
Design of DNA-Based Artificial Transmembrane Channels for Biosensing and Biomedical Applications


Abstract
:1. Introduction
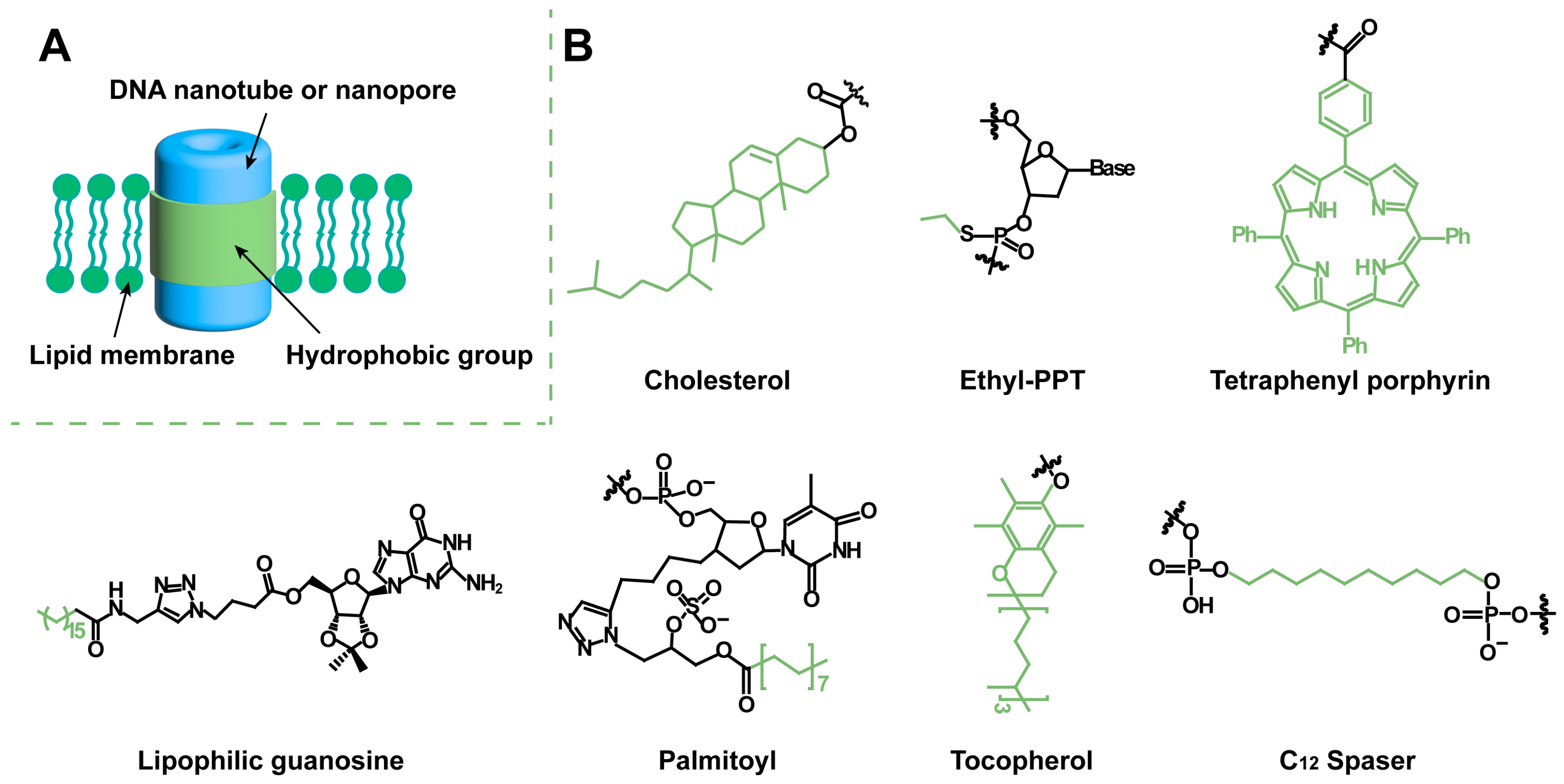
2. Hydrophobic Modification for Artificial Transmembrane Channels
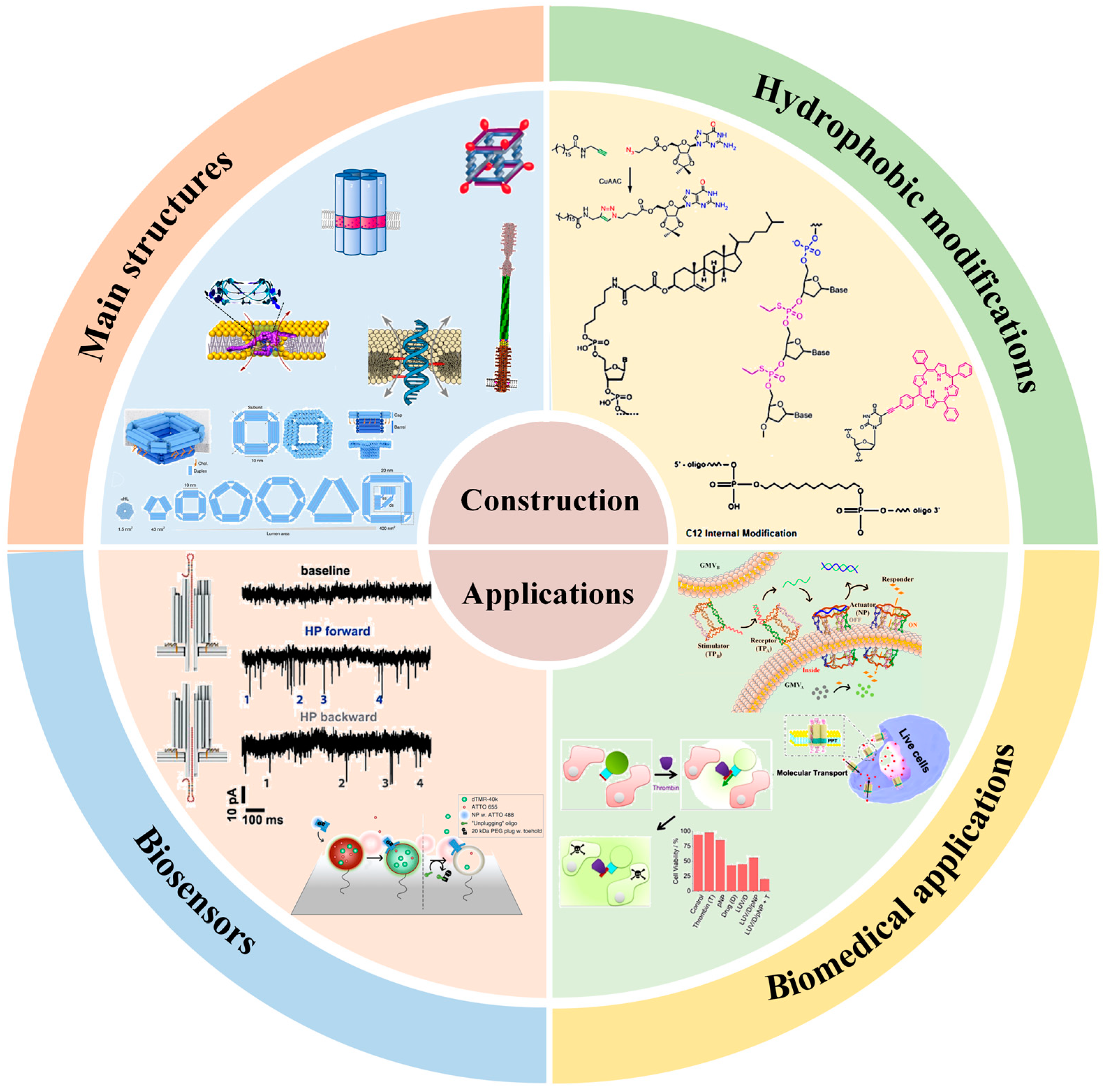
3. Design of DNA Nanostructure for Artificial Transmembrane Channels
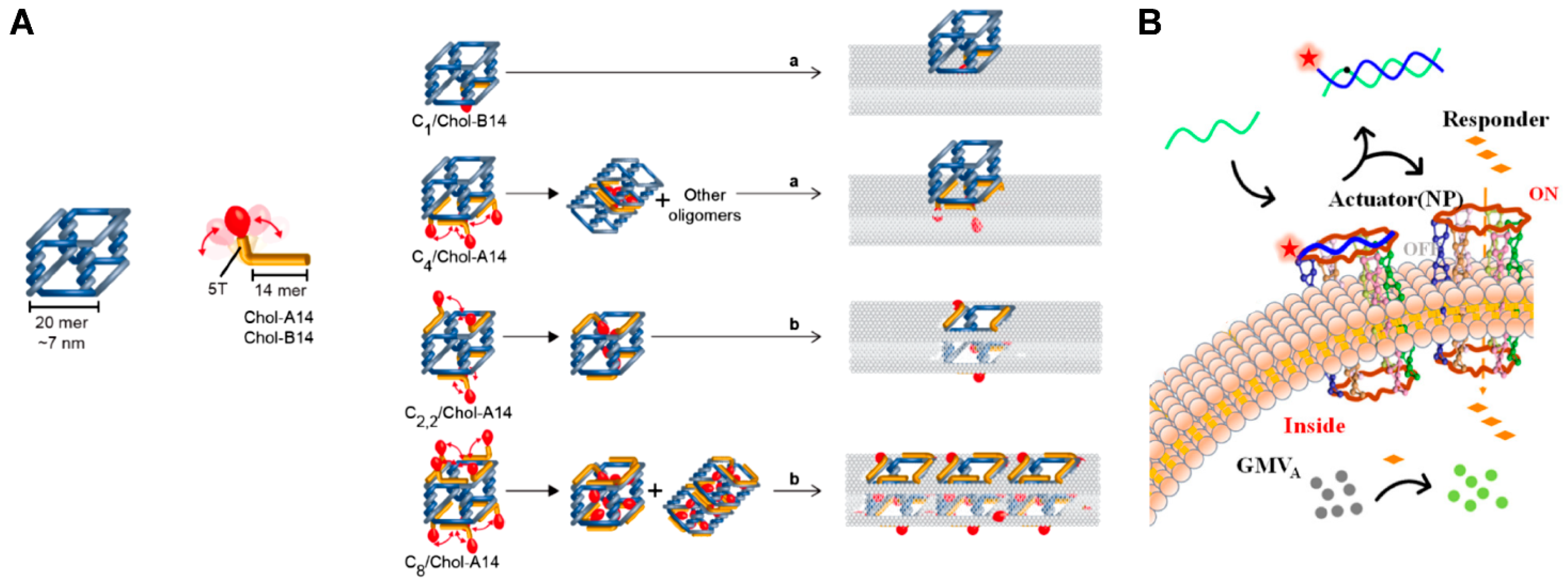
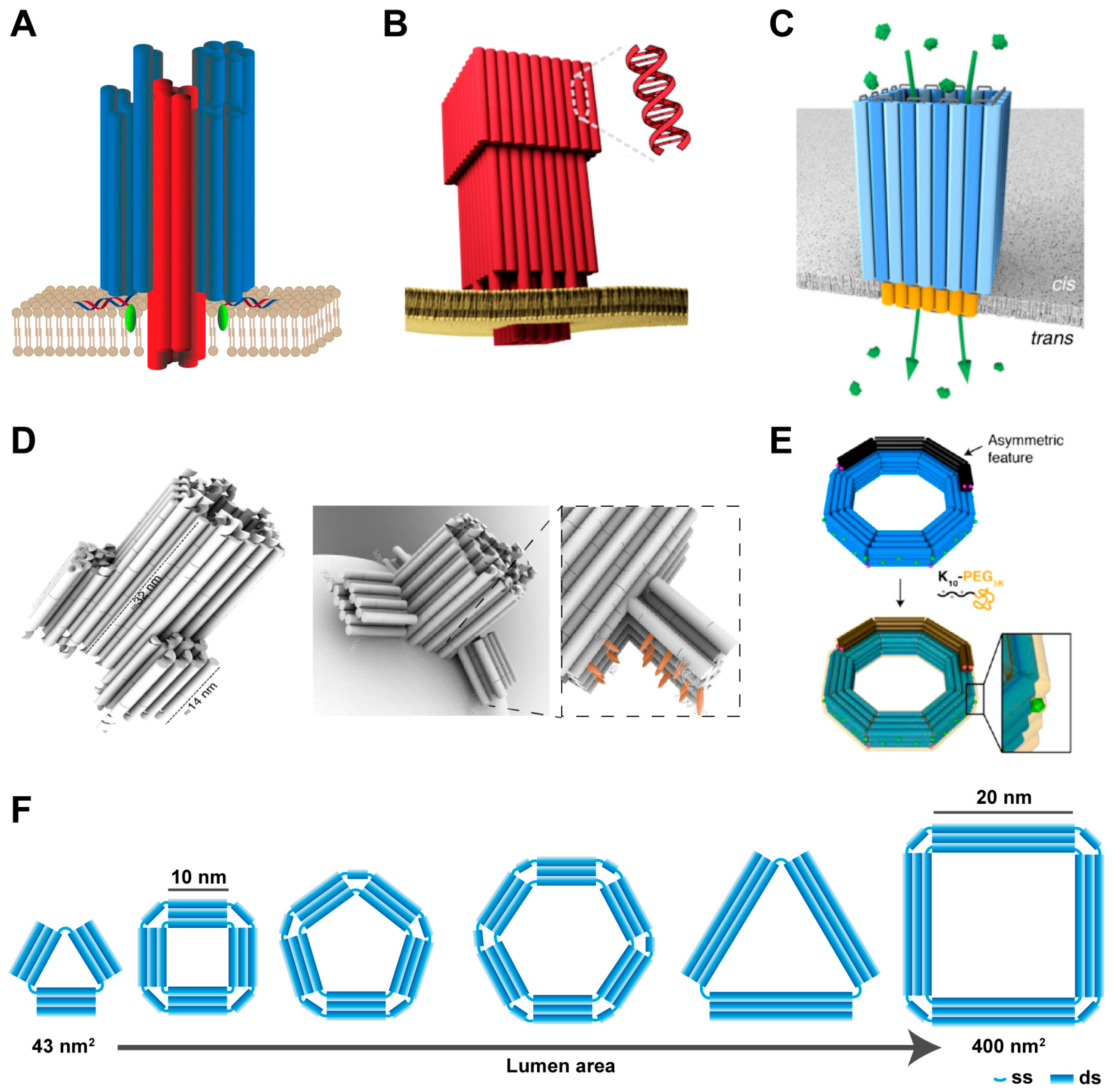
3.1. DNA Wireframe-Based Transmembrane Channels
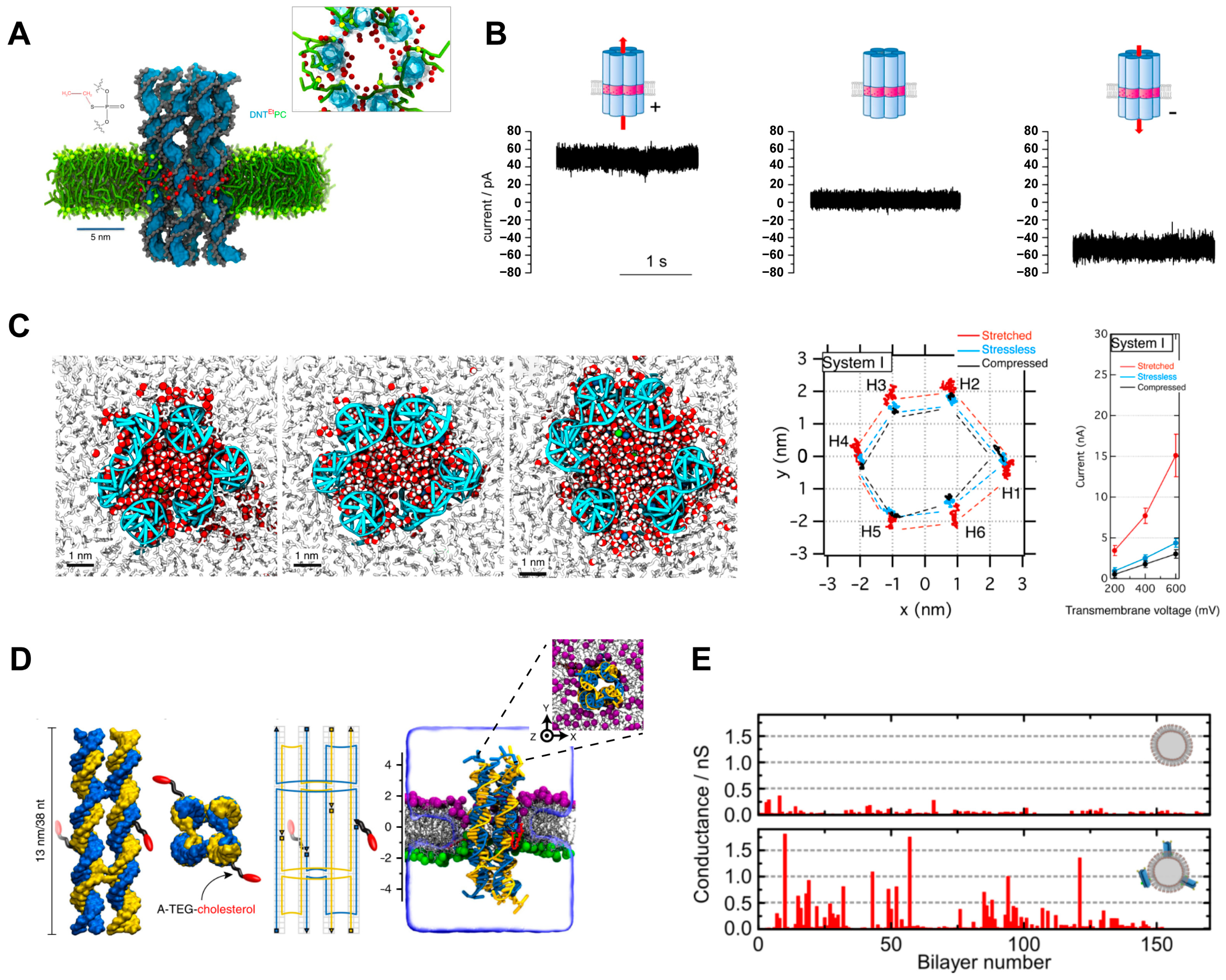
3.2. DNA Helix Bundle-Based Transmembrane Channels
3.3. DNA Tile-Based Transmembrane Channels
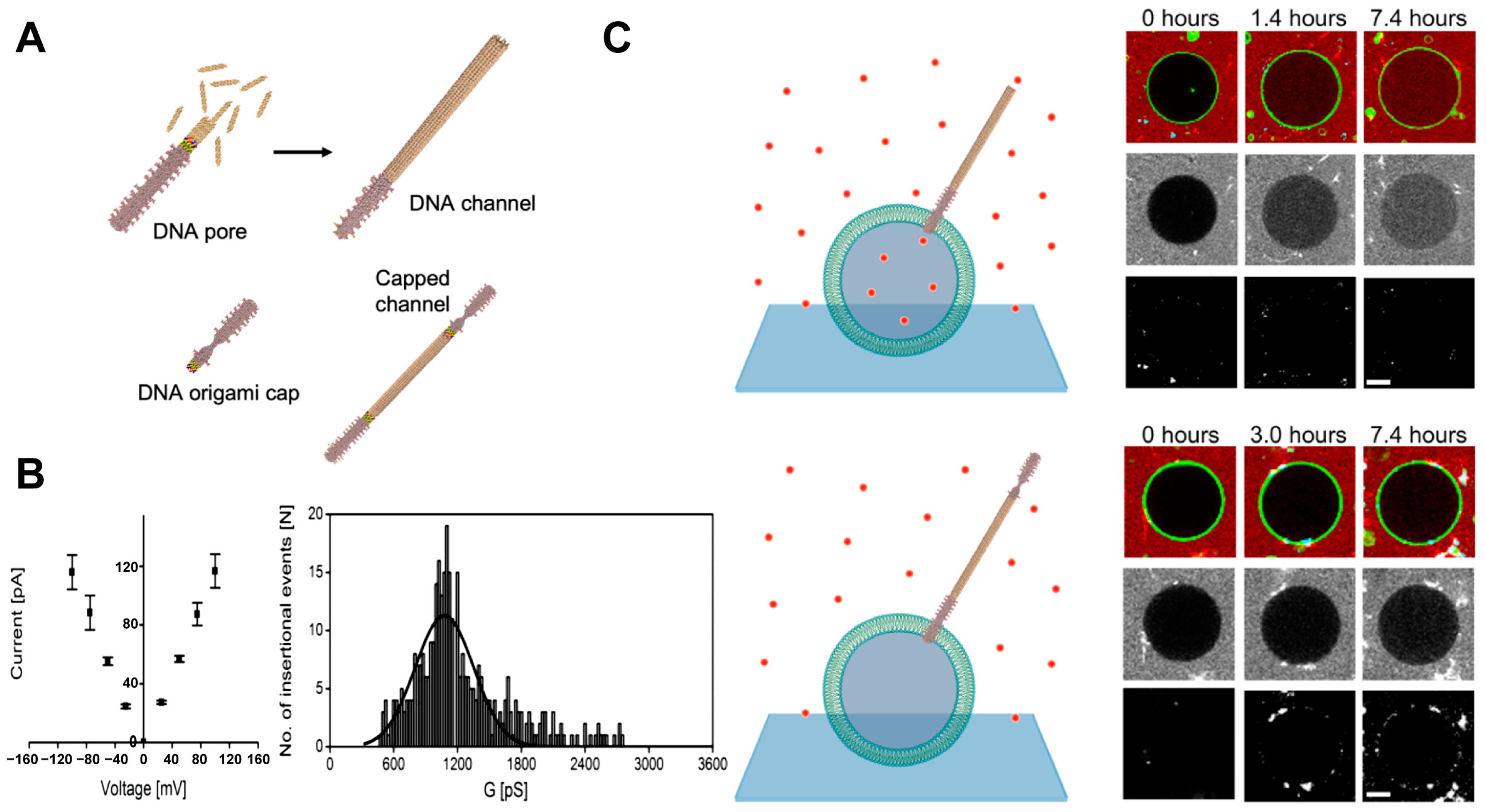
3.4. DNA Origami-Based Transmembrane Channels
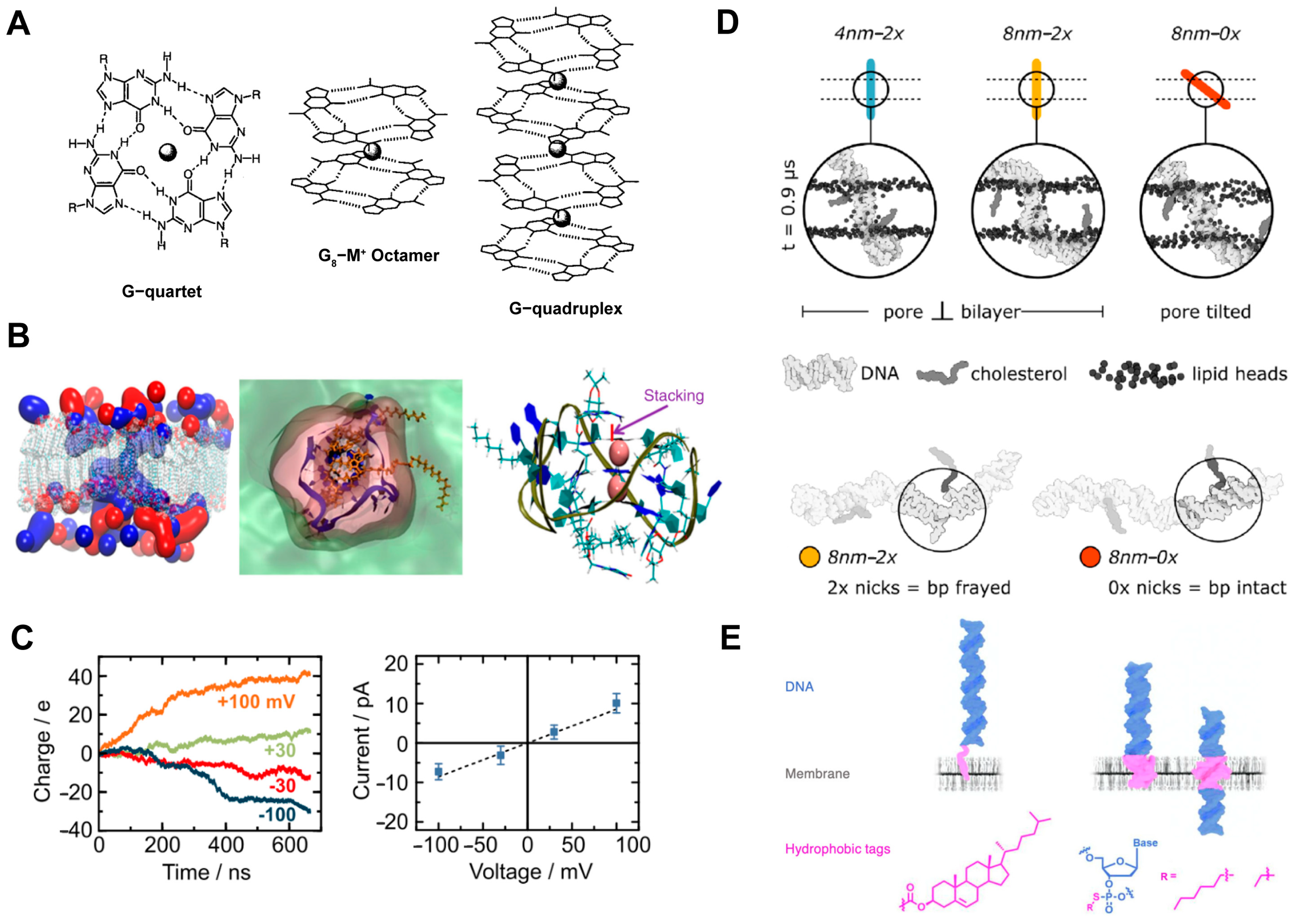
3.5. Other DNA-Based Transmembrane Channels
4. Artificial Transmembrane Channels for Biosensing and Biomedical Applications
4.1. DNA-Based Transmembrane Channels for Biosensors
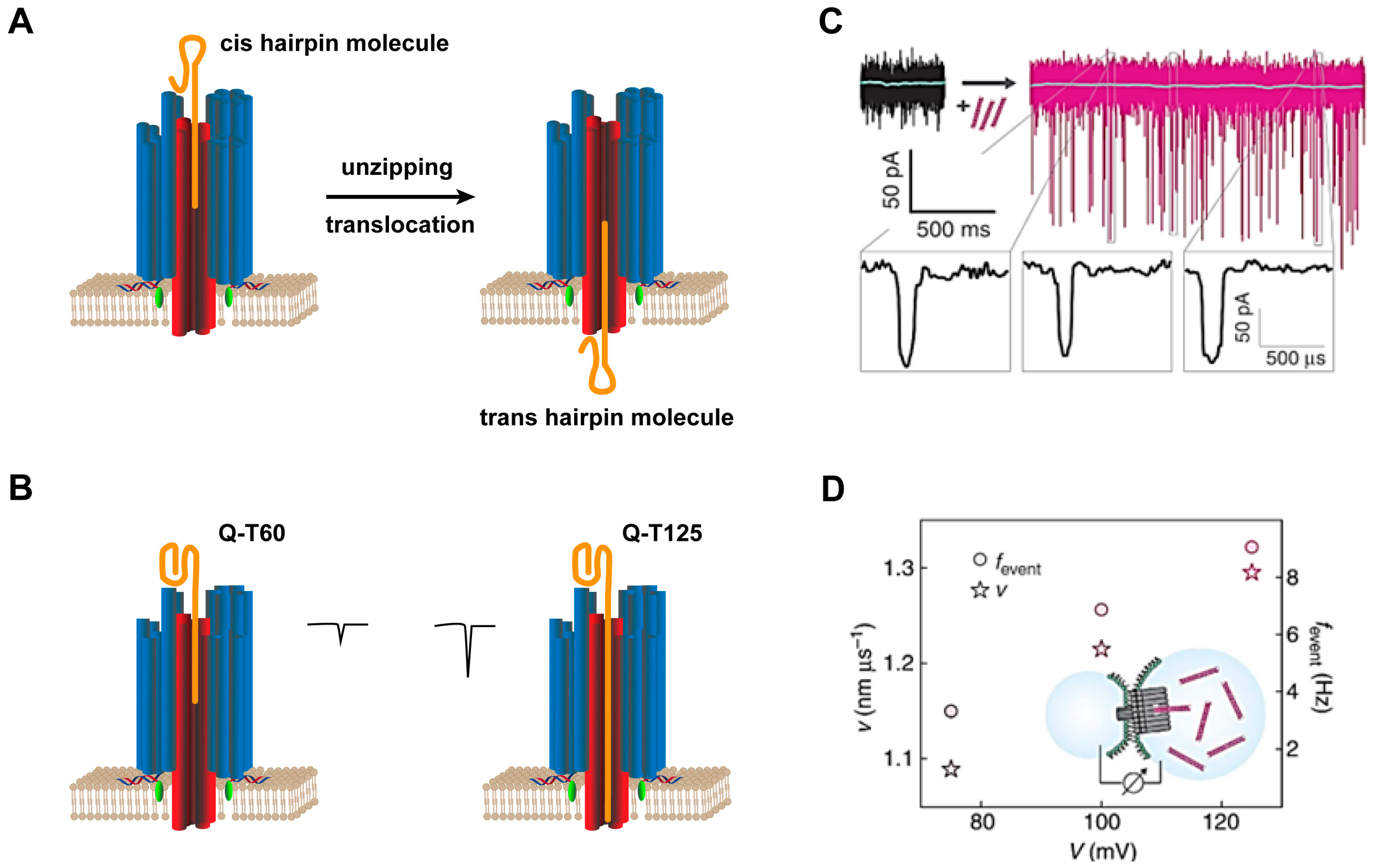
4.1.1. Single-Molecule Nanochannel Sensors
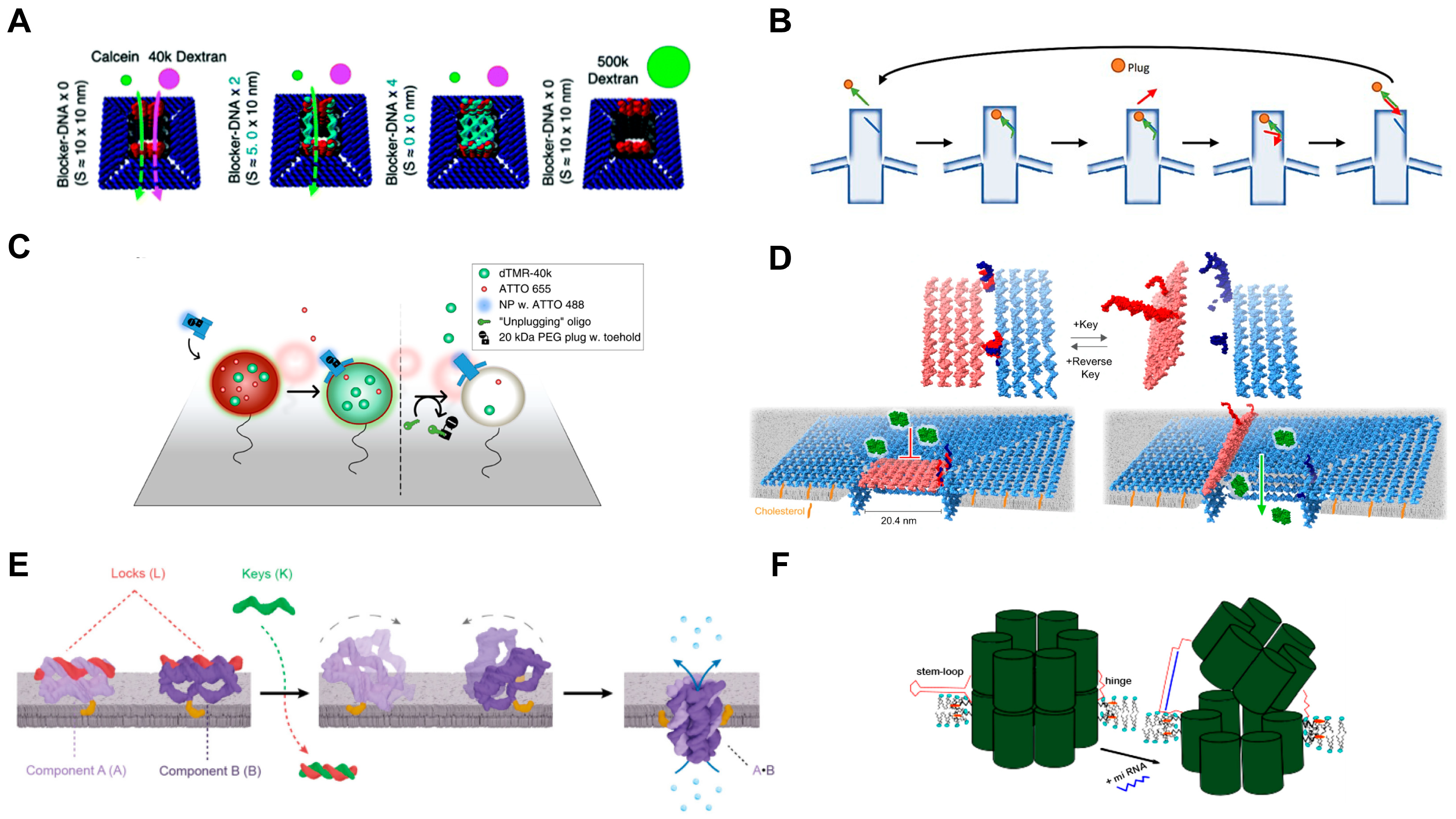
4.1.2. Ligand-Gated Artificial Transmembrane Channels
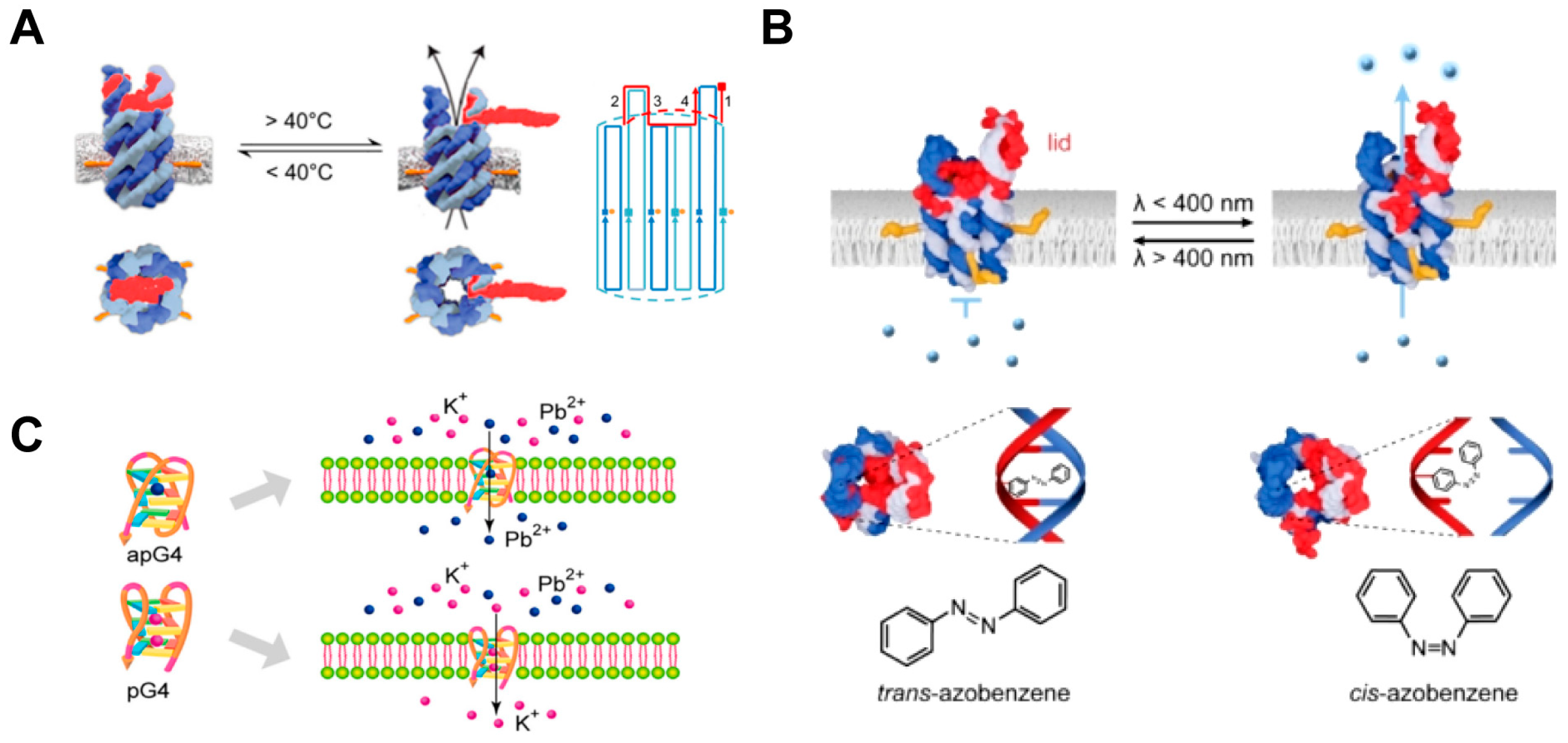
4.1.3. Environmental Stimuli-Responsive Artificial Transmembrane Channels
4.2. DNA-Based Transmembrane Channels for Biomedical Applications
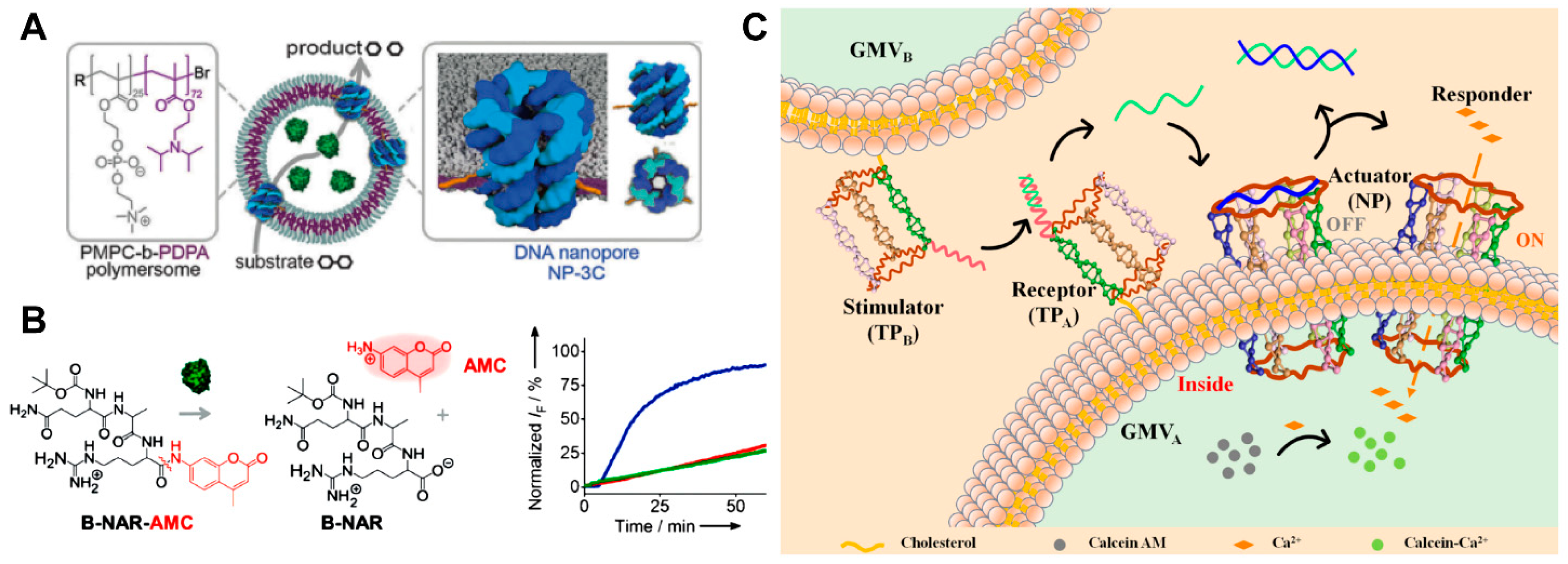
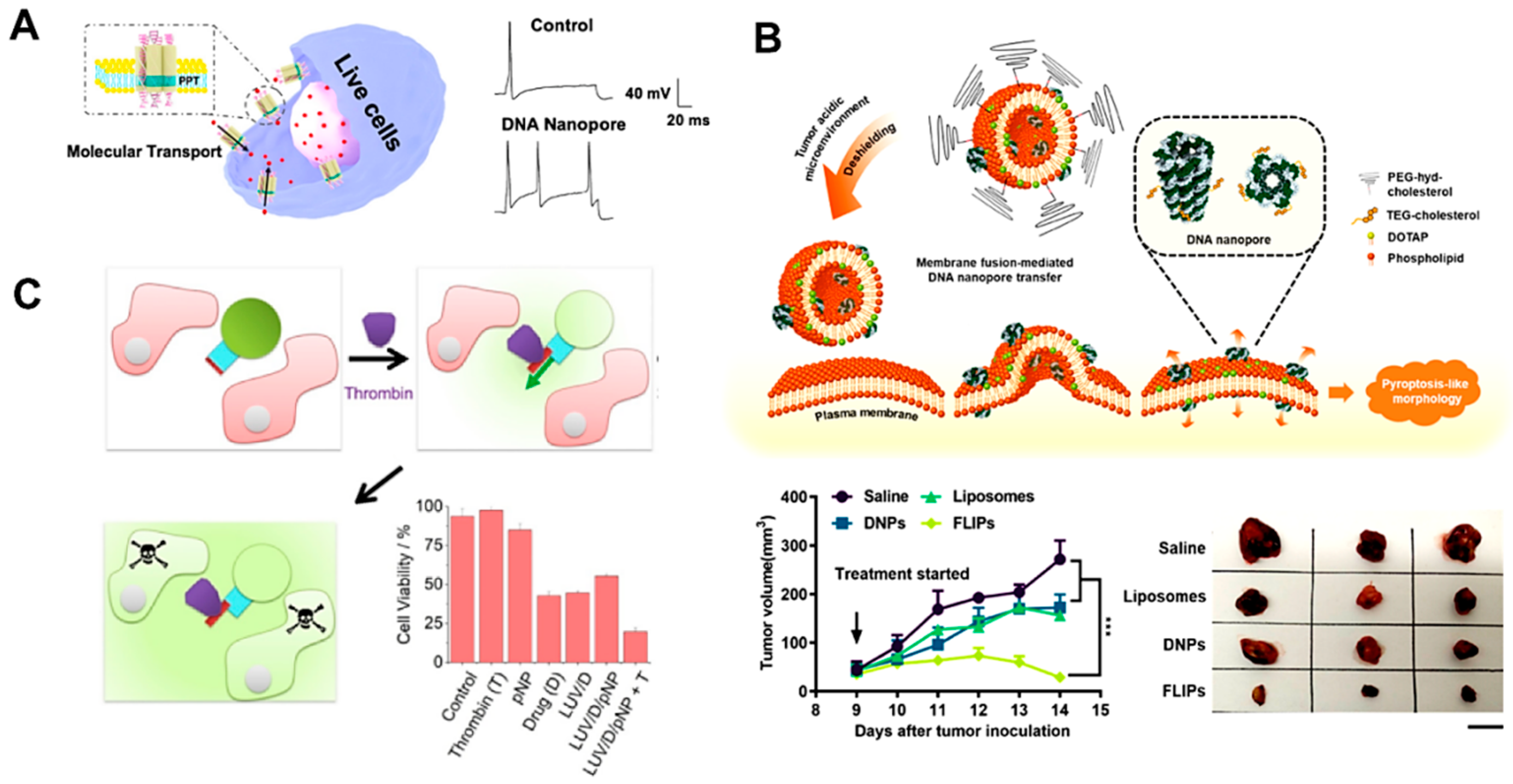
4.2.1. Cell Mimics for Transmembrane Transport
4.2.2. Transmembrane Channels for Cell Death

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Pore Size and Length (nm) | Hydrophobic Modification | Applications | Refs. |
|---|---|---|---|---|
| Cubical | 7; 7 | 4 or 8 cholesterols on the two opposite faces of the cube | Dye molecular transmembrane transport | [23] |
| Hexagonal prism | - | 4 cholesterols | Transport Ca2+ across the membrane as a part of the artificial signal transduction system | [42] |
| 6-helix bundle | 2; - | Ethyl-phosphorothioate | 1. K+ transmembrane transport 2. Selectively cytotoxic to cervical cancer cells 3. Transport ions and antitumor drugs (DOX) to neurons and cancer cells, respectively | [31,44,45] |
| 4-helix bundle | 0.8; - | 2 cholesterols | Ion conduction | [21,49] |
| Micron-length | 7.3; Micron | 12 cholesterols | Leakless end-to-end transport of carboxytetramethyl-rhodamine | [57] |
| Capped | 2; 47 | 26 cholesterols at the underside of the subunits’ caps | DNA hairpin and guanine quadruplex translocation | [25] |
| Funnel-shaped | 6; 54 | 19 cholesterols | Large ionic conductance | [26] |
| Protein-conductive | 7.5; 46 | 24 cholesterols | Proteins transmembrane transport | [27] |
| Large size-selective | 9; 32 | 18 cholesterols and 28 palmitoyls at the terminal of channel and the underside of the flap | 1. Size-specific cargo translocation 2. Real-time direct observation of sensing of an oligonucleotide unplugging strand | [28] |
| Ultrawide | 35; 10 | 32 cholesterols | Transmembrane transport of green fluorescent protein (GFP) | [29] |
| Highly shape- and size-tunable | 8.66~20; - | Underside of the subunits’ caps (10 cholesterols on each 10-nm subunit, 15 cholesterols on each 20-nm subunit) | 1. IgG sensing 2. Human SARS-CoV-2 anti-bodies detection | [30] |
| G-quadruplex and liponucleoside | - | Lipophilic guanosine binds to the G-quadruplex with a 3:1 stoichiometry | The transport of K+ across CHO and K-562 cell membranes | [33] |
| Lipophilic G-quadruplex | - | 3 C12 spacers and 1 cholesterol on each strand | Switchable K+ transport | [35] |
| Single-duplex | - | 6 tetraphenyl porphyrins | Ion conduction | [64] |
| T-like | 3.7; 27 | 57 tocopherols at the bottom of the double-layered plate | 1. DNA translocation 2. Molecular transmembrane transport | [34] |
| Ligand-controlled | -; 9.0 ± 1.5 | 3 cholesterols | Controlled DNA-triggered and charge-selective release of small-molecule cargo (sulfo-rhodamine B, SRB) from a reservoir | [66] |
| Large, square | 10; 24 | 47 strands at the bottom of the nanopore, complementary to the anchor DNA on the phospholipid membrane | Selective transmembrane transport of molecules with different sizes (calcein, 40 k or 500 k dextran) | [69] |
| Reversibly gated | 20.4; - | 64 cholesterols | 1. Dye molecular (Atto633) transmembrane transport 2. Dynamically controlled cargo (Atto633 and GFP) transport across the lipid bilayer | [67] |
| Trigger-assembled | 0.8; 7.5 | One cholesterol per component (A or B) | Ion conduction | [70] |
| Tensegrity driven | -; 4 ± 0.2 | 4 cholesterols driven channels inserted into the membrane and 2 cholesterols grafted at the ends of two strands facing each other on one side of the barrel | Controlled transmembrane transport of SRB | [72] |
| Temperature-gated | 2; - | 4 cholesterols | Permitted temperature-controlled transport of molecular cargo (SRB) across the bilayer | [73] |
| Light-triggered synthetic | 2; 12.5 | 4 cholesterols | Light-gated small-molecule (SRB) transport | [75] |
| 8-helix bundle | 4; 14 | 4 TEG-cholesterols | 1. Depolarized plasma membrane to induce pyroptosis 2. Inhibited tumor growth | [83] |
| Protein-controlled | Molecular gate approximately 13 nm × 5 nm× 5 nm | 4 cholesterols | 1. Controlled transport of molecular cargo (SRB) across lipid bilayers 2. Released cytotoxic drug (topotecan) for cell killing | [84] |
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shen, H.; Wang, Y.; Wang, J.; Li, Z.; Yuan, Q. Emerging Biomimetic Applications of DNA Nanotechnology. ACS Appl. Mater. Interfaces 2019, 11, 13859–13873. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Xiong, Q.; Zhou, K.; Feng, Q.; Liu, L.; Tian, T.; Wu, C.; Xiong, Y.; Melia, T.J.; Lusk, C.P.; et al. Functionalized DNA-Origami-Protein Nanopores Generate Large Transmembrane Channels with Programmable Size-Selectivity. J. Am. Chem. Soc. 2022, 145, 1292–1300. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, L.; Liu, S.; Li, F.; Liu, J. Bioinspired artificial nanochannels: Construction and application. Mater. Chem. Front. 2021, 5, 1610–1631. [Google Scholar] [CrossRef]
- Pugh, G.C.; Burns, J.R.; Howorka, S. Comparing proteins and nucleic acids for next-generation biomolecular engineering. Nat. Rev. Chem. 2018, 2, 113–130. [Google Scholar] [CrossRef]
- Luo, Y.; Zhu, C.; Zhang, T.; Yan, T.; Liu, J. Self-assembled Supramolecular Artificial Transmembrane Ion Channels: Recent Progress and Application. Chem. Res. Chin. Univ. 2023, 39, 3–12. [Google Scholar] [CrossRef]
- Langecker, M.; Arnaut, V.; List, J.; Simmel, F.C. DNA nanostructures interacting with lipid bilayer membranes. Acc. Chem. Res. 2014, 47, 1807–1815. [Google Scholar] [CrossRef] [PubMed]
- Seeman, N.C.; Sleiman, H.F. DNA nanotechnology. Nat. Rev. Mater. 2017, 3, 17068. [Google Scholar] [CrossRef]
- Ramezani, H.; Dietz, H. Building machines with DNA molecules. Nat. Rev. Genet. 2020, 21, 5–26. [Google Scholar] [CrossRef]
- Suzuki, Y.; Endo, M.; Sugiyama, H. Mimicking Membrane-Related Biological Events by DNA Origami Nanotechnology. ACS Nano 2015, 9, 3418–3420. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, Y.; Liu, D. DNA nanochannels. F1000Res 2017, 6, 503. [Google Scholar] [CrossRef]
- Wu, N.; Chen, F.; Zhao, Y.; Yu, X.; Wei, J.; Zhao, Y. Functional and Biomimetic DNA Nanostructures on Lipid Membranes. Langmuir 2018, 34, 14721–14730. [Google Scholar] [CrossRef] [PubMed]
- Yin, P.; Hariadi, R.F.; Sahu, S.; Choi, H.M.T.; Park, S.H.; LaBean, T.H.; Reif, J.H. Programming DNA Tube Circumferences. Science 2008, 321, 824–826. [Google Scholar] [CrossRef] [PubMed]
- Frykholm, K.; Muller, V.; KK, S.; Dorfman, K.D.; Westerlund, F. DNA in nanochannels: Theory and applications. Q. Rev. Biophys. 2022, 55, e12. [Google Scholar] [CrossRef] [PubMed]
- Lanphere, C.; Offenbartl-Stiegert, D.; Dorey, A.; Pugh, G.; Georgiou, E.; Xing, Y.; Burns, J.R.; Howorka, S. Design, assembly, and characterization of membrane-spanning DNA nanopores. Nat. Protoc. 2021, 16, 86–130. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, Y.; Liu, P.; Wang, L.; Lin, J.; Fan, C. Biomimetic DNA Nanotubes: Nanoscale Channel Design and Applications. Angew. Chem. Int. Ed. Engl. 2019, 58, 8996–9011. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Chen, L.; Gao, P.; Xia, F. Solid-state nanopore/channels meet DNA nanotechnology. Matter 2023, 6, 373–396. [Google Scholar] [CrossRef]
- Gopfrich, K.; Ohmann, A.; Keyser, U.F. Design and Assembly of Membrane-Spanning DNA Nanopores. Methods Mol. Biol. 2021, 2186, 33–48. [Google Scholar]
- Zhao, N.; Chen, Y.; Chen, G.; Xiao, Z. Artificial Cells Based on DNA Nanotechnology. ACS Appl. Bio Mater. 2020, 3, 3928–3934. [Google Scholar] [CrossRef]
- Niranjan, D.N.; Thiyagarajan, D.; Bhatia, D. DNA Origami in the Quest for Membrane Piercing. Chem. Asian. J. 2022, 17, e202200591. [Google Scholar] [CrossRef]
- Chen, J.; Seeman, N.C. Synthesis from DNA of a molecule with the connectivity of a cube. Nature 1991, 350, 631–633. [Google Scholar] [CrossRef]
- Ohmann, A.; Li, C.Y.; Maffeo, C.; Nahas, K.A.; Baumann, K.N.; Gopfrich, K.; Yoo, J.; Keyser, U.F.; Aksimentiev, A. A synthetic enzyme built from DNA flips 107 lipids per second in biological membranes. Nat. Commun. 2018, 9, 2426. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.F.; Joshi, H.; Terry, S.J.; Burns, J.R.; Aksimentiev, A.; Eggert, U.S.; Howorka, S. Hydrophobic Interactions between DNA Duplexes and Synthetic and Biological Membranes. J. Am. Chem. Soc. 2021, 143, 8305–8313. [Google Scholar] [CrossRef] [PubMed]
- Chidchob, P.; Offenbartl-Stiegert, D.; McCarthy, D.; Luo, X.; Li, J.; Howorka, S.; Sleiman, H.F. Spatial Presentation of Cholesterol Units on a DNA Cube as a Determinant of Membrane Protein-Mimicking Functions. J. Am. Chem. Soc. 2018, 141, 1100–1108. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.R.; Howorka, S. Defined Bilayer Interactions of DNA Nanopores Revealed with a Nuclease-Based Nanoprobe Strategy. ACS Nano 2018, 12, 3263–3271. [Google Scholar] [CrossRef] [PubMed]
- Langecker, M.; Arnaut, V.; Martin, T.G.; List, J.; Renner, S.; Mayer, M.; Dietz, H.; Simmel, F.C. Synthetic lipid membrane channels formed by designed DNA nanostructures. Science 2012, 338, 932–936. [Google Scholar] [CrossRef]
- Gopfrich, K.; Li, C.Y.; Ricci, M.; Bhamidimarri, S.P.; Yoo, J.; Gyenes, B.; Ohmann, A.; Winterhalter, M.; Aksimentiev, A.; Keyser, U.F. Large-Conductance Transmembrane Porin Made from DNA Origami. ACS Nano 2016, 10, 8207–8214. [Google Scholar] [CrossRef]
- Diederichs, T.; Pugh, G.; Dorey, A.; Xing, Y.; Burns, J.R.; Nguyen, Q.H.; Tornow, M.; Tampe, R.; Howorka, S. Synthetic protein-conductive membrane nanopores built with DNA. Nat. Commun. 2019, 10, 5018. [Google Scholar] [CrossRef]
- Thomsen, R.P.; Malle, M.G.; Okholm, A.H.; Krishnan, S.; Bohr, S.S.; Sorensen, R.S.; Ries, O.; Vogel, S.; Simmel, F.C.; Hatzakis, N.S.; et al. A large size-selective DNA nanopore with sensing applications. Nat. Commun. 2019, 10, 5655. [Google Scholar] [CrossRef]
- Fragasso, A.; De Franceschi, N.; Stommer, P.; van der Sluis, E.O.; Dietz, H.; Dekker, C. Reconstitution of Ultrawide DNA Origami Pores in Liposomes for Transmembrane Transport of Macromolecules. ACS Nano 2021, 15, 12768–12779. [Google Scholar] [CrossRef]
- Xing, Y.; Dorey, A.; Jayasinghe, L.; Howorka, S. Highly shape- and size-tunable membrane nanopores made with DNA. Nat. Nanotechnol. 2022, 17, 708–713. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.R.; Stulz, E.; Howorka, S. Self-assembled DNA nanopores that span lipid bilayers. Nano Lett. 2013, 13, 2351–2356. [Google Scholar] [CrossRef]
- Burns, J.R.; Gopfrich, K.; Wood, J.W.; Thacker, V.V.; Stulz, E.; Keyser, U.F.; Howorka, S. Lipid-bilayer-spanning DNA nanopores with a bifunctional porphyrin anchor. Angew. Chem. Int. Ed. Engl. 2013, 52, 12069–12072. [Google Scholar] [CrossRef]
- Debnath, M.; Chakraborty, S.; Kumar, Y.P.; Chaudhuri, R.; Jana, B.; Dash, J. Ionophore constructed from non-covalent assembly of a G-quadruplex and liponucleoside transports K+-ion across biological membranes. Nat. Commun. 2020, 11, 469. [Google Scholar] [CrossRef]
- Krishnan, S.; Ziegler, D.; Dietz, H.; Simmel, F.C. Molecular transport through large-diameter DNA nanopores. Nat. Commun. 2016, 7, 12787. [Google Scholar] [CrossRef]
- Li, C.; Chen, H.; Zhou, L.; Shi, H.; He, X.; Yang, X.; Wang, K.; Liu, J. Single-stranded DNA designed lipophilic G-quadruplexes as transmembrane channels for switchable potassium transport. Chem. Commun. 2019, 55, 12004–12007. [Google Scholar] [CrossRef]
- Aldaye, F.A.; Lo, P.K.; Karam, P.; McLaughlin, C.K.; Cosa, G.; Sleiman, H.F. Modular construction of DNA nanotubes of tunable geometry and single- or double-stranded character. Nat. Nanotechnol. 2009, 4, 349–352. [Google Scholar] [CrossRef]
- Lo, P.K.; Karam, P.; Aldaye, F.A.; McLaughlin, C.K.; Hamblin, G.D.; Cosa, G.; Sleiman, H.F. Loading and selective release of cargo in DNA nanotubes with longitudinal variation. Nat. Chem. 2010, 2, 319–328. [Google Scholar] [CrossRef]
- Hamblin, G.D.; Carneiro, K.M.M.; Fakhoury, J.F.; Bujold, K.E.; Sleiman, H.F. Rolling Circle Amplification-Templated DNA Nanotubes Show Increased Stability and Cell Penetration Ability. J. Am. Chem. Soc. 2012, 134, 2888–2891. [Google Scholar] [CrossRef]
- Hariri, A.A.; Hamblin, G.D.; Gidi, Y.; Sleiman, H.F.; Cosa, G. Stepwise growth of surface-grafted DNA nanotubes visualized at the single-molecule level. Nat. Chem. 2015, 7, 295–300. [Google Scholar] [CrossRef]
- Rahbani, J.F.; Vengut-Climent, E.; Chidchob, P.; Gidi, Y.; Trinh, T.; Cosa, G.; Sleiman, H.F. DNA Nanotubes with Hydrophobic Environments: Toward New Platforms for Guest Encapsulation and Cellular Delivery. Adv. Health Mater. 2018, 7, 1701049. [Google Scholar] [CrossRef]
- Saliba, D.; Luo, X.; Rizzuto, F.J.; Sleiman, H.F. Programming rigidity into size-defined wireframe DNA nanotubes. Nanoscale 2023, 15, 5403–5413. [Google Scholar] [CrossRef]
- Yang, Q.; Guo, Z.; Liu, H.; Peng, R.; Xu, L.; Bi, C.; He, Y.; Liu, Q.; Tan, W. A Cascade Signaling Network between Artificial Cells Switching Activity of Synthetic Transmembrane Channels. J. Am. Chem. Soc. 2020, 143, 232–240. [Google Scholar] [CrossRef]
- Yoo, J.; Aksimentiev, A. Molecular Dynamics of Membrane-Spanning DNA Channels: Conductance Mechanism, Electro-Osmotic Transport, and Mechanical Gating. J. Phys. Chem. Lett. 2015, 6, 4680–4687. [Google Scholar] [CrossRef]
- Maingi, V.; Lelimousin, M.; Howorka, S.; Sansom, M.S.P. Gating-like Motions and Wall Porosity in a DNA Nanopore Scaffold Revealed by Molecular Simulations. ACS Nano 2015, 9, 11209–11217. [Google Scholar] [CrossRef] [PubMed]
- Maingi, V.; Burns, J.R.; Uusitalo, J.J.; Howorka, S.; Marrink, S.J.; Sansom, M.S. Stability and dynamics of membrane-spanning DNA nanopores. Nat. Commun. 2017, 8, 14784. [Google Scholar] [CrossRef] [PubMed]
- Birkholz, O.; Burns, J.R.; Richter, C.P.; Psathaki, O.E.; Howorka, S.; Piehler, J. Multi-functional DNA nanostructures that puncture and remodel lipid membranes into hybrid materials. Nat. Commun. 2018, 9, 1521. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Manda, S.; Park, Y.; Demir, B.; Sanchez, J.; Anantram, M.P.; Oren, E.E.; Gopinath, A.; Rolandi, M. DNA nanopores as artificial membrane channels for bioprotonics. Nat. Commun. 2023, 14, 5364. [Google Scholar] [CrossRef] [PubMed]
- Diederichs, T.; Ahmad, K.; Burns, J.R.; Nguyen, Q.H.; Siwy, Z.S.; Tornow, M.; Coveney, P.V.; Tampe, R.; Howorka, S. Principles of Small-Molecule Transport through Synthetic Nanopores. ACS Nano 2021, 15, 16194–16206. [Google Scholar] [CrossRef]
- Gopfrich, K.; Zettl, T.; Meijering, A.E.; Hernandez-Ainsa, S.; Kocabey, S.; Liedl, T.; Keyser, U.F. DNA-Tile Structures Induce Ionic Currents through Lipid Membranes. Nano Lett. 2015, 15, 3134–3138. [Google Scholar] [CrossRef]
- Mathieu, F.; Liao, S.; Kopatsch, J.; Wang, T.; Mao, C.; Seeman, N.C. Six-Helix Bundles Designed from DNA. Nano Lett. 2005, 5, 661–665. [Google Scholar] [CrossRef]
- Endo, M.; Seeman, N.C.; Majima, T. DNA Tube Structures Controlled by a Four-Way-Branched DNA Connector. Angew. Chem. Int. Ed. Engl. 2005, 44, 6074–6077. [Google Scholar] [CrossRef]
- Mohammed, A.M.; Šulc, P.; Zenk, J.; Schulman, R. Self-assembling DNA nanotubes to connect molecular landmarks. Nat. Nanotechnol. 2016, 12, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Green, L.N.; Subramanian, H.K.K.; Mardanlou, V.; Kim, J.; Hariadi, R.F.; Franco, E. Autonomous dynamic control of DNA nanostructure self-assembly. Nat. Chem. 2019, 11, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Franco, E. Enzyme-Driven Assembly and Disassembly of Hybrid DNA-RNA Nanotubes. J. Am. Chem. Soc. 2019, 141, 7831–7841. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Phua, S.C.; Nihongaki, Y.; Li, Y.; Pacella, M.; Li, Y.; Mohammed, A.M.; Sun, S.; Inoue, T.; Schulman, R. Growth and site-specific organization of micron-scale biomolecular devices on living mammalian cells. Nat. Commun. 2021, 12, 5729. [Google Scholar] [CrossRef]
- Dhanasekar, N.N.; Li, Y.; Schulman, R. The ion permeability of DNA nanotube channels. BioRxiv 2022, 1–33. [Google Scholar] [CrossRef]
- Li, Y.; Maffeo, C.; Schulman, R. Leakless end-to-end transport of small molecules through micron-length DNA nanochannels. Sci. Adv. 2022, 8, eabq4834. [Google Scholar] [CrossRef]
- Rothemund, P.W. Folding DNA to create nanoscale shapes and patterns. Nature 2006, 440, 297–302. [Google Scholar] [CrossRef]
- Forman, S.L.; Fettinger, J.C.; Pieraccini, S.; Gottarelli, G.; Davis, J.T. Toward Artificial Ion Channels: A Lipophilic G-Quadruplex. J. Am. Chem. Soc. 2000, 122, 4060–4067. [Google Scholar] [CrossRef]
- Lee, M.P.H.; Parkinson, G.N.; Hazel, P.; Neidle, S. Observation of the Coexistence of Sodium and Calcium Ions in a DNA G-Quadruplex Ion Channel. J. Am. Chem. Soc. 2007, 129, 10106–10107. [Google Scholar] [CrossRef]
- Akhshi, P.; Mosey, N.J.; Wu, G. Free-Energy Landscapes of Ion Movement through a G-Quadruplex DNA Channel. Angew. Chem. 2012, 124, 2904–2908. [Google Scholar] [CrossRef]
- Akhshi, P.; Wu, G. Umbrella sampling molecular dynamics simulations reveal concerted ion movement through G-quadruplex DNA channels. Phys. Chem. Chem. Phys. 2017, 19, 11017–11025. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, S.; Senapati, S. Dynamics and Barrier of Movements of Sodium and Potassium Ions Across the Oxytricha nova G-Quadruplex Core. J. Phys. Chem. B 2020, 124, 11055–11066. [Google Scholar] [CrossRef]
- Göpfrich, K.; Li, C.-Y.; Mames, I.; Bhamidimarri, S.P.; Ricci, M.; Yoo, J.; Mames, A.; Ohmann, A.; Winterhalter, M.; Stulz, E.; et al. Ion Channels Made from a Single Membrane-Spanning DNA Duplex. Nano Lett. 2016, 16, 4665–4669. [Google Scholar] [CrossRef]
- Morzy, D.; Joshi, H.; Sandler, S.E.; Aksimentiev, A.; Keyser, U.F. Membrane Activity of a DNA-Based Ion Channel Depends on the Stability of Its Double-Stranded Structure. Nano Lett. 2021, 21, 9789–9796. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.R.; Seifert, A.; Fertig, N.; Howorka, S. A biomimetic DNA-based channel for the ligand-controlled transport of charged molecular cargo across a biological membrane. Nat. Nanotechnol. 2016, 11, 152–156. [Google Scholar] [CrossRef]
- Dey, S.; Dorey, A.; Abraham, L.; Xing, Y.; Zhang, I.; Zhang, F.; Howorka, S.; Yan, H. A reversibly gated protein-transporting membrane channel made of DNA. Nat. Commun. 2022, 13, 2271. [Google Scholar] [CrossRef]
- Seifert, A.; Gopfrich, K.; Keyser, U.F.; Howorka, S. Bilayer-Spanning DNA Nanopores with Voltage-Switching between Open and Closed State. ACS Nano 2015, 9, 1117–1126. [Google Scholar] [CrossRef]
- Iwabuchi, S.; Kawamata, I.; Murata, S.; Nomura, S.M. A large, square-shaped, DNA origami nanopore with sealing function on a giant vesicle membrane. Chem. Commun. 2021, 57, 2990–2993. [Google Scholar] [CrossRef]
- Lanphere, C.; Ciccone, J.; Dorey, A.; Hagleitner-Ertugrul, N.; Knyazev, D.; Haider, S.; Howorka, S. Triggered Assembly of a DNA-Based Membrane Channel. J. Am. Chem. Soc. 2022, 144, 4333–4344. [Google Scholar] [CrossRef]
- Yang, L.; Cullin, C.; Elezgaray, J. Detection of Short DNA Sequences with DNA Nanopores. Chemphyschem 2022, 23, e202200021. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, O.; Calmet, P.; Alves, I.; Lecomte, S.; Raoux, M.; Cullin, C.; Elezgaray, J. A tensegrity driven DNA nanopore. Nanoscale 2017, 9, 9762–9769. [Google Scholar] [CrossRef] [PubMed]
- Arnott, P.M.; Howorka, S. A Temperature-Gated Nanovalve Self-Assembled from DNA to Control Molecular Transport across Membranes. ACS Nano 2019, 13, 3334–3340. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Xie, G.; Kong, X.Y.; Zhang, Z.; Xiao, K.; Wen, L.; Jiang, L. Light-Controlled Ion Transport through Biomimetic DNA-Based Channels. Angew. Chem. Int. Ed. Engl. 2016, 55, 15637–15641. [Google Scholar] [CrossRef]
- Offenbartl-Stiegert, D.; Rottensteiner, A.; Dorey, A.; Howorka, S. A Light-Triggered Synthetic Nanopore for Controlling Molecular Transport Across Biological Membranes. Angew. Chem. Int. Ed. Engl. 2022, 61, e202210886. [Google Scholar] [CrossRef]
- Li, C.; Chen, H.; Yang, X.; Wang, K.; Liu, J. An ion transport switch based on light-responsive conformation-dependent G-quadruplex transmembrane channels. Chem. Commun. 2021, 57, 8214–8217. [Google Scholar] [CrossRef]
- Li, C.; Chen, H.; Chen, Q.; Shi, H.; Yang, X.; Wang, K.; Liu, J. Lipophilic G-Quadruplex Isomers as Biomimetic Ion Channels for Conformation-Dependent Selective Transmembrane Transport. Anal. Chem. 2020, 92, 10169–10176. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Wu, G.; Wang, K.; Onel, B.; Sakai, S.; Shao, Y.; Yang, D. Molecular Recognition of the Hybrid-2 Human Telomeric G-Quadruplex by Epiberberine: Insights into Conversion of Telomeric G-Quadruplex Structures. Angew. Chem. Int. Ed. Engl. 2018, 57, 10888–10893. [Google Scholar] [CrossRef]
- Lu, H.; Li, S.; Chen, J.; Xia, J.; Zhang, J.; Huang, Y.; Liu, X.; Wu, H.; Zhao, Y.; Chaia, Z.; et al. Metal ions modulate the conformation and stability of a G-quadruplex with or without a small-molecule ligand. Metallomics 2015, 7, 1508–1514. [Google Scholar] [CrossRef]
- Messager, L.; Burns, J.R.; Kim, J.; Cecchin, D.; Hindley, J.; Pyne, A.L.; Gaitzsch, J.; Battaglia, G.; Howorka, S. Biomimetic Hybrid Nanocontainers with Selective Permeability. Angew. Chem. Int. Ed. Engl. 2016, 55, 11106–11109. [Google Scholar] [CrossRef]
- Burns, J.R.; Al-Juffali, N.; Janes, S.M.; Howorka, S. Membrane-spanning DNA nanopores with cytotoxic effect. Angew. Chem. Int. Ed. Engl. 2014, 53, 12466–12470. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Gu, X.; Li, H.; Zhao, Y.; Yang, D.; Yu, W.; Han, D.; Li, J.; Tan, W. Molecular Transport through a Biomimetic DNA Channel on Live Cell Membranes. ACS Nano 2020, 14, 14616–14626. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liang, S.; Chen, Y.; Wu, M.; Zhang, Y. Destructing the Plasma Membrane with Activatable Vesicular DNA Nanopores. ACS Appl. Mater. Interfaces 2020, 12, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Lanphere, C.; Arnott, P.M.; Jones, S.F.; Korlova, K.; Howorka, S. A Biomimetic DNA-Based Membrane Gate for Protein-Controlled Transport of Cytotoxic Drugs. Angew. Chem. Int. Ed. Engl. 2021, 60, 1903–1908. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Jian, M.; Chen, H.; Zhou, B.; Shi, H.; Yang, X.; Wang, K.; Liu, J. Design of Lipophilic Split Aptamers as Artificial Carriers for Transmembrane Transport of Adenosine Triphosphate. CCS Chem. 2021, 3, 144–153. [Google Scholar] [CrossRef]
- Veetil, A.T.; Chakraborty, K.; Xiao, K.; Minter, M.R.; Sisodia, S.S.; Krishnan, Y. Cell-targetable DNA nanocapsules for spatiotemporal release of caged bioactive small molecules. Nat. Nanotechnol. 2017, 12, 1183–1189. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Li, P.; Zhao, Z.; Kong, X.Y.; Zhang, Z.; Xiao, K.; Wang, H.; Wen, L.; Jiang, L. Bacteriorhodopsin-Inspired Light-Driven Artificial Molecule Motors for Transmembrane Mass Transportation. Angew. Chem. Int. Ed. Engl. 2018, 57, 16708–16712. [Google Scholar] [CrossRef]
- List, J.; Weber, M.; Simmel, F.C. Hydrophobic actuation of a DNA origami bilayer structure. Angew. Chem. Int. Ed. Engl. 2014, 53, 4236–4239. [Google Scholar] [CrossRef]
- Bennett, I.M.; Farfano, H.M.V.; Bogani, F.; Primak, A.; Liddell, P.A.; Otero, L.; Sereno, L.; Silber, J.J.; Moore, A.L.; Moore, T.A.; et al. Active transport of Ca2+ by an artificial photosynthetic membrane. Nature 2002, 420, 398–401. [Google Scholar] [CrossRef]
- Cui, C.; Chakraborty, K.; Tang, X.A.; Schoenfelt, K.Q.; Hoffman, A.; Blank, A.; McBeth, B.; Pulliam, N.; Reardon, C.A.; Kulkarni, S.A.; et al. A lysosome-targeted DNA nanodevice selectively targets macrophages to attenuate tumours. Nat. Nanotechnol. 2021, 16, 1394–1402. [Google Scholar] [CrossRef]
- Maffeo, C.; Quednau, L.; Wilson, J.; Aksimentiev, A. DNA double helix, a tiny electromotor. Nat. Nanotechnol. 2022, 18, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Guo, P.; Jin, D.; Peng, Y.; Sun, X.; Chen, Y.; Liu, X.; Chen, W.; Wang, W.; Yan, X.; et al. Enzyme-Powered Tubular Microrobotic Jets as Bioinspired Micropumps for Active Transmembrane Drug Transport. ACS Nano 2023, 17, 5095–5107. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yu, G.; Sessler, J.L.; Shin, I.; Gale, P.A.; Huang, F. Artificial transmembrane ion transporters as potential therapeutics. Chem 2021, 7, 3256–3291. [Google Scholar] [CrossRef]
- Kanner, S.A.; Shuja, Z.; Choudhury, P.; Jain, A.; Colecraft, H.M. Targeted deubiquitination rescues distinct trafficking-deficient ion channelopathies. Nat. Methods 2020, 17, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Rastergar, A.; Soleimani, M. Hypokalaemia and hyperkalaemia. Postgrad. Med. J. 2001, 77, 759–764. [Google Scholar] [CrossRef]
- Curran, J.; Mohler, P.J. Alternative paradigms for ion channelopathies: Disorders of ion channel membrane trafficking and posttranslational modification. Annu. Rev. Physiol. 2015, 77, 505–524. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, W.; Chen, H.; Li, Y.; Liu, S.; Wang, K.; Liu, J. Design of DNA-Based Artificial Transmembrane Channels for Biosensing and Biomedical Applications. Chemosensors 2023, 11, 508. https://doi.org/10.3390/chemosensors11090508
Xu W, Chen H, Li Y, Liu S, Wang K, Liu J. Design of DNA-Based Artificial Transmembrane Channels for Biosensing and Biomedical Applications. Chemosensors. 2023; 11(9):508. https://doi.org/10.3390/chemosensors11090508
Chicago/Turabian StyleXu, Wanyu, Hui Chen, Yang Li, Shuangna Liu, Kemin Wang, and Jianbo Liu. 2023. "Design of DNA-Based Artificial Transmembrane Channels for Biosensing and Biomedical Applications" Chemosensors 11, no. 9: 508. https://doi.org/10.3390/chemosensors11090508
APA StyleXu, W., Chen, H., Li, Y., Liu, S., Wang, K., & Liu, J. (2023). Design of DNA-Based Artificial Transmembrane Channels for Biosensing and Biomedical Applications. Chemosensors, 11(9), 508. https://doi.org/10.3390/chemosensors11090508