
The Usefulness of Autoradiography for DNA Repair Proteins Activity Detection in the Cytoplasm towards Radiolabeled Oligonucleotides Containing 5′,8-Cyclo-2′-deoxyadenosine



Abstract
:1. Introduction
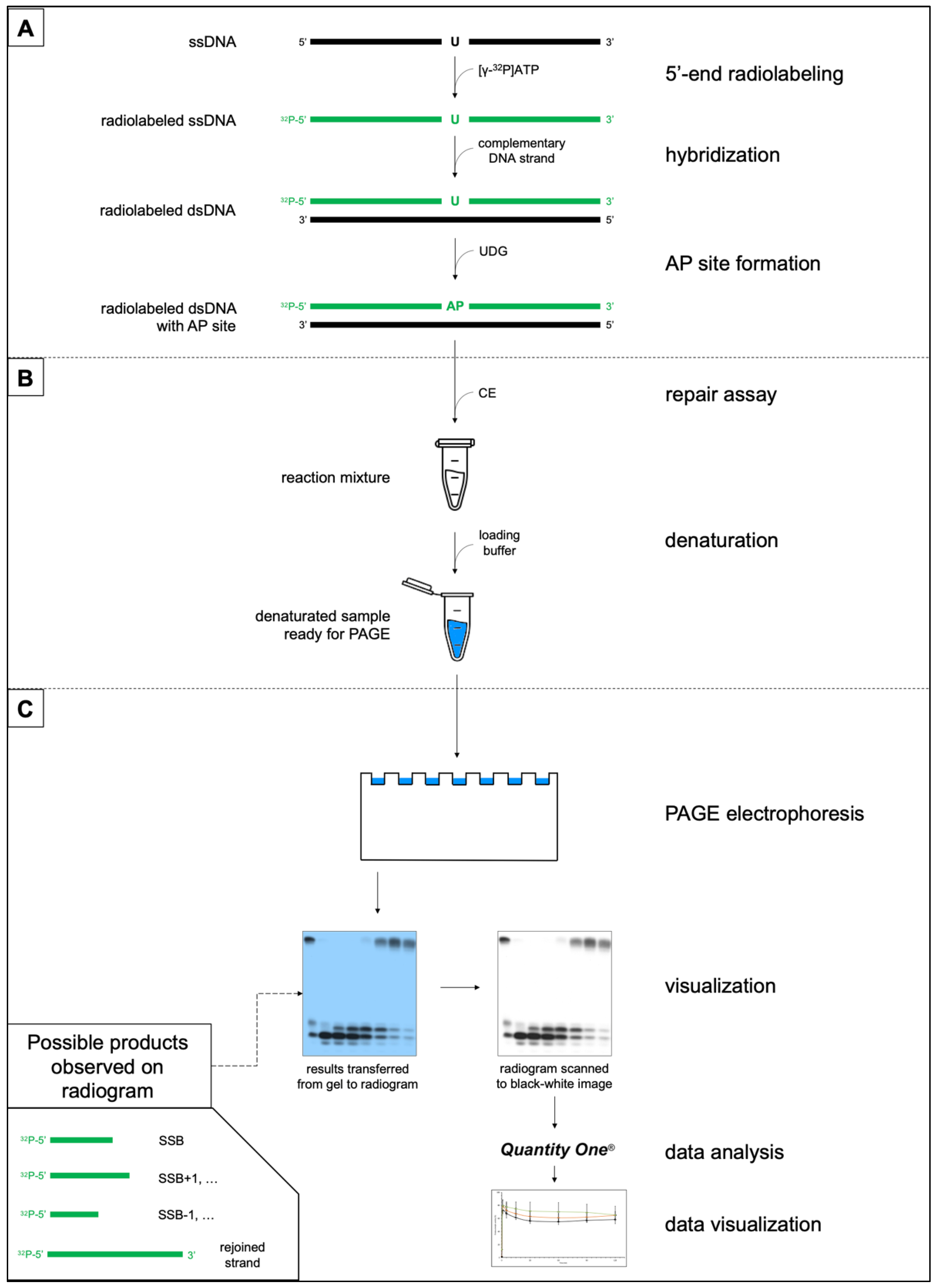
2. Materials and Methods
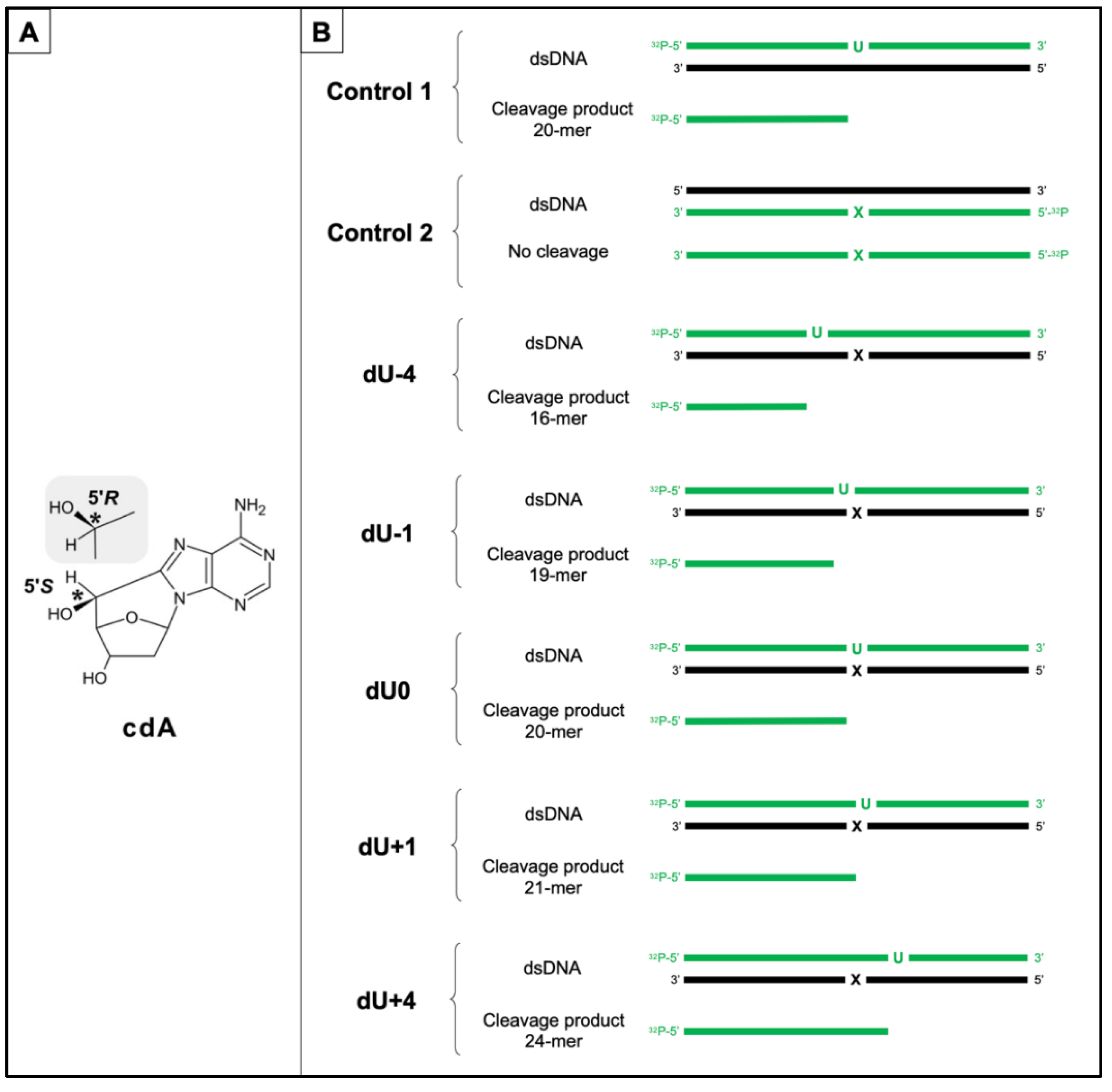
2.1. Substrate Oligonucleotides-Containing AP Site
2.2. Cell Cultures
2.3. Preparation of Cytoplasmic Extracts
2.4. Repair Assays
3. Results
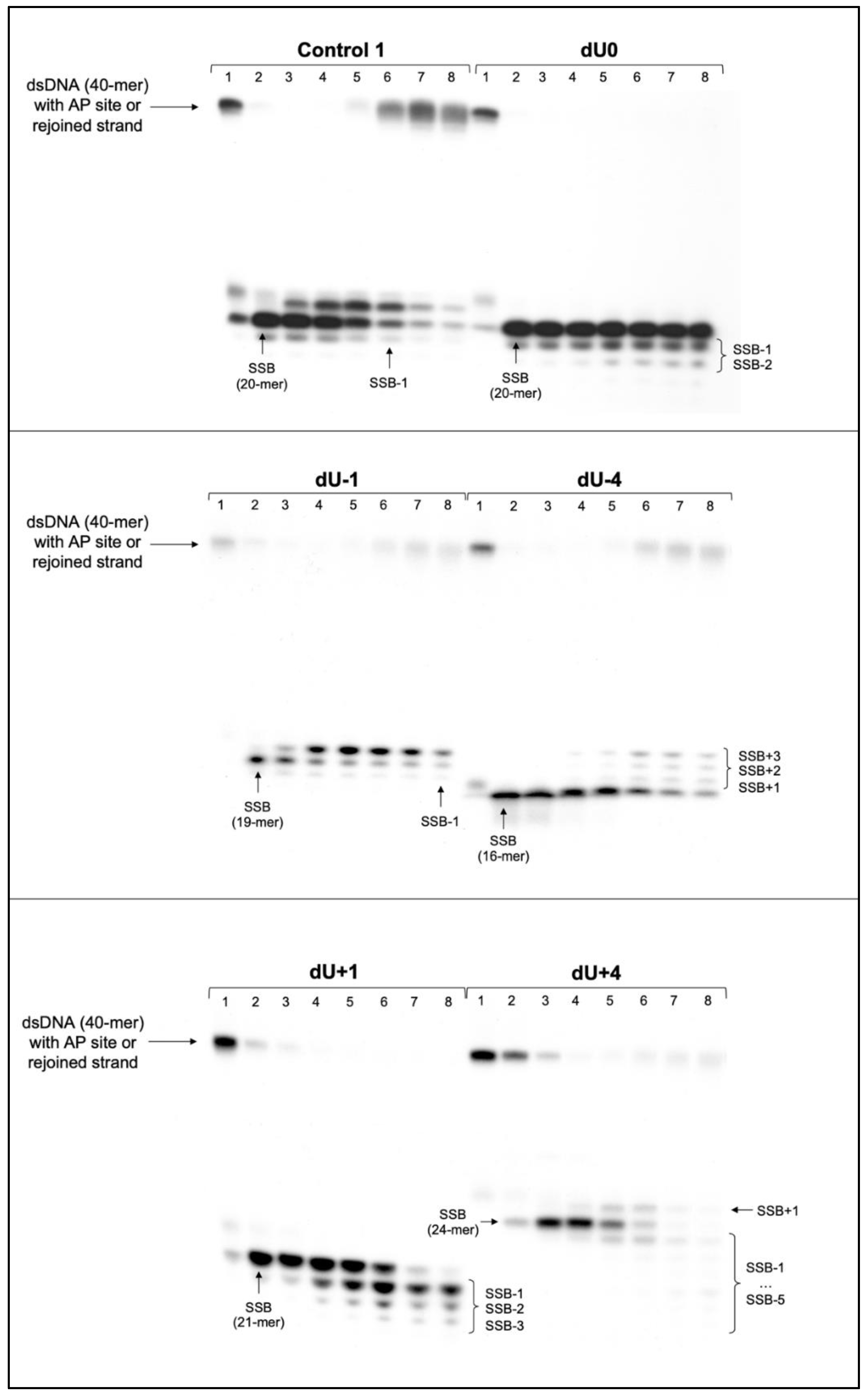
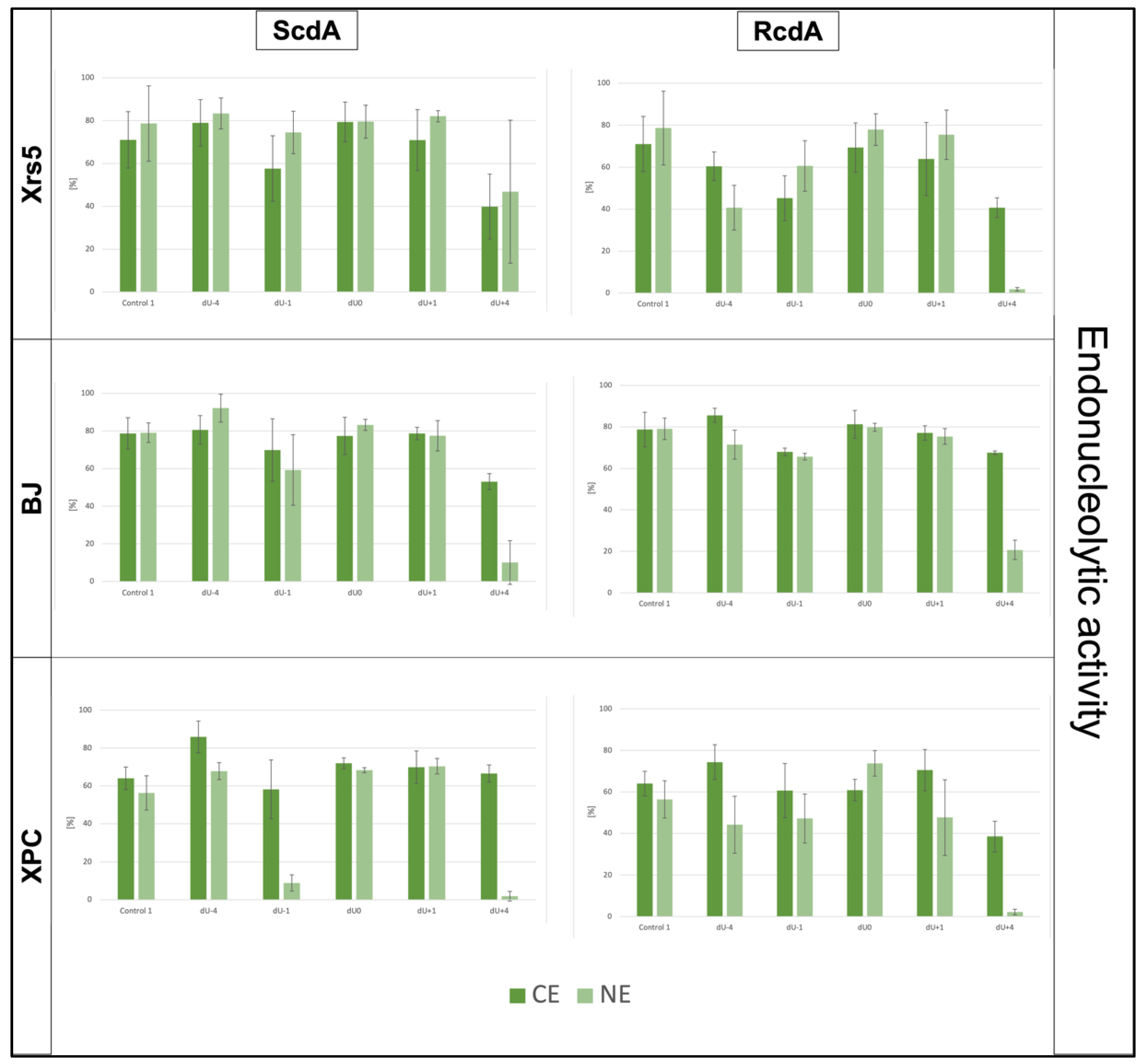
3.1. The DNA Strand Incision—Endonucleolytic Activity of Cytoplasmic Extracts
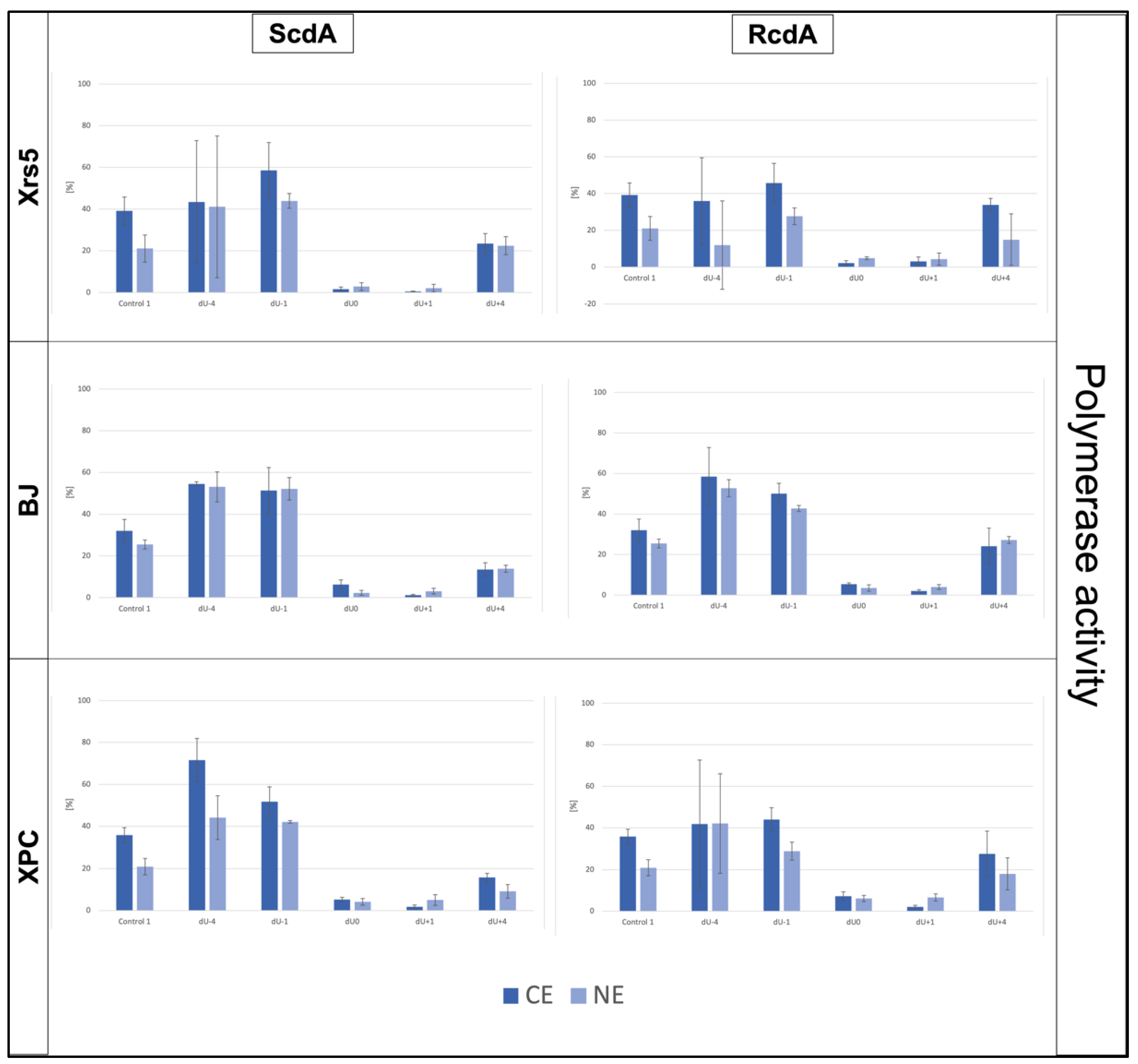
3.2. The DNA Strand Elongation—Polymerase Activity of Cytoplasmic Extracts
3.3. The DNA Strand Degradation—Exonucleolytic Activity of Cytoplasmic Extracts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bayley, S.T. Autoradiography of Single Cells. Nature 1947, 160, 193–194. [Google Scholar] [CrossRef]
- Eccles, L.J.; Lomax, M.E.; O’Neill, P. Hierarchy of Lesion Processing Governs the Repair, Double-Strand Break Formation and Mutability of Three-Lesion Clustered DNA Damage. Nucleic Acids Res. 2009, 38, 1123–1134. [Google Scholar] [CrossRef] [Green Version]
- Mourgues, S.; Lomax, M.E.; O’Neill, P. Base Excision Repair Processing of Abasic Site/Single-Strand Break Lesions within Clustered Damage Sites Associated with XRCC1 Deficiency. Nucleic Acids Res. 2007, 35, 7676–7687. [Google Scholar] [CrossRef] [Green Version]
- Lomax, M.E.; Cunniffe, S.; O’Neill, P. Efficiency of Repair of an Abasic Site within DNA Clustered Damage Sites by Mammalian Cell Nuclear Extracts. Biochemistry 2004, 43, 11017–11026. [Google Scholar] [CrossRef]
- Eccles, L.J.; Menoni, H.; Angelov, D.; Lomax, M.E.; O’Neill, P. Efficient Cleavage of Single and Clustered AP Site Lesions within Mono-Nucleosome Templates by CHO-K1 Nuclear Extract Contrasts with Retardation of Incision by Purified APE1. DNA Repair 2015, 35, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Karwowski, B.T. (5′S) 5′,8-Cyclo-2′-Deoxyadenosine Cannot Stop BER. Clustered DNA Lesion Studies. Int. J. Mol. Sci. 2021, 22, 5934. [Google Scholar] [CrossRef]
- Boguszewska, K.; Kaźmierczak-Barańska, J.; Karwowski, B.T. The Influence of 5′,8-Cyclo-2′-Deoxypurines on the Mitochondrial Repair of Clustered Dna Damage in Xrs5 Cells: The Preliminary Study. Molecules 2021, 26, 7042. [Google Scholar] [CrossRef]
- Boguszewska, K.; Szewczuk, M.; Kaźmierczak-Barańska, J.; Karwowski, B.T. How (5′S) and (5′R) 5′,8-Cyclo-2′-Deoxypurines Affect Base Excision Repair of Clustered DNA Damage in Nuclear Extracts of Xrs5 Cells? A Biochemical Study. Cells 2021, 10, 725. [Google Scholar] [CrossRef]
- Karwowski, B.; Czarny, P.; Szemraj, J.; Śliwiński, T.; Majchrzak, K.; Merecz-Sadowska, A.; Jabłkowski, M. The Influence of Hepatitis C Virus Therapy on the DNA Base Excision Repair System of Peripheral Blood Mononuclear Cells. DNA Cell Biol. 2017, 36, 535–540. [Google Scholar] [CrossRef]
- Karwowski, B.T.; Bellon, S.; O’Neill, P.; Lomax, M.E.; Cadet, J. Effects of (5′S)-5′,8-Cyclo-2′-Deoxyadenosine on the Base Excision Repair of Oxidatively Generated Clustered DNA Damage. A Biochemical and Theoretical Study. Org. Biomol. Chem. 2014, 12, 8671–8682. [Google Scholar] [CrossRef]
- Bundy, D. Autoradiography. In Current Protocols in Protein Science; John Wiley & Sons Inc.: Hoboken, NJ, USA, 1997; pp. 10.11.1–10.11.6. [Google Scholar]
- Voytas, D.; Ke, N. Detection and Quantitation of Radiolabeled Proteins and DNA in Gels and Blots. In Current Protocols in Immunology; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2002; pp. A.31.1–A.31.10. [Google Scholar]
- Lomax, M.E.; Folkes, L.K.; O’Neill, P. Biological Consequences of Radiation-Induced DNA Damage: Relevance to Radiotherapy. Clin. Oncol. 2013, 25, 578–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkart, W.; Jung, T.; Frash, G. Damage Pattern as a Function of Radiation Quality and Other Factors. Comptes Rendus l’Académie Sci. 1999, 322, 89–101. [Google Scholar] [CrossRef]
- Cadet, J.; Ravanat, J.-L.; TavernaPorro, M.; Menoni, H.; Angelov, D. Oxidatively Generated Complex DNA Damage: Tandem and Clustered Lesions. Cancer Lett. 2012, 327, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Karwowski, B.T. The Influence of Single, Tandem, and Clustered DNA Damage on the Electronic Properties of the Double Helix: A Theoretical Study. Molecules 2020, 25, 3126. [Google Scholar] [CrossRef]
- Chatgilialoglu, C.; Ferreri, C.; Geacintov, N.E.; Krokidis, M.G.; Liu, Y.; Masi, A.; Shafirovich, V.; Terzidis, M.A.; Tsegay, P.S. 5′,8-Cyclopurine Lesions in DNA Damage: Chemical, Analytical, Biological, and Diagnostic Significance. Cells 2019, 8, 513. [Google Scholar] [CrossRef] [Green Version]
- Pande, P.; Das, R.S.; Sheppard, C.; Kow, Y.W.; Basu, A.K. Repair Efficiency of (5′S)-8,5′-Cyclo-2′-Deoxyguanosine and (5′S)-8,5′-Cyclo-2′-Deoxyadenosine Depends on the Complementary Base. DNA Repair 2012, 11, 926–931. [Google Scholar] [CrossRef] [Green Version]
- Karwowski, B.T. The Influence of (5′R)- and (5′S)-5′,8-Cyclo-2′-Deoxyadenosine on UDG and HAPE1 Activity. Tandem Lesions Are the Base Excision Repair System’s Nightmare. Cells 2019, 8, 1303. [Google Scholar] [CrossRef] [Green Version]
- Jaruga, P.; Rozalski, R.; Jawien, A.; Migdalski, A.; Olinski, R.; Dizdaroglu, M. DNA Damage Products (5′R)- and (5′S)-8,5′-Cyclo-2′-Deoxyadenosines as Potential Biomarkers in Human Urine for Atherosclerosis. Biochemistry 2012, 51, 1822–1824. [Google Scholar] [CrossRef]
- Mori, T.; Nakane, H.; Iwamoto, T.; Krokidis, M.G.; Chatgilialoglu, C.; Tanaka, K.; Kaidoh, T.; Hasegawa, M.; Sugiura, S. High Levels of Oxidatively Generated DNA Damage 8,5′-Cyclo-2′-Deoxyadenosine Accumulate in the Brain Tissues of Xeroderma Pigmentosum Group A Gene-Knockout Mice. DNA Repair 2019, 80, 52–58. [Google Scholar] [CrossRef]
- Jaruga, P.; Theruvathu, J.; Dizdaroglu, M.; Brooks, P.J. Complete Release of (5′S)-8,5′-Cyclo-2′-Deoxyadenosine from Dinucleotides, Oligodeoxynucleotides and DNA, and Direct Comparison of Its Levels in Cellular DNA with Other Oxidatively Induced DNA Lesions. Nucleic Acids Res. 2004, 32, e87. [Google Scholar] [CrossRef] [Green Version]
- Pednekar, V.; Weerasooriya, S.; Jasti, V.P.; Basu, A.K. Mutagenicity and Genotoxicity of (5′S)-8,5′-Cyclo-2′-Deoxyadenosine in Escherichia coli and Replication of (5′S)-8,5′-Cyclopurine-2′-Deoxynucleosides in Vitro by Dna Polymerase Iv, Exo-Free Klenow Fragment, and Dpo4. Chem. Res. Toxicol. 2014, 27, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Jaruga, P.; Birincioglu, M.; Rodriguez, H.; Dizdaroglu, M. Mass Spectrometric Assays for the Tandem Lesion 8,5′-Cyclo-2′-Deoxyguanosine in Mammalian DNA. Biochemistry 2002, 41, 3703–3711. [Google Scholar] [CrossRef] [PubMed]
- Kuraoka, I.; Bender, C.; Romieu, A.; Cadet, J.; Wood, R.D.; Lindahl, T. Removal of Oxygen Free-Radical-Induced 5′,8-Purine Cyclodeoxynucleosides from DNA by the Nucleotide Excision-Repair Pathway in Human Cells. Proc. Natl. Acad. Sci. USA 2000, 97, 3832–3837. [Google Scholar] [CrossRef] [Green Version]
- Bukowska, B.; Karwowski, B.T. The Clustered DNA Lesions—Types, Pathways of Repair and Relevance to Human Health. Curr. Med. Chem. 2018, 25, 2722–2735. [Google Scholar] [CrossRef] [PubMed]
- Brooks, P.J. The Cyclopurine Deoxynucleosides: DNA Repair, Biological Effects, Mechanistic Insights, and Unanswered Questions. Free Radic. Biol. Med. 2017, 107, 90–100. [Google Scholar] [CrossRef] [PubMed]
- David-Cordonnier, M.H.; Cunniffe, S.M.T.; Hickson, I.D.; O’Neill, P. Efficiency of Incision of an AP Site within Clustered DNA Damage by the Major Human AP Endonuclease. Biochemistry 2002, 41, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Merecz, A.; Karwowski, B.T. DNA Tandem Lesion: 5′,8-Cyclo-2′-Deoxyadenosine. The Influence on Human Health. Mol. Biol. 2016, 50, 793–798. [Google Scholar] [CrossRef]
- Kaźmierczak-Barańska, J.; Boguszewska, K.; Szewczuk, M.; Karwowski, B.T. Effects of 5′,8′-Cyclo-2′-Deoxypurines on the Base Excisionrepair of Clustered DNA Lesions in Nuclear Extracts of the Xpc Cell Line. Cells 2021, 10, 3254. [Google Scholar] [CrossRef]
- Szewczuk, M.; Boguszewska, K.; Kaźmierczak-Barańska, J.; Karwowski, B.T. The Influence of 5′R and 5′S Cda and Cdg on the Activity of Bsmai and Sspi Restriction Enzymes. Molecules 2021, 26, 3750. [Google Scholar] [CrossRef]
- Szewczuk, M.; Boguszewska, K.; Kaźmierczak-Barańska, J.; Karwowski, B.T. When Udg and Hape1 Meet Cyclopurines. How (5′R) and (5′S) 5′,8-Cyclo-2′-Deoxyadenosine and 5′,8-Cyclo-2′-Deoxyguanosine Affect Udg and Hape1 Activity? Molecules 2021, 26, 5177. [Google Scholar] [CrossRef]
- Szewczuk, M.; Karwowski, B. The Influence of CdG on 8-OxodG Excision by OGG1 and FPG Glycosylases. Acta Biochim. Pol. 2022, 69, 227–232. [Google Scholar] [CrossRef]
- Jaruga, P.; Dizdaroglu, M. 8,5′-Cyclopurine-2′-Deoxynucleosides in DNA: Mechanisms of Formation, Measurement, Repair and Biological Effects. DNA Repair 2008, 7, 1413–1425. [Google Scholar] [CrossRef] [PubMed]
- Brooks, P.J. The 8,5′-Cyclopurine-2′-Deoxynucleosides: Candidate Neurodegenerative DNA Lesions in Xeroderma Pigmentosum, and Unique Probes of Transcription and Nucleotide Excision Repair. DNA Repair 2008, 7, 1168–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraemer, K.H.; Patronas, N.J.; Schiffmann, R.; Brooks, B.P.; Tamura, D.; DiGiovanna, J.J. Xeroderma Pigmentosum, Trichodystrophy and Cockayne Syndrome: A Complex Genotype-Phenotype Relationship. Neuroscience 2007, 145, 1388–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fayyad, N.; Kobaisi, F.; Beal, D.; Mahfouf, W.; Ged, C.; Morice-Picard, F.; Fayyad-Kazan, M.; Fayyad-Kazan, H.; Badran, B.; Rezvani, H.R.; et al. Xeroderma Pigmentosum C (XPC) Mutations in Primary Fibroblasts Impair Base Excision Repair Pathway and Increase Oxidative DNA Damage. Front. Genet. 2020, 11, 1458. [Google Scholar] [CrossRef]
- Shivji, M.K.K.; Eker, A.P.M.; Wood, R.D. DNA Repair Defect in Xeroderma Pigmentosum Group C and Complementing Factor from HeLa Cells. J. Biol. Chem. 1994, 269, 22749–22757. [Google Scholar] [CrossRef]
- Gong, L.; Zhang, Y.; Liu, C.; Zhang, M.; Han, S. Application of Radiosensitizers in Cancer Radiotherapy. Int. J. Nanomed. 2021, 16, 1083–1102. [Google Scholar] [CrossRef]
- Herath, N.I.; Berthault, N.; Thierry, S.; Jdey, W.; Lienafa, M.C.; Bono, F.; Noguiez-Hellin, P.; Sun, J.S.; Dutreix, M. Preclinical Studies Comparing Efficacy and Toxicity of DNA Repair Inhibitors, Olaparib, and AsiDNA, in the Treatment of Carboplatin-Resistant Tumors. Front. Oncol. 2019, 9, 1097. [Google Scholar] [CrossRef] [Green Version]
- Jdey, W.; Thierry, S.; Russo, C.; Devun, F.; Al Abo, M.; Noguiez-Hellin, P.; Sun, J.S.; Barillot, E.; Zinovyev, A.; Kuperstein, I.; et al. Drug-Driven Synthetic Lethality: Bypassing Tumor Cell Genetics with a Combination of AsiDNA and PARP Inhibitors. Clin. Cancer Res. 2017, 23, 1001–1011. [Google Scholar] [CrossRef] [Green Version]
- Gökirmak, T.; Nikan, M.; Wiechmann, S.; Prakash, T.P.; Tanowitz, M.; Seth, P.P. Overcoming the Challenges of Tissue Delivery for Oligonucleotide Therapeutics. Trends Pharmacol. Sci. 2021, 42, 588–604. [Google Scholar] [CrossRef]
- Hammond, S.M.; Aartsma-Rus, A.; Alves, S.; Borgos, S.E.; Buijsen, R.A.M.; Collin, R.W.J.; Covello, G.; Denti, M.A.; Desviat, L.R.; Echevarría, L.; et al. Delivery of Oligonucleotide-based Therapeutics: Challenges and Opportunities. EMBO Mol. Med. 2021, 13, e13243. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.; Foray, C.; Gatto, A.; Larcher, M.; Heinrich, S.; Lupu, M.; Mispelter, J.; Boussin, F.D.; Pouponnot, C.; Dutreix, M. AsiDNA Is a Radiosensitizer with No Added Toxicity in Medulloblastoma Pediatric Models. Clin. Cancer Res. 2021, 26, 5735–5746. [Google Scholar] [CrossRef] [PubMed]
- Johannes, L.; Lucchino, M. Current Challenges in Delivery and Cytosolic Translocation of Therapeutic RNAs. Nucleic Acid Ther. 2018, 28, 178–193. [Google Scholar] [CrossRef] [PubMed]
- Topf, U.; Wrobel, L.; Chacinska, A. Chatty Mitochondria: Keeping Balance in Cellular Protein Homeostasis. Trends Cell Biol. 2016, 26, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Chacinska, A.; Rehling, P. Moving Proteins from the Cytosol Intomitochondria. Biochem. Soc. Trans. 2004, 32, 774–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karwowski, B.T.; Grand, A.; Cadet, J. 5′,8-Cyclo-2′-Deoxyadenosine (CdA) Formation by γ-Radiation. Theoretical Quantum Mechanics Study. Acta Biochim. Pol. 2009, 56, 655–662. [Google Scholar] [CrossRef]
- Karwowski, B.T. Formation of 5′,8-Cyclo-2′-Deoxyadenosine in Single Strand DNA. Theoretical Quantum Mechanics Study. Org. Biomol. Chem. 2010, 8, 1603. [Google Scholar] [CrossRef]
- Guerrero, C.R.; Wang, J.; Wang, Y. Induction of 8,5′-Cyclo-2′-Deoxyadenosine and 8,5′-Cyclo-2′-Deoxyguanosine in Isolated DNA by Fenton-Type Reagents. Chem. Res. Toxicol. 2013, 26, 1361–1366. [Google Scholar] [CrossRef] [Green Version]
- Kaniak-Golik, A.; Skoneczna, A. Mitochondria-Nucleus Network for Genome Stability. Free. Radic. Biol. Med. 2015, 82, 73–104. [Google Scholar] [CrossRef] [Green Version]
- Shaughnessy, D.T.; McAllister, K.; Worth, L.; Haugen, A.C.; Meyer, J.N.; Domann, F.E.; van Houten, B.; Mostoslavsky, R.; Bultman, S.J.; Baccarelli, A.A.; et al. Mitochondria, Energetics, Epigenetics, and Cellular Responses to Stress. Environ. Health Perspect. 2015, 122, 1271–1278. [Google Scholar] [CrossRef]
- Izumi, T.; Mellon, I. Base Excision Repair and Nucleotide Excision Repair; Elsevier Inc.: Amsterdam, The Netherlands, 2016; ISBN 9780128033456. [Google Scholar]
- Prakash, A.; Doublie, S. Base Excision Repair in the Mitochondria. J. Cell Biochem. 2015, 116, 1490–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frosina, G.; Fortini, P.; Rossi, O.; Carrozzino, F.; Raspaglio, G.; Cox, L.S.; Lane, D.P.; Abbondandolo, A.; Dogliotti, E. Two Pathways for Base Excision Repair in Mammalian Cells. J. Biol. Chem. 1996, 271, 9573–9578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romieu, A.; Gasparutto, D.; Cadet, J. Synthesis and Characterization of Oligonucleotides Containing 5′,8- Cyclopurine 2′-Deoxyribonucleosides: (5′R)-5′,8-Cyclo-2′-Deoxyadenosine, (5′S)-5′,8-Cyclo-2′-Deoxyguanosine, and (5′R)-5′,8-Cyclo-2′-Deoxyguanosine. Chem. Res. Toxicol. 1999, 12, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Walker, G.; Chatterjee, N. Mechanisms of DNA Damage, Repair, and Mutagenesis. Environ. Mol. Mutagenesis 2017, 58, 235–263. [Google Scholar] [CrossRef]
- Vilenchik, M.M.; Knudson, A.G. Endogenous DNA Double-Strand Breaks: Production, Fidelity of Repair, and Induction of Cancer. Proc. Natl. Acad. Sci. USA 2003, 100, 12871–12876. [Google Scholar] [CrossRef] [Green Version]
- Tubbs, A.; Nussenzweig, A. Endogenous DNA Damage as a Source of Genomic Instability in Cancer. Cell 2017, 168, 644–656. [Google Scholar] [CrossRef] [Green Version]
- Jang, B.; Kwon, H.; Katila, P.; Lee, S.J.; Lee, H. Dual Delivery of Biological Therapeutics for Multimodal and Synergistic Cancer Therapies. Adv. Drug Deliv. Rev. 2016, 98, 113–133. [Google Scholar] [CrossRef]
- Takakura, K.; Kawamura, A.; Torisu, Y.; Koido, S.; Yahagi, N.; Saruta, M. The Clinical Potential of Oligonucleotide Therapeutics against Pancreatic Cancer. Int. J. Mol. Sci. 2019, 20, 3331. [Google Scholar] [CrossRef] [Green Version]
- Lundin, K.E.; Gissberg, O.; Smith, C.I.E. Oligonucleotide Therapies: The Past and the Present. Hum. Gene Ther. 2015, 26, 475–485. [Google Scholar] [CrossRef] [Green Version]
- Roberts, T.C.; Langer, R.; Wood, M.J.A. Advances in Oligonucleotide Drug Delivery. Nat. Rev. Drug Discov. 2020, 19, 673–694. [Google Scholar] [CrossRef]
- Jasti, V.P.; Das, R.S.; Hilton, B.A.; Weerasooriya, S.; Zou, Y.; Basu, A.K. (5′S)-8,5′-Cyclo-2′-Deoxyguanosine Is a Strong Block to Replication, a Potent Pol V-Dependent Mutagenic Lesion, and Is Inefficiently Repaired in Escherichia coli. Biochemistry 2011, 50, 3862–3865. [Google Scholar] [CrossRef] [PubMed]
- Krokan, H.E.; Bjørås, M. Base Excision Repair. Cold Spring Harb. Perspect. Biol. 2013, 5, a012583. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| xrs5 [%] | BJ [%] | XPC [%] | |
|---|---|---|---|
| Endonucleolytic activity (1 min) | 71.0 | 78.73 | 64.01 |
| Polymerase activity (30 min) | 39.19 | 31.98 | 35.82 |
| Exonucleolytic activity (120 min) | 2.38 | 4.69 | 8.89 |
| Strand Incision | xrs5 | BJ | XPC |
|---|---|---|---|
| CE | +4 < −1 < +1 < −4 < 0 (ScdA) +4 < −1 < −4 < +1 < 0 (RcdA) | +4 < −1 < 0 < +1 < −4 (ScdA) +4 ~ −1 < +1 < 0 < −4 (RcdA) | −1 < +4 < +1 < 0 < −4 (ScdA) +4 < −1 ~ 0 < +1 < −4 (RcdA) |
| NE | +4 < −1 < 0 < +1 < −4 (ScdA) +4 < −4 < −1 < +1 < 0 (RcdA) | +4 < −1 < +1 < 0 < −4 (ScdA) +4 < −1 < −4 < +1 < 0 (RcdA) | +4 < −1 < −4 < 0 < +1 (ScdA) +4 < −1 < −4 < +1 < 0 (RcdA) |
| Strand Elongation | xrs5 | BJ | XPC |
|---|---|---|---|
| CE | +4 < −4 < −1 (ScdA) +4 < −4 < −1 (RcdA) | +4 < −1 < −4 (ScdA) +4 < −1 < −4 (RcdA) | +4 < −1 < −4 (ScdA) +4 < −4 < −1 (RcdA) |
| NE | +4 < −4 < −1 (ScdA) −4 < +4 < −1 (RcdA) | +4 < −1 < −4 (ScdA) +4 < −1 < −4 (RcdA) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boguszewska, K.; Karwowski, B.T.; Kaźmierczak-Barańska, J. The Usefulness of Autoradiography for DNA Repair Proteins Activity Detection in the Cytoplasm towards Radiolabeled Oligonucleotides Containing 5′,8-Cyclo-2′-deoxyadenosine. Chemosensors 2022, 10, 204. https://doi.org/10.3390/chemosensors10060204
Boguszewska K, Karwowski BT, Kaźmierczak-Barańska J. The Usefulness of Autoradiography for DNA Repair Proteins Activity Detection in the Cytoplasm towards Radiolabeled Oligonucleotides Containing 5′,8-Cyclo-2′-deoxyadenosine. Chemosensors. 2022; 10(6):204. https://doi.org/10.3390/chemosensors10060204
Chicago/Turabian StyleBoguszewska, Karolina, Bolesław T. Karwowski, and Julia Kaźmierczak-Barańska. 2022. "The Usefulness of Autoradiography for DNA Repair Proteins Activity Detection in the Cytoplasm towards Radiolabeled Oligonucleotides Containing 5′,8-Cyclo-2′-deoxyadenosine" Chemosensors 10, no. 6: 204. https://doi.org/10.3390/chemosensors10060204
APA StyleBoguszewska, K., Karwowski, B. T., & Kaźmierczak-Barańska, J. (2022). The Usefulness of Autoradiography for DNA Repair Proteins Activity Detection in the Cytoplasm towards Radiolabeled Oligonucleotides Containing 5′,8-Cyclo-2′-deoxyadenosine. Chemosensors, 10(6), 204. https://doi.org/10.3390/chemosensors10060204