. The Structure and Function of the Lymph Node
7. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Choi, I.; Lee, S.; Hong, Y.K. The New Era of the Lymphatic System: No Longer Secondary to the Blood Vascular System. Cold Spring Harb. Perspect. Med. 2012, 2, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filchenkov, A.A. Lymphangiogenesis and metastasis of tumors. Creat. Surg. Oncol. 2010, No. 3. 80–90. (In Russian) [Google Scholar]
- Itkin, M.; McCormack, F.; Dori, Y. Diagnosis and Treatment of Lymphatic Plastic Bronchitis in Adults Using Advanced Lymphatic Imaging and Percutaneous Embolization. Ann. Am. Thorac. Soc. 2016, 13, 1689–1696. [Google Scholar] [CrossRef] [PubMed]
- Pamarthi, V.; Pabon-Ramos, W.M.; Marnell, V.; Hurwitz, L.M. MRI of the Central Lymphatic System: Indications, Imaging Technique, and Pre-Procedural Planning. Top. Magn. Reson. Imaging 2017, 26, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Louveau, A.; Smirnov, I.; Keyes, T.J.; Eccles, J.D.; Rouhani, S.J.; Peske, J.D.; Derecki, N.C.; Castle, D.; Mandell, J.W.; Lee, K.S.; et al. Structural and functional features of central nervous system lymphatic vessels. Nature 2015, 523, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Aspelund, A.; Antila, S.; Proulx, S.T.; Karlsen, T.V.; Karaman, S.; Detmar, M.; Wiig, H.; Alitalo, K. A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J. Exp. Med. 2015, 212, 991–999. [Google Scholar] [CrossRef] [PubMed]
- Nikolenko, V.N.; Oganesyan, M.V.; Yakhno, N.N.; Orlov, E.A.; Porubayeva, E.E.; Popova, E.Y. The brain’ sglymphatic system: Physiological anatomy and clinical perspectives. Neurol. Neuropsychiatry Psychosom. 2018, 10, 94–100. [Google Scholar] [CrossRef]
- Munn, L.L.; Padera, T.P. Imaging the lymphatic system. Microvasc. Res. 2014, 96, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Wendt, J.A.; Rasmussen, J.C.; Adams, K.E.; Marshall, M.V.; Sevick-Muraca, E.M. New Horizons for Imaging Lymphatic Function. Ann. N. Y. Acad. Sci. 2008, 1131, 13–36. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.F.; Lu, Q.; Jiang, Z.H.; Wang, C.G.; Zhou, J.G. Anatomic and functional evaluation of the lymphatics and lymph nodes in diagnosis of lymphatic circulation disorders with contrast magnetic resonance lymphangiography. J. Vasc. Surg. 2009, 49, 980–987. [Google Scholar] [CrossRef] [Green Version]
- Sevick-Muraca, E.M.; Sharma, R.; Rasmussen, J.C.; Marshall, M.V.; Wendt, J.A.; Pham, H.Q.; Bonefas, E.; Houston, J.P.; Sampath, L.; Adams, K.E.; et al. Imaging of Lymph Flow in Breast Cancer Patients after Microdose Administration of a Near-Infrared Fluorophore: Feasibility Study. Radiology 2008, 246, 734–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, S.; Sevick-Muraca, E.M. Noninvasive Quantitative Imaging of Lymph Function in Mice. Lymphat. Res. Biol. 2007, 5, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Wang, W.; Rasmussen, J.C.; Joshi, A.; Houston, J.P.; Adams, K.E.; Cameron, A.; Ke, S.; Kwon, S.; Mawad, M.E.; et al. Quantitative imaging of lymph function. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H3109–H3118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, J.B.; Zawieja, D.C.; Gashev, A.A.; Cote, G.L. Measuring microlymphatic flow using fast video microscopy. J. Biomed. Opt. 2005, 10, 064016. [Google Scholar] [CrossRef] [Green Version]
- Zawieja, S.D.; Castorena-Gonzalez, J.A.; Dixon, B.; Davis, M.J. Experimental Models Used to Assess Lymphatic Contractile Function. Lymphat. Res. Biol. 2017, 15, 331–342. [Google Scholar] [CrossRef]
- Blatter, C.; Meijer, E.F.J.; Nam, A.S.; Jones, D.; Bouma, B.E.; Padera, T.P.; Vakoc, B.J. In vivo label-free measurement of lymph flow velocity and volumetric flow rates using Doppler optical coherence tomography. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Quick, C.M.; Venugopal, A.M.; Gashev, A.A.; Zawieja, D.C.; Stewart, R.H. Intrinsic pump-conduit behavior of lymphangions. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R1510–R1518. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, A.J.; Arkill, K.P.; Tabor, G.R.; McHale, N.G.; Winlove, C.P. Modeling flow in collecting lymphatic vessels: One-dimensional flow through a series of contractile elements. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H305–H313. [Google Scholar] [CrossRef] [Green Version]
- Lobov, G.I.; Pan’kova, M.N.; Abdreshov, S.N. Phase and tonic contractions of lymphatic vessels and nodes under the action of atrial natriuretic peptide. Reg. Blood Circ. Microcirc. 2015, 14, 72–77. (In Russian) [Google Scholar] [CrossRef] [Green Version]
- Ohhashi, T.; Mizuno, R.; Ikomi, F.; Kawai, Y. Current topics of physiology and pharmacology in the lymphatic system. Pharmacol. Ther. 2005, 105, 165–188. [Google Scholar] [CrossRef] [Green Version]
- Venugopal, A.M.; Stewart, R.H.; Laine, G.A.; Dongaonkar, R.M.; Quick, C.M. Lymphangion coordination minimally affects mean flow in lymphatic vessels. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H1183–H1189. [Google Scholar] [CrossRef]
- Bertram, C.D.; Macaskill, C.; Moore, J.E. Simulation of a Chain of Collapsible Contracting Lymphangions With Progressive Valve Closure. J. Biomech. Eng. 2010, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margaris, K.N.; Black, R.A. Modelling the lymphatic system: Challenges and opportunities. J. R. Soc. Interface 2012, 9, 601–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roose, T.; Tabor, G. Multiscale Modelling of Lymphatic Drainage. In Multiscale Computer Modeling in Biomechanics and Biomedical Engineering; Springer: Berlin/Heidelberg, Germany, 2012; pp. 149–176. [Google Scholar] [CrossRef]
- Guyton, A.C.; Hall, J.E. Textbook of Medical Physiology; Logosphere: Moscow, Russia, 2008; p. 1296. (In Russian) [Google Scholar]
- Schmidt, R.; Thews, G. Human Physiology; Mir: Moscow, Russia, 2005; Volume 2, p. 314. (In Russian) [Google Scholar]
- McKinley, M.; O’Loughlin, V.D. Human Anatomy; McGraw-Hill: New York, NY, USA, 2012; p. 966. [Google Scholar]
- Moore, J.E.; Bertram, C.D. Lymphatic System Flows. Ann. Rev. Fluid Mech. 2018, 50, 459–482. [Google Scholar] [CrossRef]
- Borzyak, E.; Bocharov, V.; Sapin, M. Human Anatomy; Medicine: Moscow, Russia, 1993; Volume 2, p. 560. (In Russian) [Google Scholar]
- Sinelnikov, R.; Sinelnikov, Y. Atlas of Human Anatomy. The Doctrine of the Vessels; Medicine: Moscow, Russia, 1996; Volume 3, p. 219. (In Russian) [Google Scholar]
- Petrenko, V.M. Functional Morphology of Lymphatic Vessels; DEAN: Saint Petersburg, Russia, 2008; p. 400. (In Russian) [Google Scholar]
- Lobov, G.I.; Pan’kova, M.N. Lymph transport in lymphatic nodes: Mechanisms of regulation. Ross. Fiziol. Zhurnal Im. I.M. Sechenova 2013, 98, 1350–1361. (In Russian) [Google Scholar]
- Zawieja, D.C. Contractile Physiology of Lymphatics. Lymphat. Res. Biol. 2009, 7, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quéré, I. Description anatomique et histologique, physiologie du système lymphatique. La Presse Médicale 2010, 39, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Schulte-Merker, S.; Sabine, A.; Petrova, T.V. Lymphatic vascular morphogenesis in development, physiology, and disease. J. Cell Biol. 2011, 193, 607–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krstic, R. Human Microscopic Anatomy: An Atlas for Students of Medicine and Biology; World and Education: Moscow, Russia, 2010; p. 608. (In Russian) [Google Scholar]
- Reddy, N.P.; Krouskop, T.A.; Paul, H. Newell, j. Biomechanics of a Lymphatic Vessel. J. Vasc. Res. 1975, 12, 261–278. [Google Scholar] [CrossRef]
- Gajani, G.S.; Boschetti, F.; Negrini, D.; Martellaccio, R.; Milanese, G.; Bizzarri, F.; Brambilla, A. A lumped model of lymphatic systems suitable for large scale simulations. In Proceedings of the 2015 European Conference on Circuit Theory and Design (ECCTD), Trondheim, Norway, 24–26 August 2015. [Google Scholar] [CrossRef]
- Contarino, C.; Toro, E.F. A one-dimensional mathematical model of collecting lymphatics coupled with an electro-fluid-mechanical contraction model and valve dynamics. Biomech. Model. Mechanobiol. 2018, 17, 1687–1714. [Google Scholar] [CrossRef] [Green Version]
- Mozokhina, A.S.; Mukhin, S.I. Pressure Gradient Influence on Global Lymph Flow. In Trends in Biomathematics: Modeling, Optimization and Computational Problems; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; pp. 325–334. [Google Scholar] [CrossRef]
- Tretyakova, R.M.; Lobov, G.I.; Bocharov, G.A. Modelling lymph flow in the lymphatic system: From 0D to 1D spatial resolution. Math. Model. Nat. Phenom. 2018, 13, 45. [Google Scholar] [CrossRef]
- Milišić, V.; Quarteroni, A. Analysis of lumped parameter models for blood flow simulations and their relation with 1D models. ESAIM Math. Model. Numer. Anal. 2004, 38, 613–632. [Google Scholar] [CrossRef]
- Kokalari, I.; Karaja, T.; Guerrisi, M. Review on lumped parameter method for modeling the blood flow in systemic arteries. J. Biomed. Sci. Eng. 2013, 6, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Rahbar, E.; Moore, J.E.J. A model of a radially expanding and contracting lymphangion. J. Biomech. 2011, 44, 1001–1007. [Google Scholar] [CrossRef] [Green Version]
- Reddy, N.P.; Krouskop, T.A.; Newell, P.H. A computer model of the lymphatic system. Comput. Biol. Med. 1977, 7, 181–197. [Google Scholar] [CrossRef]
- Sherwin, S.; Franke, V.; Peiró, J.; Parker, K. One-dimensional modelling of a vascular network in space-time variables. J. Eng. Math. 2003, 47, 217–250. [Google Scholar] [CrossRef]
- Bunicheva, A.Y.; Mukhin, S.I.; Sosnin, N.V.; Khrulenko, A.B. Mathematical modeling of quasi-one-dimensional hemodynamics. Comput. Math. Math. Phys. 2015, 55, 1381–1392. [Google Scholar] [CrossRef]
- Barnard, A.L.; Hunt, W.; Timlake, W.; Varley, E. A Theory of Fluid Flow in Compliant Tubes. Biophys. J. 1966, 6, 717–724. [Google Scholar] [CrossRef] [Green Version]
- Phionik, O. Clinical and Morpho-Functional Bases for Diagnostic and Treatment of Lymphedema of Low Limbs. Ph.D. Thesis, St Petersburg University, Saint Petersburg, Russia, 2008. (In Russian). [Google Scholar]
- Zweifach, B.W. Micropressure measurements in the terminal lymphatics. Bibl. Anat. 1973, 12, 361–365. [Google Scholar]
- Mynard, J.P.; Davidson, M.R.; Penny, D.J.; Smolich, J.J. A simple, versatile valve model for use in lumped parameter and one-dimensional cardiovascular models. Int. J. Numer. Methods Biomed. Eng. 2011, 28, 626–641. [Google Scholar] [CrossRef]
- Simakov, S.; Gamilov, T.; Soe, Y.N. Computational study of blood flow in lower extremities under intense physical load. Russ. J. Numer. Anal. Math. Model. 2013, 28. [Google Scholar] [CrossRef]
- Mozokhina, A.S.; Mukhin, S.I. Quasi-One-Dimensional Flow of a Fluid with Anisotropic Viscosity in a Pulsating Vessel. Differ. Equ. 2018, 54, 938–944. [Google Scholar] [CrossRef]
- Mozokhina, A.; Mukhin, S.; Koshelev, V. Quasi-Onedimensional Approach for Modeling the Lymph Flow in the Lymphatic System; MAKS Press: Moscow, Russia, 2017; p. 20. (In Russian) [Google Scholar]
- Jamalian, S.; Bertram, C.D.; Richardson, W.J.; Moore, J.E. Parameter sensitivity analysis of a lumped-parameter model of a chain of lymphangions in series. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H1709–H1717. [Google Scholar] [CrossRef] [PubMed]
- Quick, C.M.; Ngo, B.L.; Venugopal, A.M.; Stewart, R.H. Lymphatic pump-conduit duality: Contraction of postnodal lymphatic vessels inhibits passive flow. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H662–H668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vassilevski, Y.V.; Salamatova, V.Y.; Simakov, S.S. On the elasticity of blood vessels in one-dimensional problems of hemodynamics. Comput. Math. Math. Phys. 2015, 55, 1567–1578. [Google Scholar] [CrossRef]
- Bertram, C.; Macaskill, C.; Moore, J. Pump function curve shape for a model lymphatic vessel. Med. Eng. Phys. 2016, 38, 656–663. [Google Scholar] [CrossRef] [Green Version]
- Absi, R. Revisiting the pressure-area relation for the flow in elastic tubes: Application to arterial vessels. Ser. Biomech. 2018, 32, 47–59. [Google Scholar]
- Macdonald, A.J. The Computational Modelling of Collecting Lymphatic Vessels. Ph.D. Thesis, University of Exeter, England, UK, 2008. [Google Scholar]
- Mozokhina, A.; Lobov, G. Simulation of lymph flow with consideration of natural gravity force influence. ITM Web Conf. 2020, 31, 01003. [Google Scholar] [CrossRef] [Green Version]
- Reddy, N.; Patel, K. A mathematical model of flow through the terminal lymphatics. Med. Eng. Phys. 1995, 17, 134–140. [Google Scholar] [CrossRef]
- Mozokhina, A.S.; Mukhin, S.I.; Lobov, G.I. Pump efficiency of lymphatic vessels: Numeric estimation. Russ. J. Numer. Anal. Math. Model. 2019, 34, 261–268. [Google Scholar] [CrossRef]
- Franzone, P.C.; Pavarino, L.F.; Scacchi, S. Mathematical Cardiac Electrophysiology; Springer International Publishing: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Tretyakova, R.; Savinkov, R.; Lobov, G.; Bocharov, G. Developing Computational Geometry and Network Graph Models of Human Lymphatic System. Computation 2017, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Plasticboy. Plasticboy Pictures 2009 CC. Available online: http://www.plasticboy.co.uk/store/Human_Lymphatic_System_no_textures.html (accessed on 21 April 2020).
- Mendoza, E.; Schmid-Schonbein, G.W. A Model for Mechanics of Primary Lymphatic Valves. J. Biomech. Eng. 2003, 125, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Heppell, C.; Richardson, G.; Roose, T. A Model for Fluid Drainage by the Lymphatic System. Bull. Math. Biol. 2012, 75, 49–81. [Google Scholar] [CrossRef] [PubMed]
- Galie, P.; Spilker, R.L. A Two-Dimensional Computational Model of Lymph Transport Across Primary Lymphatic Valves. J. Biomech. Eng. 2009, 131. [Google Scholar] [CrossRef]
- Novkovic, M.; Onder, L.; Cheng, H.W.; Bocharov, G.; Ludewig, B. Integrative Computational Modeling of the Lymph Node Stromal Cell Landscape. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Mueller, S.N.; Germain, R.N. Stromal cell contributions to the homeostasis and functionality of the immune system. Nat. Rev. Immunol. 2009, 9, 618–629. [Google Scholar] [CrossRef]
- Turley, S.J.; Fletcher, A.L.; Elpek, K.G. The stromal and haematopoietic antigen-presenting cells that reside in secondary lymphoid organs. Nat. Rev. Immunol. 2010, 10, 813–825. [Google Scholar] [CrossRef]
- Link, A.; Vogt, T.K.; Favre, S.; Britschgi, M.R.; Acha-Orbea, H.; Hinz, B.; Cyster, J.G.; Luther, S.A. Fibroblastic reticular cells in lymph nodes regulate the homeostasis of naive T cells. Nat. Immunol. 2007, 8, 1255–1265. [Google Scholar] [CrossRef]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002. [Google Scholar]
- Bachmann, M.F.; Oxenius, A. Interleukin 2: From immunostimulation to immunoregulation and back again. EMBO Rep. 2007, 8, 1142–1148. [Google Scholar] [CrossRef] [Green Version]
- Hochrein, H.; Shortman, K.; Vremec, D.; Scott, B.; Hertzog, P.; O’Keeffe, M. Differential Production of IL-12, IFN-α, and IFN-γ by Mouse Dendritic Cell Subsets. J. Immunol. 2001, 166, 5448–5455. [Google Scholar] [CrossRef] [Green Version]
- Cooper, L.J.; Heppell, J.P.; Clough, G.F.; Ganapathisubramani, B.; Roose, T. An Image-Based Model of Fluid Flow Through Lymph Nodes. Bull. Math. Biol. 2015, 78, 52–71. [Google Scholar] [CrossRef] [PubMed]
- Jafarnejad, M.; Woodruff, M.C.; Zawieja, D.C.; Carroll, M.C.; Moore, J. Modeling Lymph Flow and Fluid Exchange with Blood Vessels in Lymph Nodes. Lymphat. Res. Biol. 2015, 13, 234–247. [Google Scholar] [CrossRef] [Green Version]
- Kislitsyn, A.; Savinkov, R.; Novkovic, M.; Onder, L.; Bocharov, G. Computational Approach to 3D Modeling of the Lymph Node Geometry. Computation 2015, 3, 222–234. [Google Scholar] [CrossRef]
- Savinkov, R.; Kislitsyn, A.; Watson, D.J.; van Loon, R.; Sazonov, I.; Novkovic, M.; Onder, L.; Bocharov, G. Data-driven modelling of the FRC network for studying the fluid flow in the conduit system. Eng. Appl. Artif. Intell. 2017, 62, 341–349. [Google Scholar] [CrossRef] [Green Version]
- Donovan, G.M.; Lythe, G. T cell and reticular network co-dependence in HIV infection. J. Theor. Biol. 2016, 395, 211–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pepper, M.S.; Tille, J.C.; Nisato, R.; Skobe, M. Lymphangiogenesis and tumor metastasis. Cell Tissue Res. 2003, 314, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Eymard, N.; Volpert, V.; Quere, I.; Lajoinie, A.; Nony, P.; Cornu, C. A 2D Computational Model of Lymphedema and of its Management with Compression Device. Math. Model. Nat. Phenom. 2017, 12, 180–195. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mozokhina, A.; Savinkov, R. Mathematical Modelling of the Structure and Function of the Lymphatic System. Mathematics 2020, 8, 1467. https://doi.org/10.3390/math8091467
Mozokhina A, Savinkov R. Mathematical Modelling of the Structure and Function of the Lymphatic System. Mathematics. 2020; 8(9):1467. https://doi.org/10.3390/math8091467
Chicago/Turabian StyleMozokhina, Anastasia, and Rostislav Savinkov. 2020. "Mathematical Modelling of the Structure and Function of the Lymphatic System" Mathematics 8, no. 9: 1467. https://doi.org/10.3390/math8091467
APA StyleMozokhina, A., & Savinkov, R. (2020). Mathematical Modelling of the Structure and Function of the Lymphatic System. Mathematics, 8(9), 1467. https://doi.org/10.3390/math8091467