
Kinetics of Odorant Recognition with a Graphene-Based Olfactory Receptor Mimicry


Abstract
:1. Introduction
2. Materials and Methods
- TcasOBP9A: AMSEAQLKAAVKLVRNMCQPKSKATNEDIEKMHHGDWNIDRTAMCYMHCALNSNKLITKENV
- TcasOBP9B: AMSEAQMKAALKLVRNVCQPKTKATNEQIEAMHTGNWDLDKNGKCYMWCILNMYKLIGKDNS
- TcasOBP9A: FNRDYAITLAEKNLPTALKTASIEAANLCKDSAKTLDDKCVAAYEISKCLYESNPEKYFLP
- TcasOBP9B: FDWEAGIATLKAQAPESVRDPAIASVNNCKDAVKTTSDKCEAAYEIAHCMYLDNPEKYFLP
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grünig, M.; Mazzi, D.; Calanca, P.; Karger, D.N.; Pellissier, L. Crop and forest pest metawebs shift towards increased linkage and suitability overlap under climate change. Commun. Biol. 2020, 3, 233. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 361, 916–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mozūraitis, R.; Hajkazemian, M.; Zawada, J.W.; Szymczak, J.; Pålsson, K.; Sekar, V.; Biryukova, I.; Friedländer, M.R.; Koekemoer, L.L.; Baird, J.K.; et al. Male swarming aggregation pheromones increase female attraction and mating success among multiple African malaria vector mosquito species. Nat. Ecol. Evol. 2020, 4, 1395–1401. [Google Scholar] [CrossRef] [PubMed]
- McBride, C.S.; Baier, F.; Omondi, A.B.; Spitzer, S.A.; Lutomiah, J.; Sang, R.; Ignell, R.; Vosshall, L.B. Evolution of mosquito preference for humans linked to an odorant receptor. Nature 2014, 515, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Raji, J.I.; Melo, N.; Castillo, J.S.; Gonzalez, S.; Saldana, V.; Stensmyr, M.C.; DeGennaro, M. Aedes aegypti mosquitoes detect acidic volatiles found in human odor using the IR8a pathway. Curr. Biol. 2019, 29, 1253–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francke, W.; Schulz, S. 4.04-Pheromones of terrestrial invertebrates. In Comprehensive Natural Products II; Elsevier: Amsterdam, The Netherlands, 2010; pp. 153–223. [Google Scholar]
- Quiroz, A.; Pettersson, J.; Pickett, J.A.; Wadhams, L.J.; Niemeyer, H.M. Semiochemicals Mediating Spacing Behavior of Bird Cherry-Oat Aphid, Rhopalosiphum padi Feeding on Cereals. J. Chem. Ecol. 1997, 23, 2599–2607. [Google Scholar] [CrossRef]
- Aphids as Crop Pest; Van Emden, H.F.; Harrington, R. (Eds.) Cabi: Wallingford, UK, 2014. [Google Scholar]
- Jones, W.D.; Nguyen, T.A.T.; Kloss, B.; Lee, K.J.; Vosshall, L.B. Functional conservation of an insect odorant receptor gene across 250 million years of evolution. Curr. Biol. 2005, 15, R119–R121. [Google Scholar] [CrossRef] [Green Version]
- Trebels, B.; Dippel, S.; Schaaf, M.; Balakrishnan, K.; Wimmer, E.A.; Schachtner, J. Adult neurogenesis in the mushroom bodies of red flour beetles (Tribolium castaneum, Herbst) is influenced by the olfactory environment. Sci. Rep. 2020, 10, 1090. [Google Scholar] [CrossRef]
- Engsontia, P.; Sanderson, A.P.; Cobb, M.; Walden, K.K.; Robertson, H.M.; Brown, S. The red flour beetle’s large nose: An expanded odorant receptor gene family in Tribolium castaneum. Insect Biochem. Mol. Biol. 2008, 38, 387–397. [Google Scholar] [CrossRef]
- Dippel, S.; Kollmann, M.; Oberhofer, G.; Montino, A.; Knoll, C.; Krala, M.; Wimmer, E.A. Morphological and transcriptomic analysis of a beetle chemosensory system reveals a gnathal olfactory center. BMC Biol. 2016, 14, 90. [Google Scholar] [CrossRef] [Green Version]
- Larisika, M.; Kotlowski, C.; Steininger, C.; Mastrogiacomo, R.; Pelosi, P.; Schütz, S.; Peteu, S.; Kleber, C.; Reiner-Rozman, C.; Nowak, C.; et al. Electronic olfactory sensor based on A. mellifera odorant-binding protein 14 on a reduced graphene oxide field-effect transistor. Angew. Chem. Int. Ed. 2015, 54, 13245–13248. [Google Scholar] [CrossRef] [PubMed]
- Kotlowski, C.; Larisika, M.; Guerin, P.M.; Kleber, C.; Kröber, T.; Mastrogiacomo, R.; Nowak, C.; Pelosi, P.; Schütz, S.; Schwaighofer, A.; et al. Fine discrimination of volatile compounds by graphene-immobilized odorant-binding proteins. Sens. Actuators B Chem. 2018, 256, 564–572. [Google Scholar] [CrossRef]
- Daly, N.L.; Craik, D.J. Bioactive cystine knot proteins. Curr. Opin. Chem. Biol. 2011, 15, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Lustbader, J.W.; Liu, Y.; Canfield, R.E.; Hendrickson, W.A. Structure of human chorionic gonadotropin at 2.6 Å resolution from MAD analysis of the selenomethionyl protein. Structure 1994, 2, 545–558. [Google Scholar] [CrossRef] [Green Version]
- Montino, A.; Balakrishnan, K.; Dippel, S.; Trebels, B.; Neumann, P.; Wimmer, E.A. Mutually Exclusive Expression of Closely Related Odorant-Binding Proteins 9A and 9B in the Antenna of the Red Flour Beetle Tribolium castaneum. Biomolecules 2021, 11, 1502. [Google Scholar] [CrossRef]
- Damberger, F.F.; Michel, E.; Ishida, Y.; Leal, W.S.; Wüthrich, K. Pheromone discrimination by a pH-tuned polymorphism of the Bombyx mori pheromone-binding protein. Proc. Natl. Acad. Sci. USA 2013, 110, 18680–18685. [Google Scholar] [CrossRef] [Green Version]
- Kaissling, K.E. Responses of insect olfactory neurons to single pheromone molecules. In Olfactory Concepts of Insect Control-Alternative to Insecticides; Springer: Cham, Switzerland, 2019; pp. 1–27. [Google Scholar]
- Meyer, S.A.; Ozbay, B.N.; Potcoava, M.; Salcedo, E.; Restrepo, D.; Gibson, E.A. Super-resolution imaging of ciliary microdomains in isolated olfactory sensory neurons using a custom two-color stimulated emission depletion microscope. J. Biomed. Opt. 2016, 21, 066017. [Google Scholar] [CrossRef] [Green Version]
- Wiltgen, S.M.; Smith, I.F.; Parker, I. Superresolution localization of single functional IP3R channels utilizing Ca2+ flux as a readout. Biophys. J. 2010, 99, 437–446. [Google Scholar] [CrossRef] [Green Version]
- Paczkowski, S. Insect Olfaction as an Information Filter for Chemo-Analytical Applications. Doctoral Dissertation, Georg-August-University Göttingen, Göttingen, Germany, 2014. [Google Scholar]
- Hummers, W.S.; Offeman, R.E. Preparation of Graphitic Oxide. J. Am. Chem. Soc. 1958, 80, 1339. [Google Scholar] [CrossRef]
- Reiner-Rozman, C.; Kotlowski, C.; Knoll, W. Electronic biosensing with functionalized rGO FETs. Biosensors 2016, 6, 17. [Google Scholar] [CrossRef] [Green Version]
- Predator, F.P. Olfactory Responses of Two Coleopteran Species: The Stored Product Pest Tribolium castaneum and the Forest Pest Predator Dastarcus helophoroides. Doctoral Dissertation, Georg-August-University Göttingen, Göttingen, Germany, 2019. [Google Scholar]
- Bonazza, C.; Zhu, J.; Hasler, R.; Mastrogiacomo, R.; Pelosi, P.; Knoll, W. Responses of the Pheromone-Binding Protein of the Silk Moth Bombyx mori on a Graphene Biosensor Match Binding Constants in Solution. Sensors 2021, 21, 499. [Google Scholar] [CrossRef] [PubMed]
- Dykyj, J.; Svoboda, J.; Wilhoit, R.C.; Frenkel, M.; Hall, K.R. Vapor Pressure and Antoine Constants for Oxygen Containing Organic Compounds; Springer: Berlin, Germany, 2000. [Google Scholar]
- Sander, R. Compilation of Henry’s law constants (version 4.0) for water as solvent. Atmos. Chem. Phys. 2015, 15, 4399–4981. [Google Scholar] [CrossRef] [Green Version]
- Rutter, G.M.; Jung, S.; Klimov, N.N.; Newell, D.B.; Zhitenev, N.B.; Stroscio, J.A. Microscopic polarization in bilayer graphene. Nat. Phys. 2011, 7, 649–655. [Google Scholar] [CrossRef]
- Macchia, E.; Alberga, D.; Manoli, K.; Mangiatordi, G.F.; Magliulo, M.; Palazzo, G.; Giordano, F.; Lattanzi, G.; Torsi, L. Organic bioelectronics probing conformational changes in surface confined proteins. Sci. Rep. 2016, 6, 28085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Kazzy, M.; Hurot, C.; Weerakkody, J.S.; Buhot, A.; Hou, Y. Biomimetic Olfactory Biosensors and Bioelectronic Noses; IFSA Publishing: Barcelona, Spain, 2020; pp. 15–63. [Google Scholar]
- Ricatti, J.; Acquasaliente, L.; Ribaudo, G.; De Filippis, V.; Bellini, M.; Llovera, R.E.; Barollo, S.; Pezzani, R.; Zagotto, G.; Persaud, K.C.; et al. Effects of point mutations in the binding pocket of the mouse major urinary protein MUP20 on ligand affinity and specificity. Sci. Rep. 2019, 9, 300. [Google Scholar] [CrossRef] [Green Version]
- Katada, S.; Hirokawa, T.; Oka, Y.; Suwa, M.; Touhara, K. Structural basis for a broad but selective ligand spectrum of a mouse olfactory receptor: Mapping the odorant-binding site. J. Neurosci. 2005, 25, 1806–1815. [Google Scholar] [CrossRef]




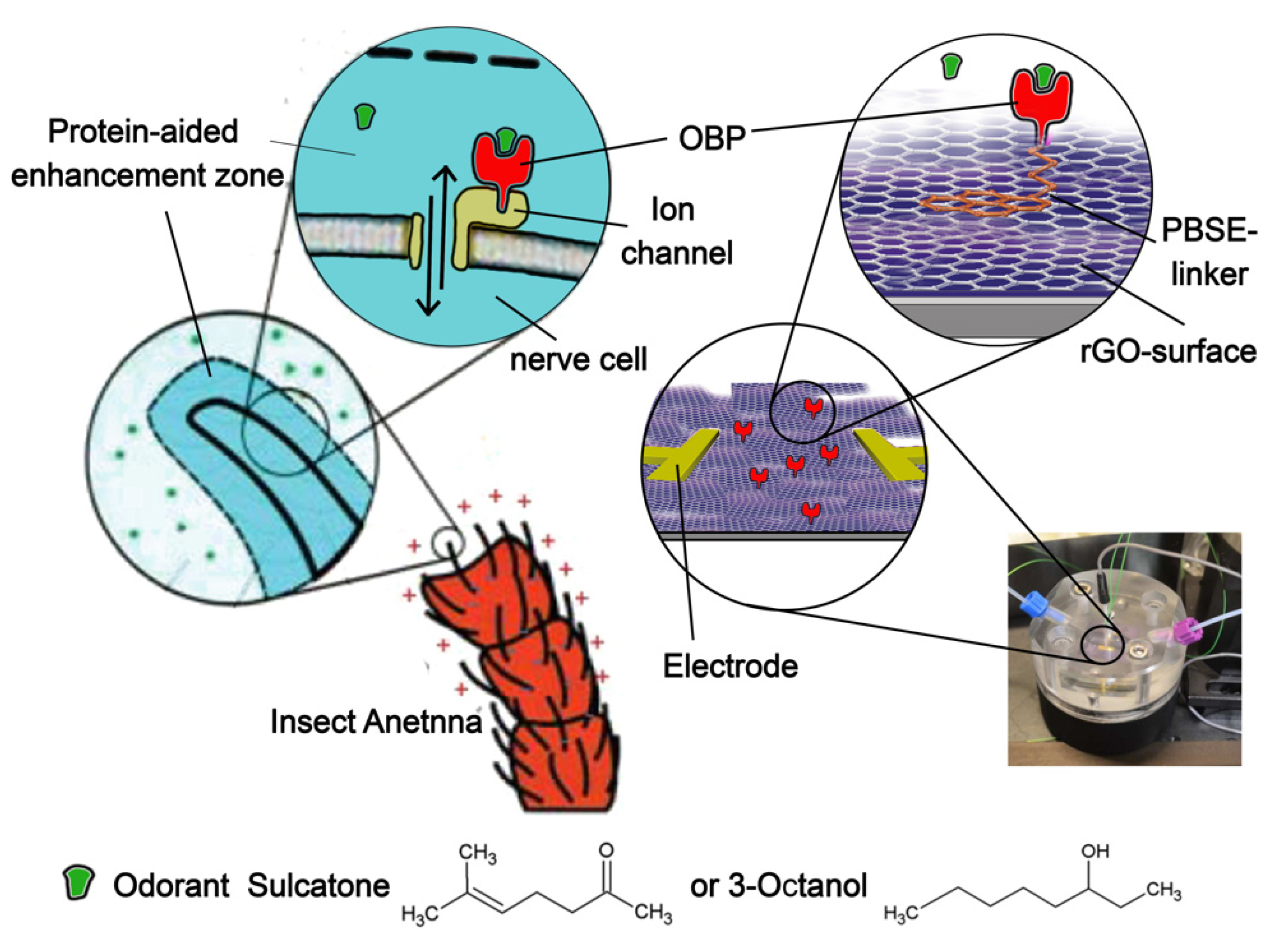{kind=link}
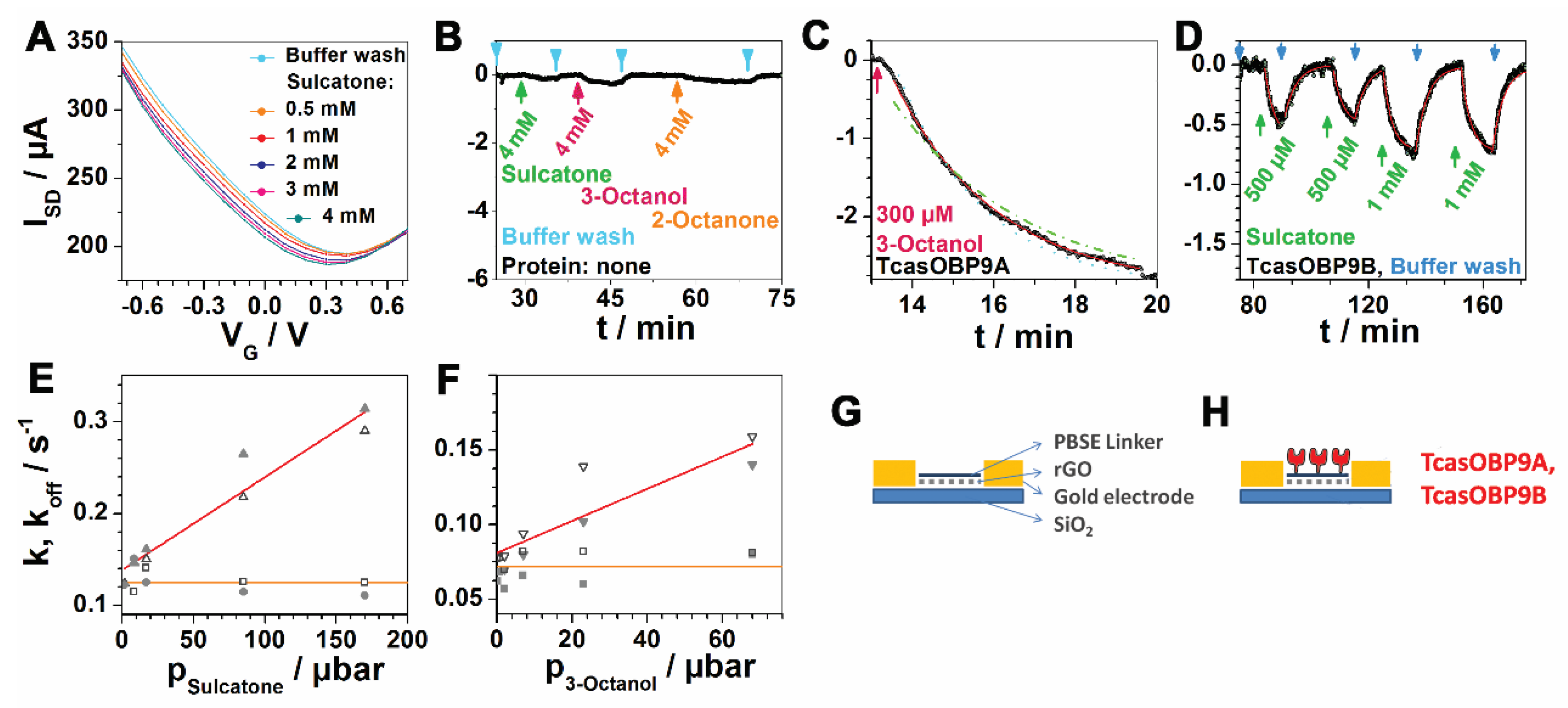{kind=link}
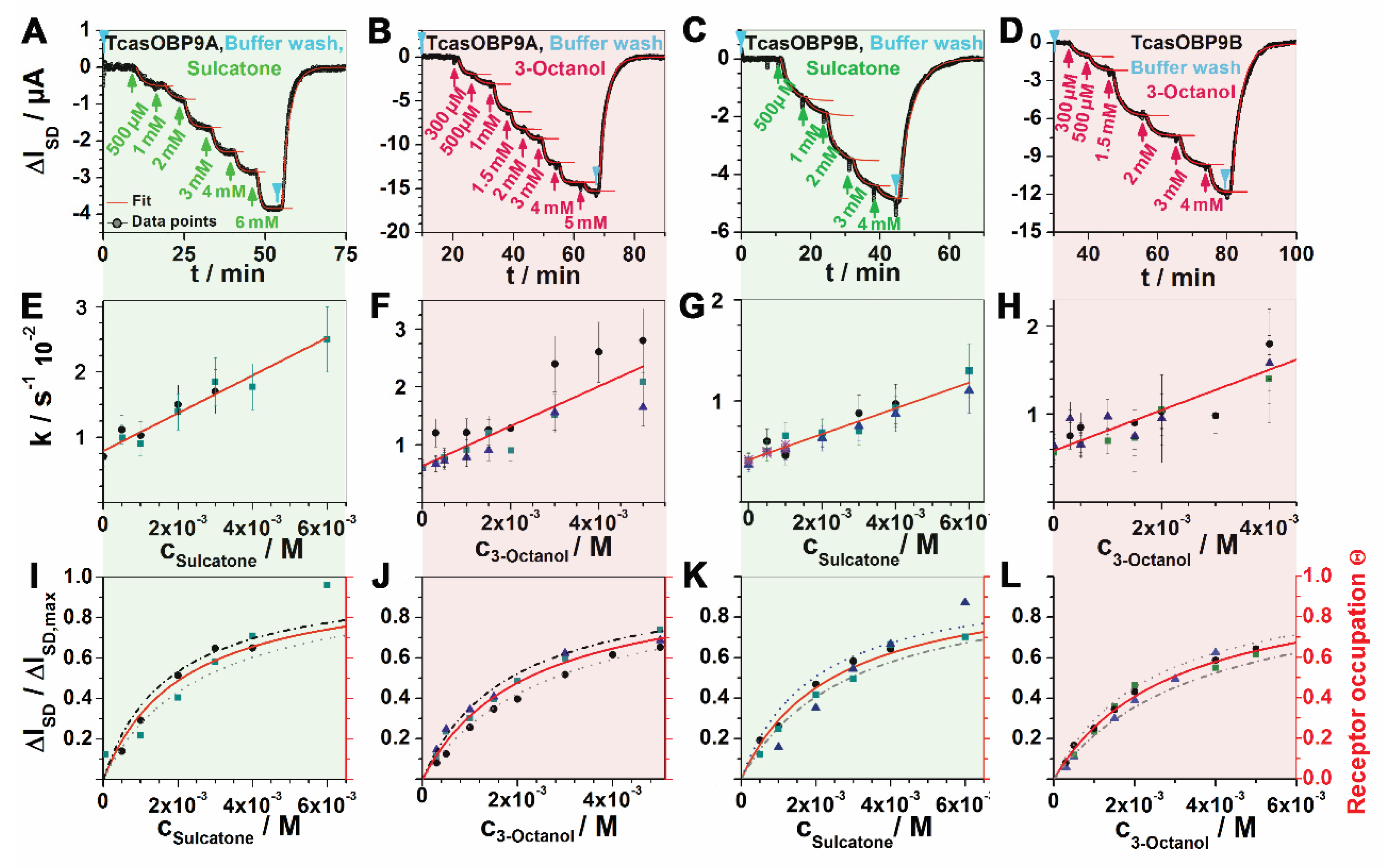{kind=link}
| Protein (Species) | Ligand | KD/μM | kon /M−1 s−1 | koff /s−1 |
|---|---|---|---|---|
| TcasOBP9A | 6-Methyl-5- hepten-2-one (Sulcatone) | 2100 | 2.9 | 0.008 |
| (Tribolium castaneum) | 3-Octanol | 2300 | 3.4 | 0.006 |
| TcasOBP9B | 6-Methyl-5- hepten-2-one (Sulcatone) | 3000 | 1.3 | 0.004 |
| (Tribolium castaneum) | 3-Octanol | 2900 | 2.4 | 0.006 |
| AmelOBP14 [13] | Homovanillic acid | 4 | 1130 | 0.008 |
| (Apis mellifera) | Methyl vanillate | 20 | 230 | 0.010 |
| Eugenol | 40 | 170 | 0.006 | |
| Citral | 800 | 9 | 0.003 | |
| Methyl eugenol | 1400 | 6 | 0.006 | |
| Geraniol | 3300 | 3 | 0.003 | |
| 6-Methyl-5- hepten-2-one (Sulcatone) | 3500 | 5 | 0.008 | |
| Amel SS-OBP14 [13] (Apis mellifera) | Eugenol | 4 | 750 | 0.006 |
| BmorPBP1 [26] | Bombykol (pH 8) | 5 | 20 | 0.000094 |
| (Bombyx mori) | Bombykol (pH 4) | - | no binding | no binding |
| Antenna EAG [25] | 6-Methyl-5- hepten-2-one (Sulcatone) | 1300 | 73 | 0.095 |
| (Tribolium castaenum) | 3-Octanol | 2500 | 32 | 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonazza, C.; Bonazza, K. Kinetics of Odorant Recognition with a Graphene-Based Olfactory Receptor Mimicry. Chemosensors 2022, 10, 203. https://doi.org/10.3390/chemosensors10060203
Bonazza C, Bonazza K. Kinetics of Odorant Recognition with a Graphene-Based Olfactory Receptor Mimicry. Chemosensors. 2022; 10(6):203. https://doi.org/10.3390/chemosensors10060203
Chicago/Turabian StyleBonazza, Caroline, and Klaus Bonazza. 2022. "Kinetics of Odorant Recognition with a Graphene-Based Olfactory Receptor Mimicry" Chemosensors 10, no. 6: 203. https://doi.org/10.3390/chemosensors10060203
APA StyleBonazza, C., & Bonazza, K. (2022). Kinetics of Odorant Recognition with a Graphene-Based Olfactory Receptor Mimicry. Chemosensors, 10(6), 203. https://doi.org/10.3390/chemosensors10060203