
A Taste Bud Organoid-Based Microelectrode Array Biosensor for Taste Sensing

 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Taste Bud Organoids
2.2. Immunostaining of Taste Bud Organoids
2.3. MEA Setup and Measurement
3. Results and Discussion
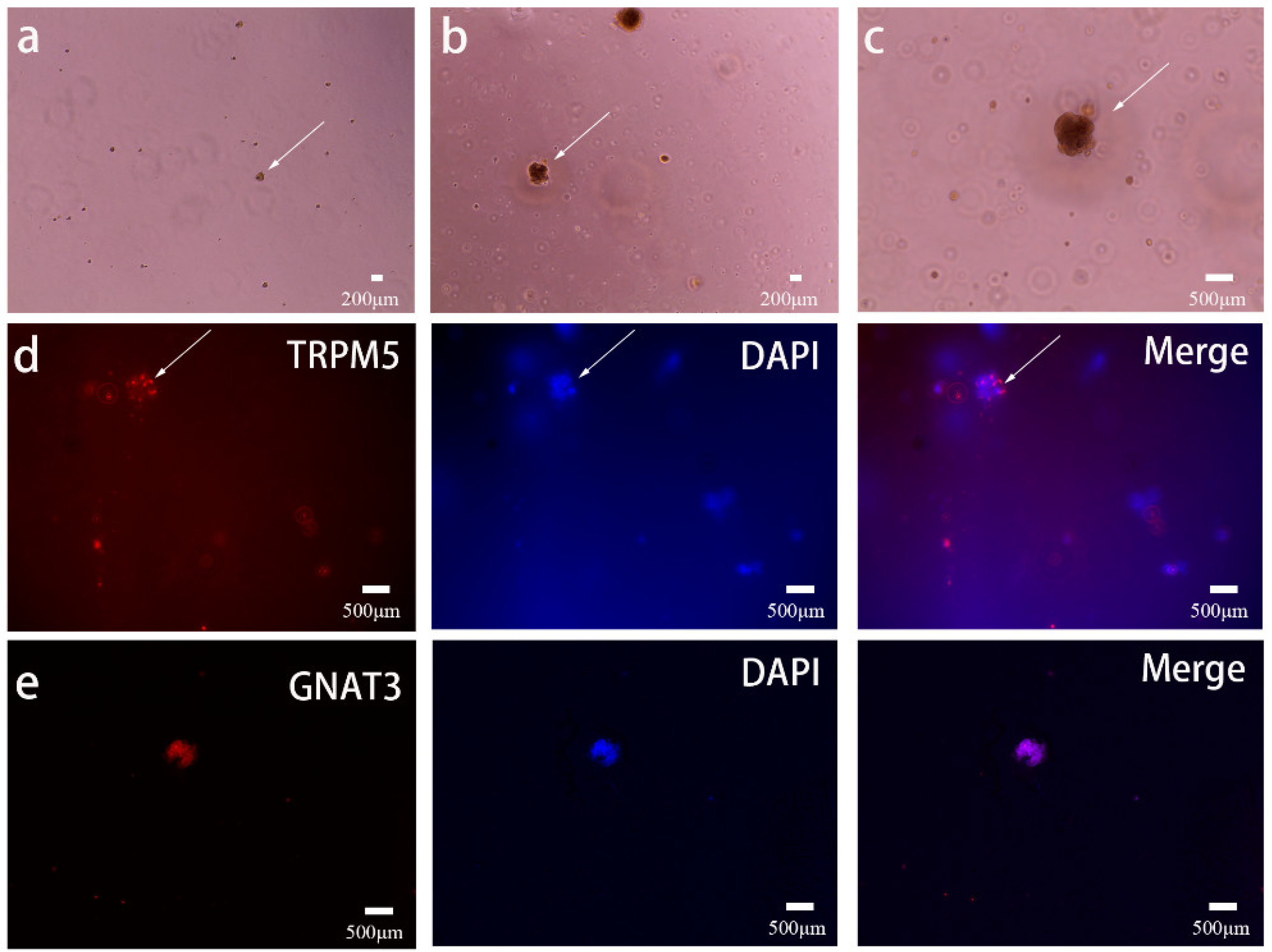
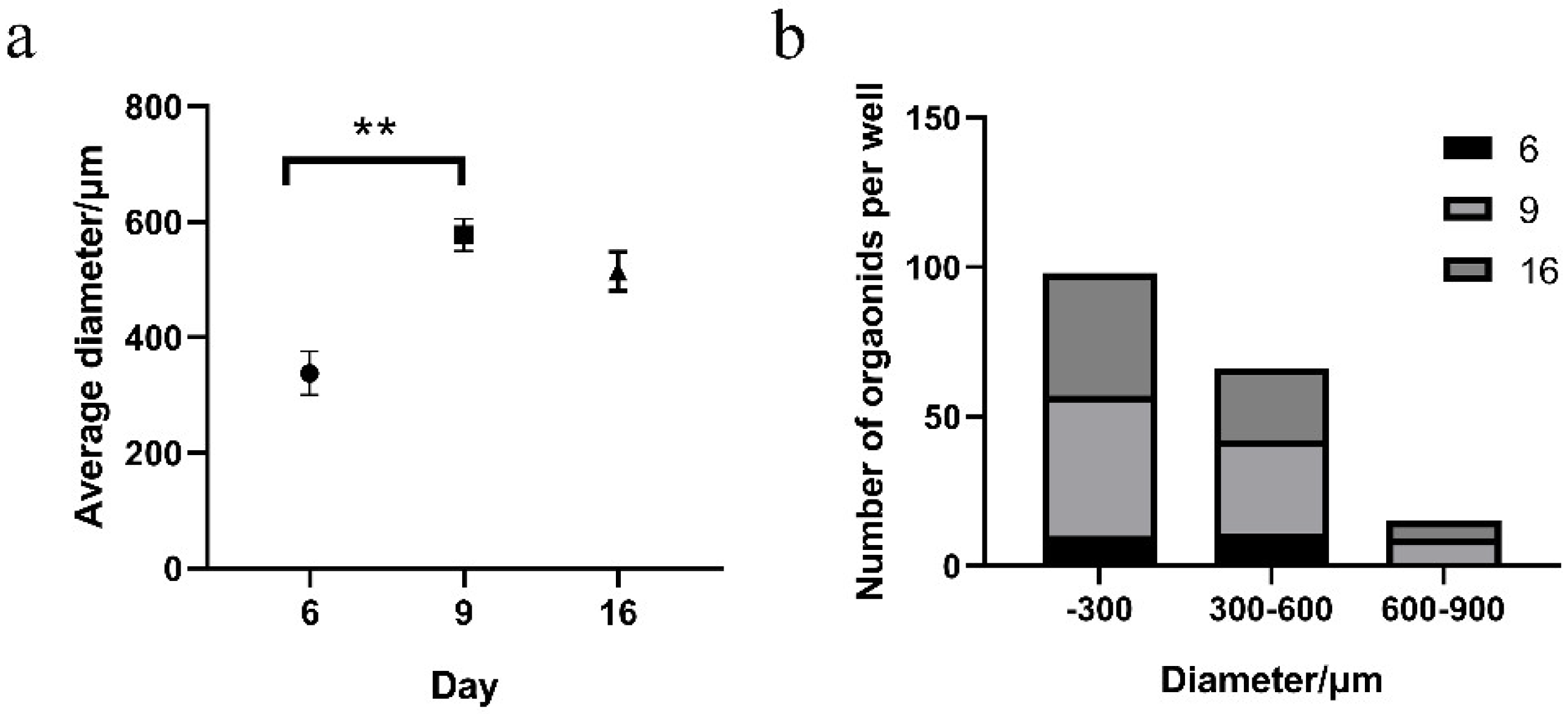
3.1. Preparation of Functional Taste Bud Organoids
3.2. Coupling of Taste Bud Organoids with the MEA Chip
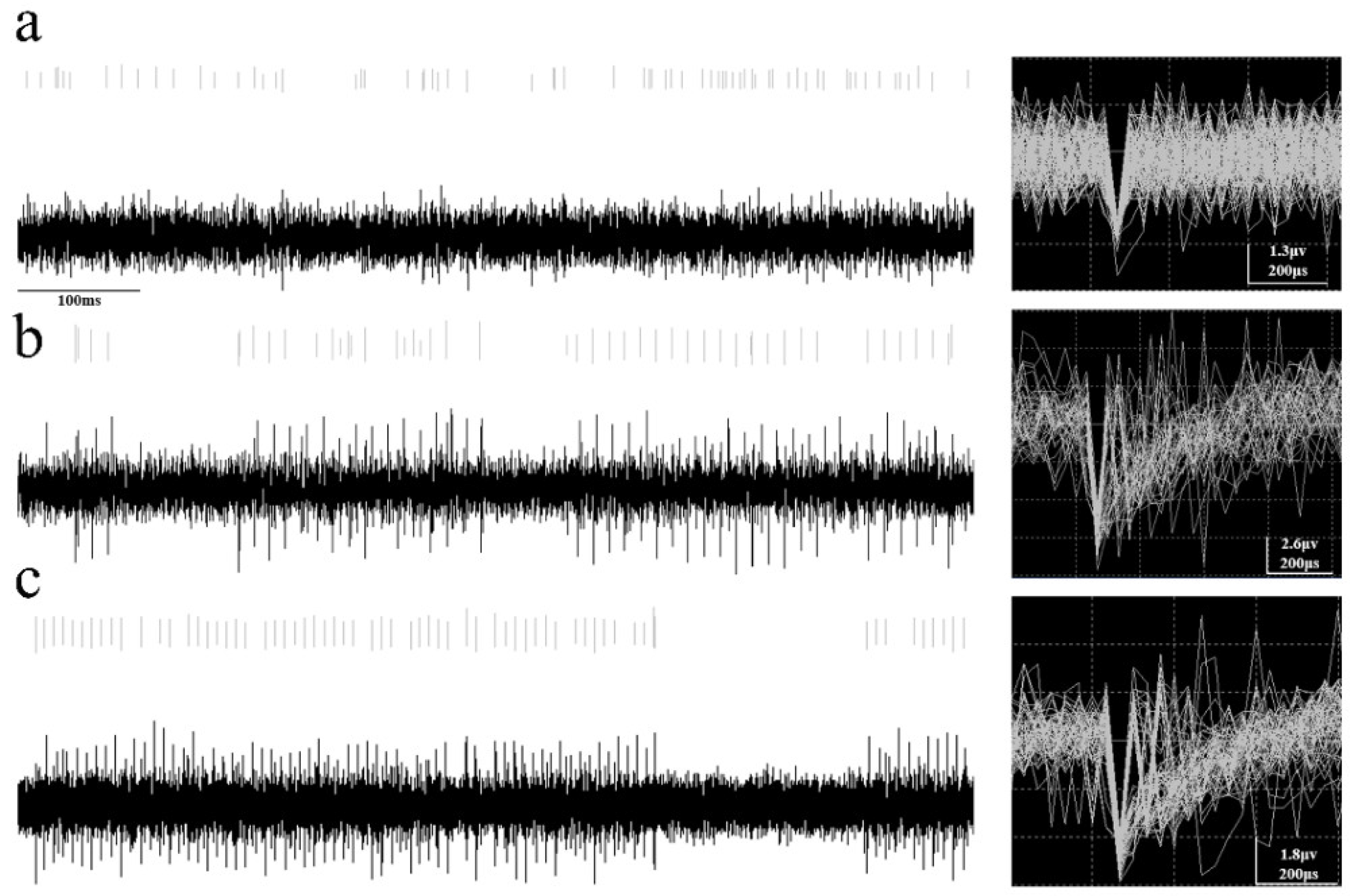
3.3. Performance Testing of Taste Bud Organoid-Based Biosensors
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barlow, L.A. Progress and renewal in gustation: New insights into taste bud development. Development 2015, 142, 3620–3629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardiner, J.; Barton, D.; Vanslambrouck, J.M.; Braet, F.; Hall, D.; Marc, J.; Overall, R. Defects in tongue papillae and taste sensation indicate a problem with neurotrophic support in various neurological diseases. Neurosci. Rev. J. Bringing Neurobiol. Neurol. Psychiatr. 2008, 14, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Wu, Y.; Liao, L.; Tian, W. Oral Organoids: Progress and Challenges. J. Dent. Res. 2021, 100, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Kalmykov, A.; Huang, C.; Bliley, J.; Shiwarski, D.; Tashman, J.; Abdullah, A.; Rastogi, S.K.; Shukla, S.; Mataev, E.; Feinberg, A.W.; et al. Organ-on-e-chip: Three-dimensional self-rolled biosensor array for electrical interrogations of human electrogenic spheroids. Sci. Adv. 2019, 5, eaax0729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.S.; Cho, A.N.; Jin, Y.; Kim, J.; Kim, S.; Cho, S.W. Bio-artificial tongue with tongue extracellular matrix and primary taste cells. Biomaterials 2018, 151, 24–37. [Google Scholar] [CrossRef]
- Qiu, C.; Cui, C.; Hautefort, C.; Haehner, A.; Zhao, J.; Yao, Q.; Zeng, H.; Nisenbaum, E.J.; Liu, L.; Zhao, Y.; et al. Olfactory and Gustatory Dysfunction as an Early Identifier of COVID-19 in Adults and Children: An International Multicenter Study. Otolaryngol. Head Neck Surg. 2020, 163, 714–721. [Google Scholar] [CrossRef]
- Barker, K.E.; Batstone, M.D.; Savage, N.W. Comparison of treatment modalities in burning mouth syndrome. Aust. Dent. J. 2009, 54, 300–305. [Google Scholar] [CrossRef]
- Hampf, G.; Aalberg, V.; Sundén, B. Experiences from a facial pain unit. J. Craniomandib. Disord. Facial Oral Pain 1990, 4, 267–272. [Google Scholar]
- Yun, J.; Cho, A.N.; Cho, S.W.; Nam, Y.S. DNA-mediated self-assembly of taste cells and neurons for taste signal transmission. Biomater. Sci. 2018, 6, 3388–3396. [Google Scholar] [CrossRef]
- Deshpande, D.A.; Wang, W.C.; McIlmoyle, E.L.; Robinett, K.S.; Schillinger, R.M.; An, S.S.; Sham, J.S.; Liggett, S.B. Bitter taste receptors on airway smooth muscle bronchodilate by localized calcium signaling and reverse obstruction. Nat. Med. 2010, 16, 1299–1304. [Google Scholar] [CrossRef] [Green Version]
- Ren, W.; Lewandowski, B.C.; Watson, J.; Aihara, E.; Iwatsuki, K.; Bachmanov, A.A.; Margolskee, R.F.; Jiang, P. Single Lgr5- or Lgr6-expressing taste stem/progenitor cells generate taste bud cells ex vivo. Proc. Natl. Acad. Sci. USA 2014, 111, 16401–16406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Q.; Chen, S.; Rao, X.; Li, Y.; Pan, M.; Fu, G.; Yao, Y.; Gao, X.; Tang, P.; Zhou, Y.; et al. Inhibition of SIRT1 promotes taste bud stem cell survival and mitigates radiation-induced oral mucositis in mice. Am. J. Transl. Res. 2019, 11, 4789–4799. [Google Scholar] [PubMed]
- Drost, J.; Clevers, H. Organoids in cancer research. Nat. Rev. Cancer 2018, 18, 407–418. [Google Scholar] [CrossRef]
- Lacroix, J.J.; Campos, F.V.; Frezza, L.; Bezanilla, F. Molecular bases for the asynchronous activation of sodium and potassium channels required for nerve impulse generation. Neuron 2013, 79, 651–657. [Google Scholar] [CrossRef] [Green Version]
- Cummings, T.A.; Daniels, C.; Kinnamon, S.C. Sweet taste transduction in hamster: Sweeteners and cyclic nucleotides depolarize taste cells by reducing a K+ current. J. Neurophysiol. 1996, 75, 1256–1263. [Google Scholar] [CrossRef] [PubMed]
- Avenet, P.; Lindemann, B. Noninvasive recording of receptor cell action potentials and sustained currents from single taste buds maintained in the tongue: The response to mucosal NaCl and amiloride. J. Membr. Biol. 1991, 124, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Fujiyama, R.; Okada, Y.; Sato, T. Sour transduction involves activation of NPPB-sensitive conductance in mouse taste cells. J. Neurophysiol. 1998, 80, 1852–1859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furue, H.; Yoshii, K. In situ tight-seal recordings of taste substance-elicited action currents and voltage-gated Ba currents from single taste bud cells in the peeled epithelium of mouse tongue. Brain Res. 1997, 776, 133–139. [Google Scholar] [CrossRef]
- Roper, S.D.; Chaudhari, N. Taste buds: Cells, signals and synapses. Nat. Rev. Neurosci. 2017, 18, 485–497. [Google Scholar] [CrossRef]
- Rössler, P.; Kroner, C.; Freitag, J.; Noè, J.; Breer, H. Identification of a phospholipase C beta subtype in rat taste cells. Eur. J. Cell Biol. 1998, 77, 253–261. [Google Scholar] [CrossRef]
- Miyoshi, M.A.; Abe, K.; Emori, Y. IP(3) receptor type 3 and PLCbeta2 are co-expressed with taste receptors T1R and T2R in rat taste bud cells. Chem. Senses 2001, 26, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhao, Z.; Margolskee, R.; Liman, E. The transduction channel TRPM5 is gated by intracellular calcium in taste cells. J. Neurosci. 2007, 27, 5777–5786. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Wang, J.; Chen, W.; Zhao, L.; Wu, C.; Wang, P. Dual functional extracellular recording using a light-addressable potentiometric sensor for bitter signal transduction. Anal. Chim. Acta 2018, 1022, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhang, F.; Zhang, D.; Hu, N.; Hsia, K.J.; Wang, P. Extracellular potentials recording in intact taste epithelium by microelectrode array for a taste sensor. Biosens. Bioelectron. 2013, 43, 186–192. [Google Scholar] [CrossRef]
- Ahn, S.R.; An, J.H.; Song, H.S.; Park, J.W.; Lee, S.H.; Kim, J.H.; Jang, J.; Park, T.H. Duplex Bioelectronic Tongue for Sensing Umami and Sweet Tastes Based on Human Taste Receptor Nanovesicles. ACS Nano 2016, 10, 7287–7296. [Google Scholar] [CrossRef]
- Wang, J.; Kong, S.; Chen, F.; Chen, W.; Du, L.; Cai, W.; Huang, L.; Wu, C.; Zhang, D.W. A bioelectronic taste sensor based on bioengineered Escherichia coli cells combined with ITO-constructed electrochemical sensors. Anal. Chim. Acta 2019, 1079, 73–78. [Google Scholar] [CrossRef]
- Son, M.; Kim, D.; Ko, H.J.; Hong, S.; Park, T.H. A portable and multiplexed bioelectronic sensor using human olfactory and taste receptors. Biosens. Bioelectron. 2017, 87, 901–907. [Google Scholar] [CrossRef]
- Wu, C.; Du, L.; Zou, L.; Huang, L.; Wang, P. A biomimetic bitter receptor-based biosensor with high efficiency immobilization and purification using self-assembled aptamers. Analyst 2013, 138, 5989–5994. [Google Scholar] [CrossRef]
- Chen, J.; Wise, K.D.; Hetke, J.F.; Bledsoe, S.C., Jr. A multichannel neural probe for selective chemical delivery at the cellular level. IEEE Trans. Bio-Med. Eng. 1997, 44, 760–769. [Google Scholar] [CrossRef]
- Takeuchi, S.; Ziegler, D.; Yoshida, Y.; Mabuchi, K.; Suzuki, T. Parylene flexible neural probes integrated with microfluidic channels. Lab Chip 2005, 5, 519–523. [Google Scholar] [CrossRef]
- McCall, J.G.; Qazi, R.; Shin, G.; Li, S.; Ikram, M.H.; Jang, K.I.; Liu, Y.; Al-Hasani, R.; Bruchas, M.R.; Jeong, J.W.; et al. Preparation and implementation of optofluidic neural probes for in vivo wireless pharmacology and optogenetics. Nat. Protoc. 2017, 12, 219–237. [Google Scholar] [CrossRef] [PubMed]
- Blanche, T.J.; Spacek, M.A.; Hetke, J.F.; Swindale, N.V. Polytrodes: High-density silicon electrode arrays for large-scale multiunit recording. J. Neurophysiol. 2005, 93, 2987–3000. [Google Scholar] [CrossRef] [PubMed]
- Obien, M.E.; Deligkaris, K.; Bullmann, T.; Bakkum, D.J.; Frey, U. Revealing neuronal function through microelectrode array recordings. Front. Neurosci. 2014, 8, 423. [Google Scholar] [CrossRef] [Green Version]
- Lebreton, F.; Pirog, A.; Belouah, I.; Bosco, D.; Berney, T.; Meda, P.; Bornat, Y.; Catargi, B.; Renaud, S.; Raoux, M.; et al. Slow potentials encode intercellular coupling and insulin demand in pancreatic beta cells. Diabetologia 2015, 58, 1291–1299. [Google Scholar] [CrossRef] [Green Version]
- Hennig, M.H.; Hurwitz, C.; Sorbaro, M. Scaling Spike Detection and Sorting for Next-Generation Electrophysiology. Adv. Neurobiol. 2019, 22, 171–184. [Google Scholar] [CrossRef] [Green Version]
- Takekawa, T.; Isomura, Y.; Fukai, T. Spike sorting of heterogeneous neuron types by multimodality-weighted PCA and explicit robust variational Bayes. Front. Neuroinform. 2012, 6, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quiroga, R.Q.; Nadasdy, Z.; Ben-Shaul, Y. Unsupervised spike detection and sorting with wavelets and superparamagnetic clustering. Neural Comput. 2004, 16, 1661–1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, C.A., Jr.; Springer, P.A.; Loeb, G.E.; Berwald-Netter, Y.; Okun, L.M. A miniature microelectrode array to monitor the bioelectric activity of cultured cells. Exp. Cell Res. 1972, 74, 61–66. [Google Scholar] [CrossRef]
- Pine, J. Recording action potentials from cultured neurons with extracellular microcircuit electrodes. J. Neurosci. Methods 1980, 2, 19–31. [Google Scholar] [CrossRef]
- Delgado Ruz, I.; Schultz, S.R. Localising and classifying neurons from high density MEA recordings. J. Neurosci. Methods 2014, 233, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Ren, W.; Liu, Q.; Zhang, X.; Yu, Y. Age-related taste cell generation in circumvallate papillae organoids via regulation of multiple signaling pathways. Exp. Cell Res. 2020, 394, 112150. [Google Scholar] [CrossRef] [PubMed]
- Kinnamon, S.C. Taste receptor signalling from tongues to lungs. Acta Physiol. 2012, 204, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Le Gléau, L.; Rouault, C.; Osinski, C.; Prifti, E.; Soula, H.A.; Debédat, J.; Busieau, P.; Amouyal, C.; Clément, K.; Andreelli, F.; et al. Intestinal alteration of α-gustducin and sweet taste signaling pathway in metabolic diseases is partly rescued after weight loss and diabetes remission. Am. J. Physiology. Endocrinol. Metab. 2021, 321, E417–E432. [Google Scholar] [CrossRef] [PubMed]
- Parsons, J.T.; Horwitz, A.R.; Schwartz, M.A. Cell adhesion: Integrating cytoskeletal dynamics and cellular tension. Nat. Rev. Mol. Cell Biol. 2010, 11, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Kanchanawong, P.; Shtengel, G.; Pasapera, A.M.; Ramko, E.B.; Davidson, M.W.; Hess, H.F.; Waterman, C.M. Nanoscale architecture of integrin-based cell adhesions. Nature 2010, 468, 580–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paszek, M.J.; DuFort, C.C.; Rossier, O.; Bainer, R.; Mouw, J.K.; Godula, K.; Hudak, J.E.; Lakins, J.N.; Wijekoon, A.C.; Cassereau, L.; et al. The cancer glycocalyx mechanically primes integrin-mediated growth and survival. Nature 2014, 511, 319–325. [Google Scholar] [CrossRef] [Green Version]
- Lewis, C.M.; Bosman, C.A.; Fries, P. Recording of brain activity across spatial scales. Curr. Opin. Neurobiol. 2015, 32, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Vatanparast, J.; Khalili, S.; Naseh, M. Dual effects of eugenol on the neuronal excitability: An in vitro study. Neurotoxicology 2017, 58, 84–91. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Zhu, P.; Tian, Y.; Chen, Y.; Liu, Y.; Wang, M.; Chen, W.; Du, L.; Wu, C. A Taste Bud Organoid-Based Microelectrode Array Biosensor for Taste Sensing. Chemosensors 2022, 10, 208. https://doi.org/10.3390/chemosensors10060208
Liu S, Zhu P, Tian Y, Chen Y, Liu Y, Wang M, Chen W, Du L, Wu C. A Taste Bud Organoid-Based Microelectrode Array Biosensor for Taste Sensing. Chemosensors. 2022; 10(6):208. https://doi.org/10.3390/chemosensors10060208
Chicago/Turabian StyleLiu, Shuge, Ping Zhu, Yulan Tian, Yating Chen, Yage Liu, Miaomiao Wang, Wei Chen, Liping Du, and Chunsheng Wu. 2022. "A Taste Bud Organoid-Based Microelectrode Array Biosensor for Taste Sensing" Chemosensors 10, no. 6: 208. https://doi.org/10.3390/chemosensors10060208
APA StyleLiu, S., Zhu, P., Tian, Y., Chen, Y., Liu, Y., Wang, M., Chen, W., Du, L., & Wu, C. (2022). A Taste Bud Organoid-Based Microelectrode Array Biosensor for Taste Sensing. Chemosensors, 10(6), 208. https://doi.org/10.3390/chemosensors10060208