
Monitoring of Lactate in Interstitial Fluid, Saliva and Sweat by Electrochemical Biosensor: The Uncertainties of Biological Interpretation



Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
2.2. Preparation and Use of Electrodes
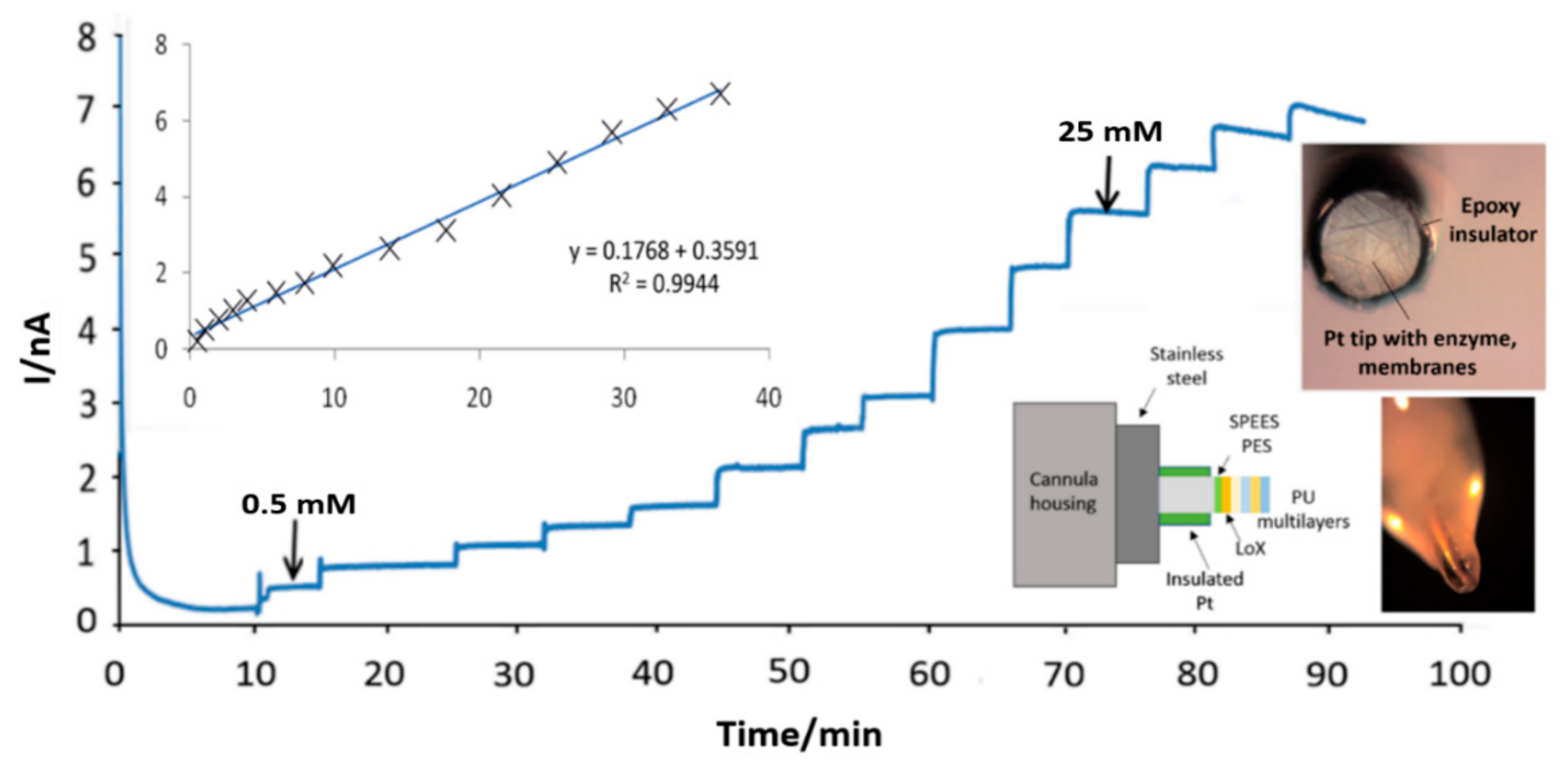
2.3. Physiological Measurement
3. Results and Discussion
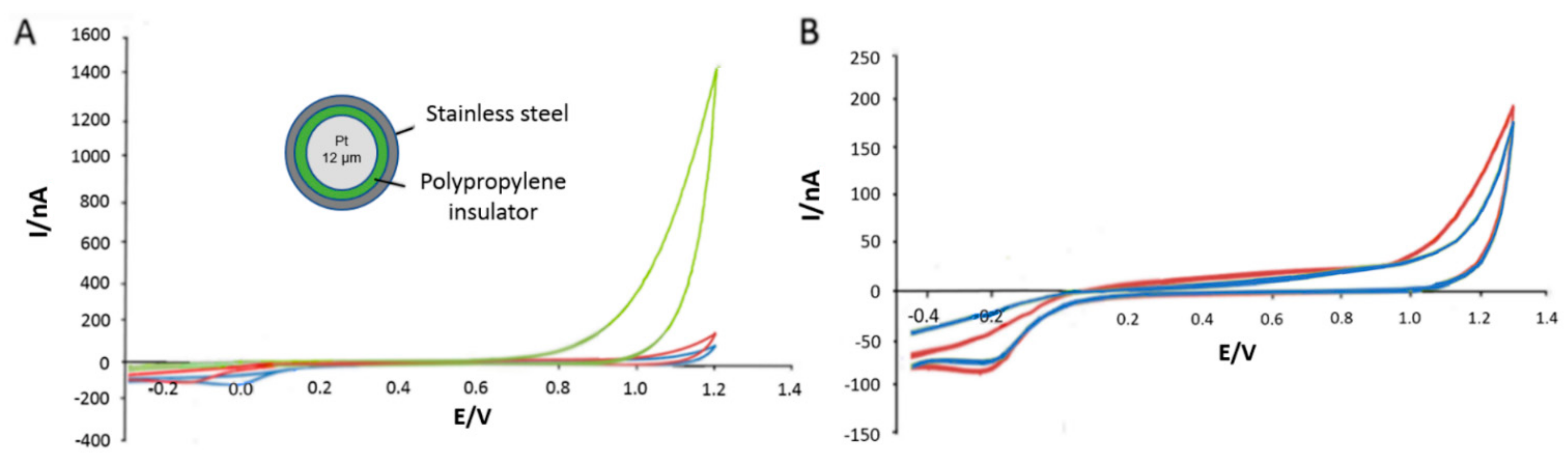
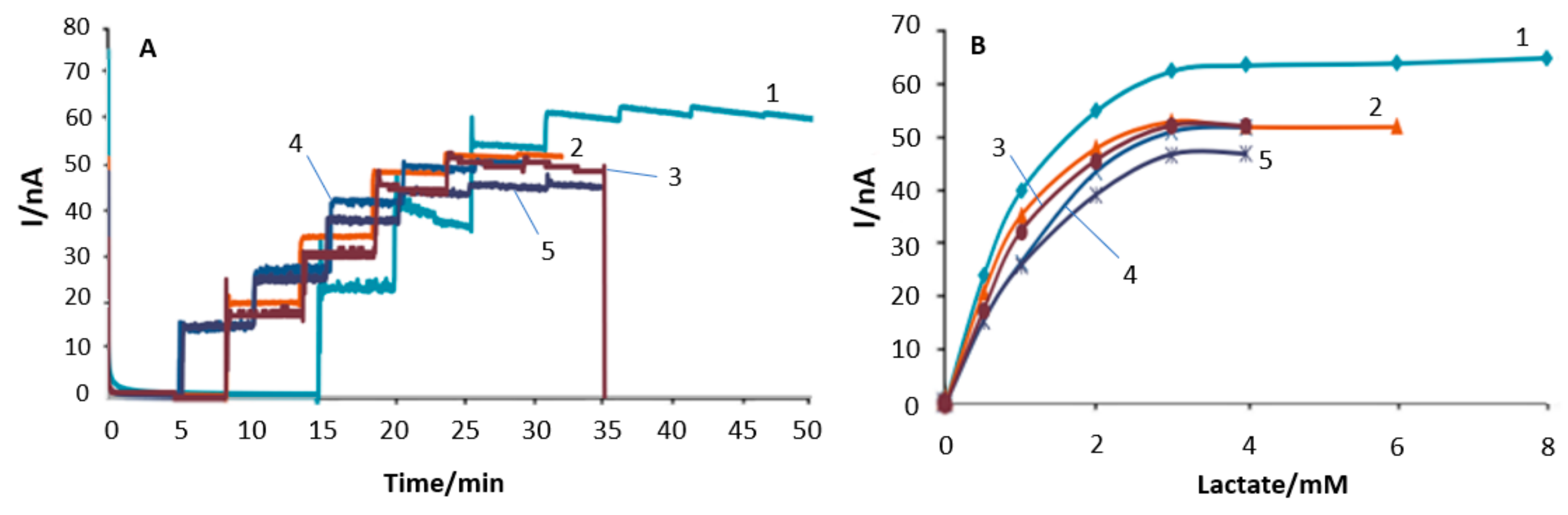
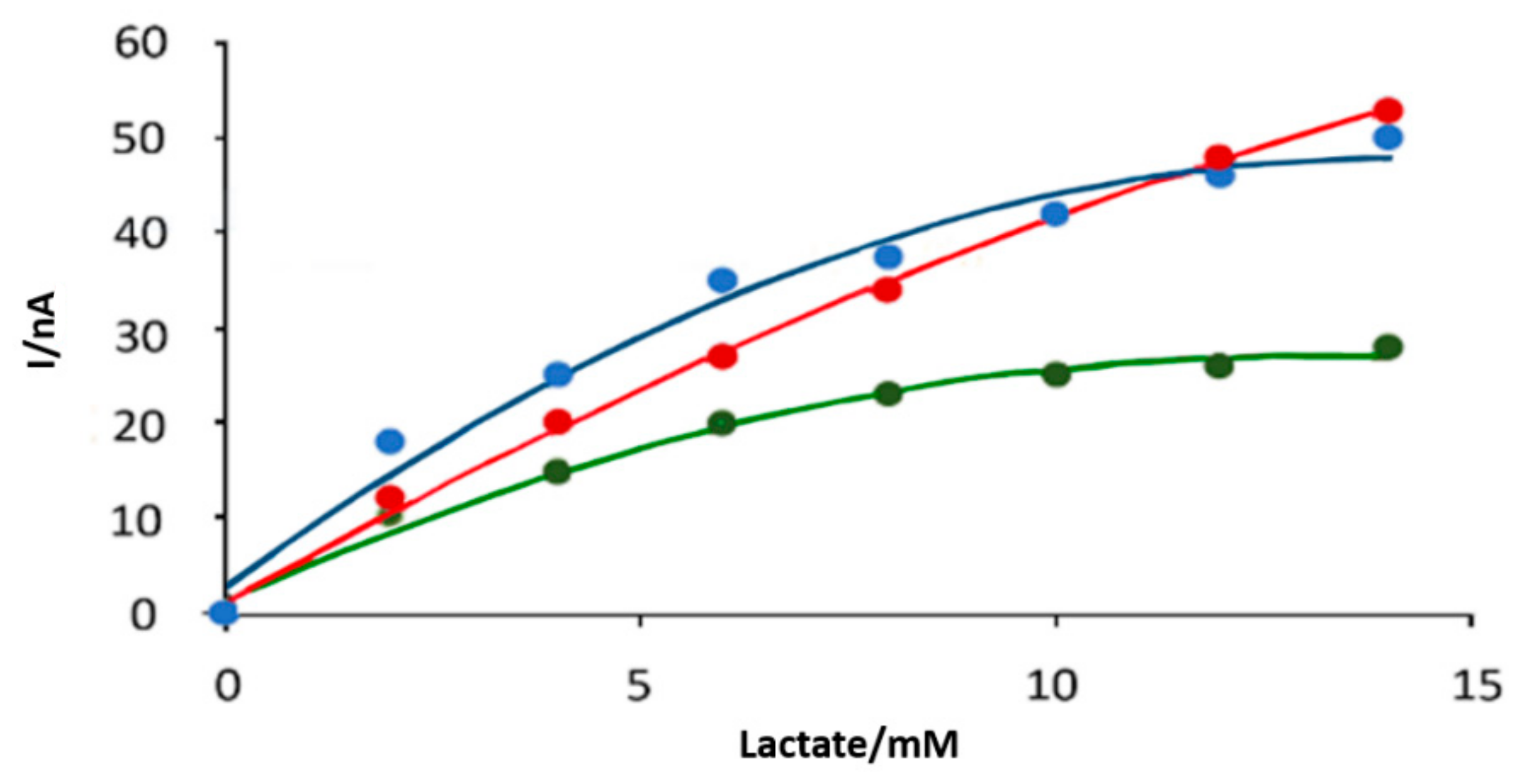
3.1. In Vitro Optimisation of Needle Electrodes
3.2. Polymer Barrier Membrane Effects on Sensors
3.3. Sensor pO2 Dependence
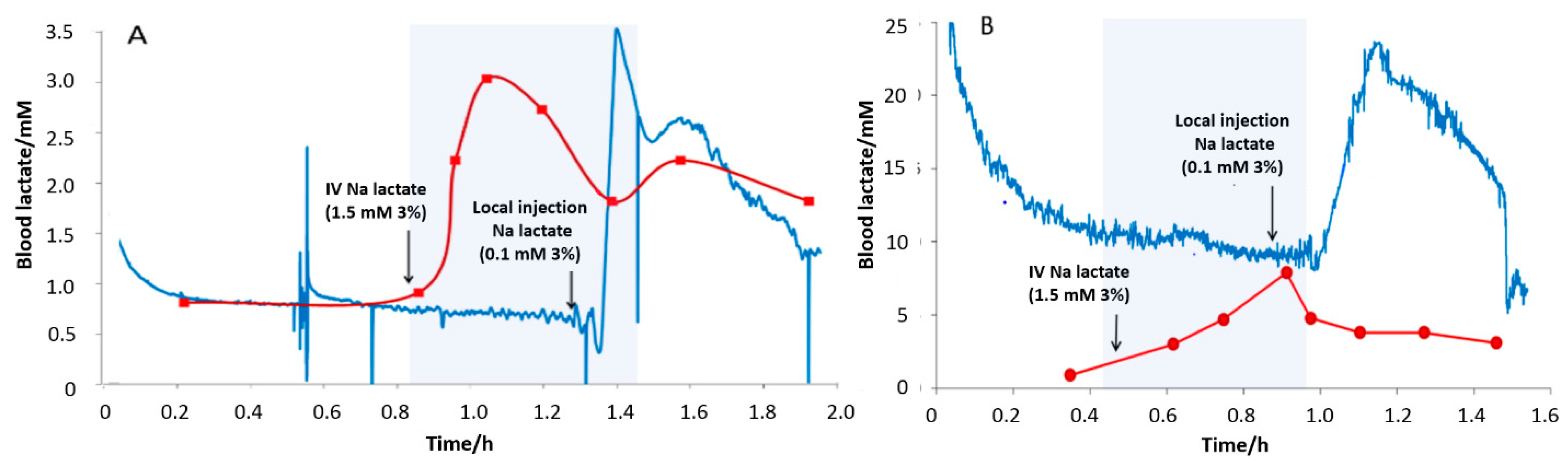
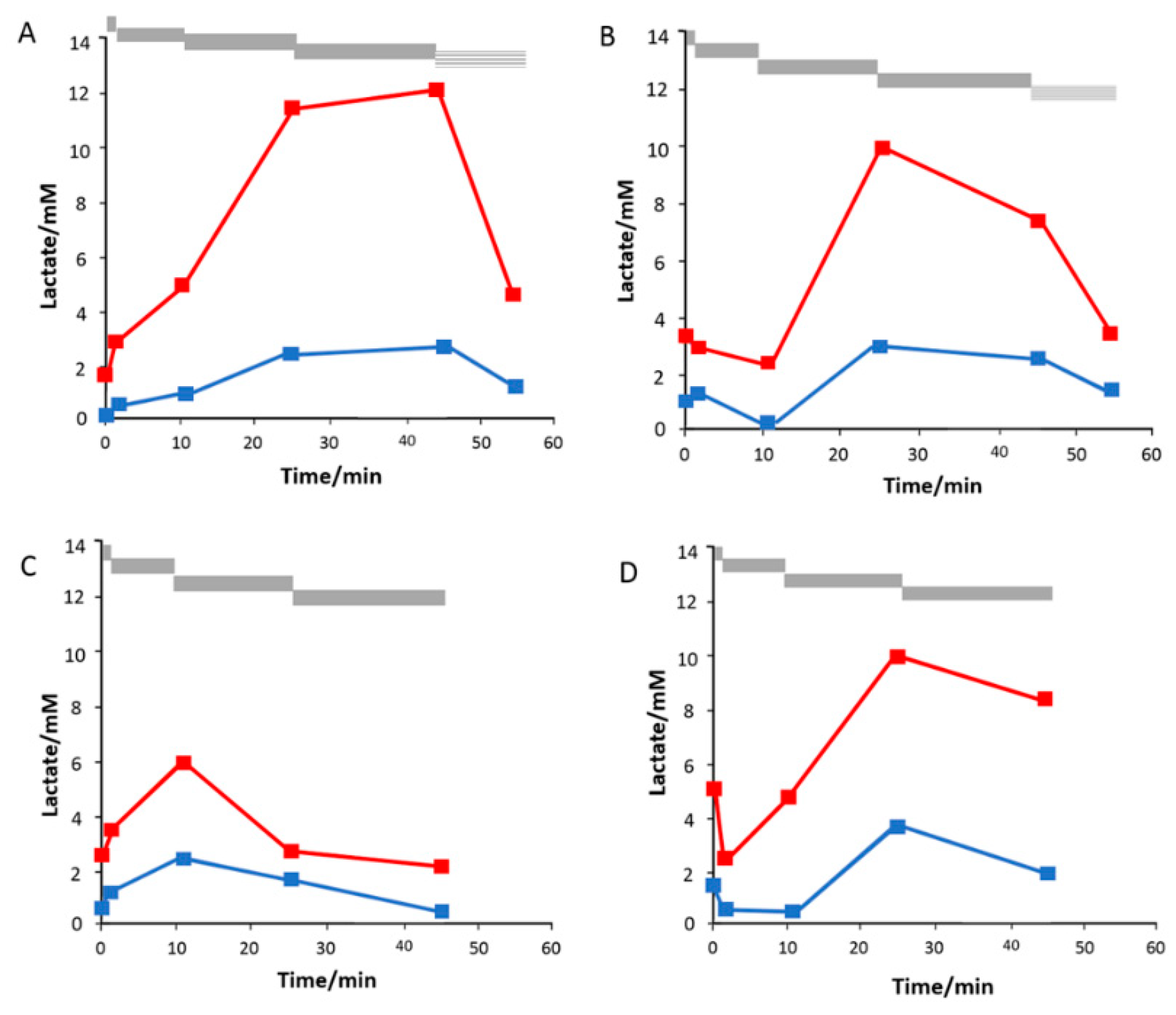
3.4. Subcutaneous Tissue Lactate Monitoring in the Rat
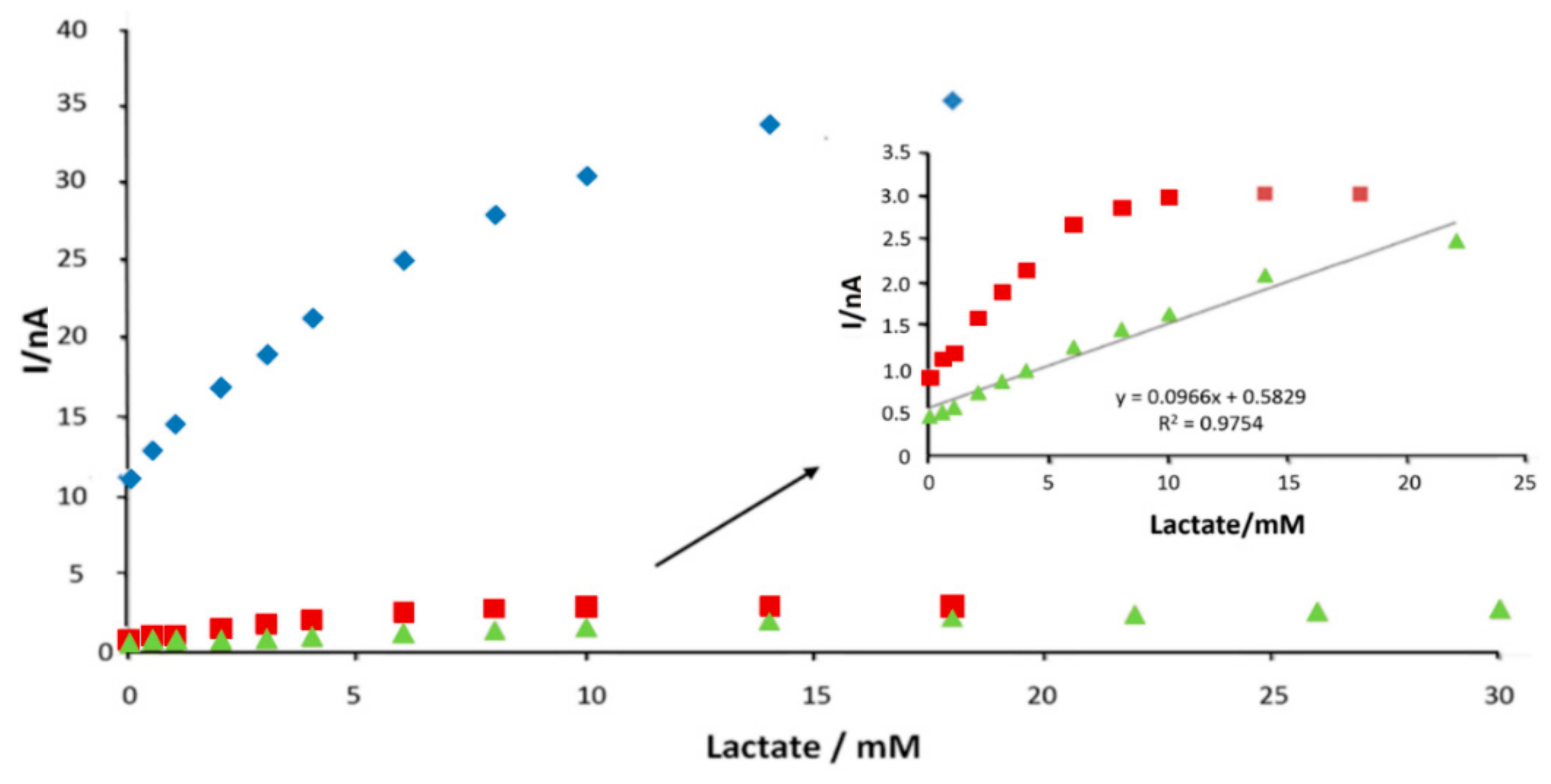
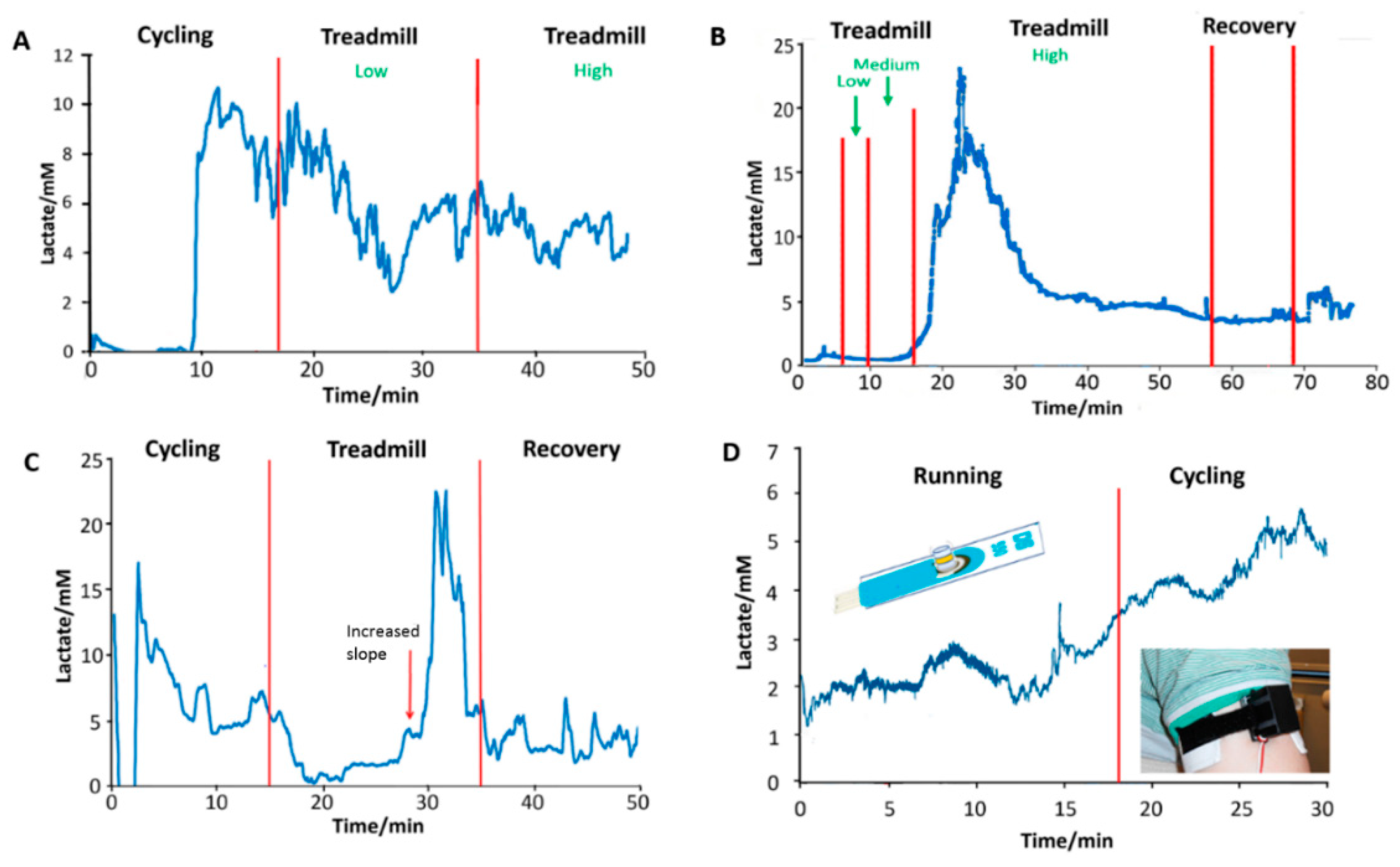
3.5. Saliva Measurement in Exercise
3.6. Sweat Monitoring in Exercise
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Adeva-Andany, M.; Lopez-Ojen, M.; Funcasta-Calderon, R.; Ameneiros-Rodriguez, E.; Donapetry-Garcia, C.; Vila-Altesor, M.; Rodriguez-Seijas, J. Comprehensive review on lactate metabolism in human health. Mitochondrion 2014, 17, 76–100. [Google Scholar] [CrossRef]
- Weiss, I.K.; Fink, S.; Harrison, R.; Feldman, J.D.; Brill, J.E. Clinical use of continuous arterial blood gas monitoring in the pediatric intensive care unit. Pediatrics 1999, 103, 440–445. [Google Scholar] [CrossRef]
- Brooks, G.A. Cell-cell and intracellular lactate shuttles. J. Physiol. 2009, 587, 5591–5600. [Google Scholar] [CrossRef]
- Brinkmann, B.; Fechner, G.; Karger, B.; DuChesne, A. Ketoacidosis and lactic acidosis—Frequent causes of death in chronic alcoholics? Int. J. Legal Med. 1998, 111, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Handy, J.M.; Soni, N. Physiological effects of hyperchloraemia and acidosis. Br. J. Anaesth. 2008, 101, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, M.A.; Williams, P.E.; Cherrington, A.D. Effect of a mixed meal on hepatic lactate and glucogneogenic precursor metabolism in dogs. Am. J. Physiol. 1984, 247, E362–E369. [Google Scholar] [CrossRef] [PubMed]
- Levy, B. Lactate and shock state: The metabolic view. Curr. Opin. Crit. Care 2006, 12, 315–321. [Google Scholar] [CrossRef]
- Crane, B.C.; Barwell, N.P.; Gopal, P.; Gopichand, M.; Higgs, T.; James, T.D.; Jones, C.M.; Mackenzie, A.; Mulavisala, K.P.; Paterson, W. The development of a continuous intravascular glucose monitoring sensor. J. Diabetes Sci. Technol. 2015, 9, 751–761. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Doloff, J.C.; Yesilyurt, V.; Sadraei, A.; McGarrigle, J.J.; Commis, M.; Veiseh, O.; Farah, S.; Isa, D.; Ghanis, S.; et al. Reduction of measurement noise in a continuous glucose monitor by coating the sensor with a zwitterionic polymer. Nat. Biomed. Eng. 2018, 2, 894–906. [Google Scholar] [CrossRef]
- Rigby, G.P.; Ahmed, S.; Horseman, G.; Vadgama, P. In vivo glucose monitoring with open microflow—Influences of fluid composition and preliminary evaluation in man. Anal. Chim. Acta 1999, 385, 23–32. [Google Scholar] [CrossRef]
- Chen, C.; Zhao, X.L.; Li, Z.H.; Zhu, Z.G.; Qian, S.H.; Flewitt, A.J. Current and emerging technology for continuous glucose monitoring. Sensors 2017, 17, 182. [Google Scholar] [CrossRef] [Green Version]
- Hoss, U.; Budiman, E.S.; Liu, H.; Christiansen, M.P. Continuous glucose monitoring in the subcutaneous tissue over a 14-day sensor wear period. J. Diabetes Sci. Technol. 2013, 7, 1210–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spehar-Deleze, A.M.; Anastasova, S.; Vadgama, P. Electropolymerised phenolic films as internal barriers for oxidase enzyme biosensors. Electroanalysis 2014, 26, 1335–1344. [Google Scholar] [CrossRef]
- Spehar-Deleze, A.; Anastasova, S.; Popplewell, J.; Vadgama, P. Extreme physiological state: Development of tissue lactate sensor. In Proceedings of the 2012 Ninth International Conference on Wearable and Implantable Body Sensor Networks, London, UK, 9–12 May 2012; IEEE: Piscataway, NJ, USA, 2012; pp. 17–21. [Google Scholar] [CrossRef]
- Nightingale, A.M.; Leong, C.L.; Burnish, R.A.; Hassan, S.U.; Zhang, Y.; Clough, G.F.; Boutelle, M.G.; Voegeli, D.; Niu, X.Z. Monitoring biomolecule concentrations in tissue using a wearable droplet microfluidic-based sensor. Nat. Commun. 2019, 10, 2741. [Google Scholar] [CrossRef] [Green Version]
- Gowers, S.A.N.; Curto, V.F.; Seneci, C.A.; Wang, C.; Anastasova, S.; Vadgama, P.; Yang, G.Z.; Boutelle, M.G. 3D printed microfluidic device with integrated biosensors for online analysis of subcutaneous human microdialysate. Anal. Chem. 2015, 87, 7763–7770. [Google Scholar] [CrossRef] [PubMed]
- Kastellorizios, M.; Burgess, D. Continuous Metabolic Monitoring Based on Multi-Analyte Biomarkers to Predict Exhaustion. Sci. Rep. 2015, 5, 10603. [Google Scholar] [CrossRef] [Green Version]
- Sardesai, N.P.; Ganesana, M.; Karimi, A.; Leiter, J.C.; Andreescu, S. Platinum-Doped ceria based biosensor for in vitro and in vivo monitoring of lactate during hypoxia. Anal. Chem. 2015, 87, 2996–3003. [Google Scholar] [CrossRef] [PubMed]
- Poscia, A.; Messeri, D.; Moscone, D.; Ricci, F.; Valgimigli, F. A novel continuous subcutaneous lactate monitoring system. Biosens. Bioelectron. 2005, 20, 2244–2250. [Google Scholar] [CrossRef] [Green Version]
- Ellmerer, M.; Schaupp, L.; Trajanoski, Z.; Jobst, G.; Moser, I.; Urban, G.; Skrabal, F.; Wach, P. Continuous measurement of subcutaneous lactate concentration during exercise by combining open-flow microperfusion and thin-film lactate sensors. Biosens. Bioelectron. 1998, 13, 1007–1013. [Google Scholar] [CrossRef]
- Ellmerer, M.; Schaupp, L.; Sendlhofer, G.; Wutte, A.; Brunner, G.A.; Trajanoski, Z.; Skrabal, F.; Wach, P.; Pieber, T.R. Lactate metabolism of subcutaneous adipose tissue studied by open flow microperfusion. J. Clin. Endocr. Metab. 1998, 83, 4394–4401. [Google Scholar] [CrossRef]
- Wolf, A.; Renehan, K.; Ho, K.K.Y.; Carr, B.D.; Chen, C.V.; Cornell, M.S.; Ye, M.Y.; Rojas-Pena, A.; Chen, H. Evaluation of continuous lactate monitoring systems within a heparinized in vivo porcine model intravenously and subcutaneously. Biosensors 2018, 8, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gargiulo, S.; Gramanzini, M.; Liuzzi, R.; Greco, A.; Brunetti, A.; Vesce, G. Effects of some anesthetic agents on skin microcirculation evaluated by laser Doppler perfusion imaging in mice. BMC Vet. Res. 2013, 9, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.B.; Zhang, Y.N.; Wilson, G.S. A needle-type enzyme-based lactate sensor for in-vivo monitoring. Anal. Chim. Acta 1993, 281, 503–511. [Google Scholar] [CrossRef]
- Pfeiffer, D.; Moller, B.; Klimes, N.; Szeponik, J. Amperometric lactate oxidase catheter for real-time lactate monitoring based on thin film technology. Biosens. Bioelectron. 1997, 12, 539–550. [Google Scholar] [CrossRef]
- Ward, W.K.; House, J.L.; Birck, J.; Anderson, E.M.; Jansen, L.B. A wire-based dual-analyte sensor for glucose and lactate: In vitro and in vivo evaluation. Diabetes Technol. Ther. 2004, 6, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Rong, Z.M.; Leitao, E.; Popplewell, J.; Alp, B.; Vadgama, P. Needle enzyme electrode for lactate measurement in vivo. IEEE Sens. J. 2008, 8, 113–120. [Google Scholar] [CrossRef]
- Rigby, G.P.; Crump, P.W.; Vadgama, P. Stabilized needle electrode system for in vivo glucose monitoring based on open flow microperfusion. Analyst 1996, 121, 871–875. [Google Scholar] [CrossRef]
- McGarraugh, G. The chemistry of commercial continuous glucose monitors. Diabetes Technol. Ther. 2009, 11, S17–S24. [Google Scholar] [CrossRef]
- Jackson, W.F.; Duling, B.R. Toxic effects of silver-silver chloride electrodes on vascular smooth muscle. Circ. Res. 1983, 53, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Kraft, C.N.; Hansis, M.; Arens, S.; Menger, M.D.; Vollmar, B. Striated muscle microvascular response to silver implants: A comparative in vivo study with titanium and stainless steel. J. Biomed. Mater. Res. 2000, 49, 192–199. [Google Scholar] [CrossRef]
- Kaiser, J.P.; Roesslein, M.; Diener, L.; Wichser, A.; Nowack, B.; Wick, P. Cytotoxic effects of nanosilver are highly dependent on the chloride concentration and the presence of organic compounds in the cell culture media. J. Nanobiotechnol. 2017, 15, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taurino, I.; Reiss, R.; Richter, M.; Fairhead, M.; Thony-Meyer, L.; De Micheli, G.; Carrara, S. Comparative study of three lactate oxidases from Aerococcus viridans for biosensing applications. Electrochim. Acta 2013, 93, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Yorita, K.; Matsuoka, T.; Misaki, H.; Massey, V. Interaction of two arginine residues in lactate oxidase with the enzyme flavin: Conversion of FMN to 8-formyl-FMN. Proc. Natl. Acad. Sci. USA 2000, 97, 13039–13044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.S.; Lian, J.Z.; Xue, H.L.; Jiang, Y.Q.; Wu, M.B.; Lin, J.P.; Yang, L.R. Enzymatic preparation of pyruvate by a whole-cell biocatalyst coexpressing L-lactate oxidase and catalase. Process Biochem. 2020, 96, 113–121. [Google Scholar] [CrossRef]
- Unterweger, B.; Stoisser, T.; Leitgeb, S.; Birner-Grunberger, R.; Nidetzky, B. Engineering of Aerococcus viridans L-Lactate Oxidase for Site-Specific PEGylation: Characterization and Selective Bioorthogonal Modification of a S218C Mutant. Bioconj. Chem. 2012, 23, 1406–1414. [Google Scholar] [CrossRef] [PubMed]
- Hiraka, K.; Kojima, K.; Tsugawa, W.; Asano, R.; Ikebukuro, K.; Sode, K. Rational engineering of Aerococcus viridans L-lactate oxidase for the mediator modification to achieve quasi-direct electron transfer type lactate sensor. Biosens. Bioelectron. 2020, 151, 111974. [Google Scholar] [CrossRef]
- Stoisser, T.; Rainer, D.; Leitgeb, S.; Wilson, D.K.; Nidetzky, B. The Ala95-to-Gly substitution in Aerococcus viridans l-lactate oxidase revisited—Structural consequences at the catalytic site and effect on reactivity with O2 and other electron acceptors. FEBS J. 2015, 282, 562–578. [Google Scholar] [CrossRef] [PubMed]
- Ashok, Y.; Maksimainen, M.M.; Kallio, T.; Kilpelainen, P.; Lehtio, L. FMN-dependent oligomerization of putative lactate oxidase from Pediococcus acidilactici. PLoS ONE 2020, 15, e0223870. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.M.; An, W.S. New clinical criteria for septic shock: Serum lactate level as new emerging vital sign. J. Thorac. Dis. 2016, 8, 1388–1390. [Google Scholar] [CrossRef]
- Reddy, S.M.; Vadgama, P.M. A study of the permeability properties of surfactant modified poly(vinyl chloride) membranes. Anal. Chim. Acta 1997, 350, 67–76. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, G.; Pumera, M. Beyond platinum: Bubble-propelled micromotors based on Ag and MnO2 catalysts. JACS 2014, 136, 2719–2722. [Google Scholar] [CrossRef] [PubMed]
- Anastasova, S.; Spehar-Deleze, A.M.; Kwasnicki, R.M.; Yang, G.Z.; Vadgama, P. Electrochemical monitoring of subcutaneous tissue pO(2) fluctuations during exercise using a semi-implantable needle electrode. Electroanalysis 2020, 32, 2393–2403. [Google Scholar] [CrossRef]
- Anastasova, A.; Spehar-Deleze, A.-M.; Bickham, D.; Uebel, P.; Schmidt, M.; Russell, P.; Vadgama, P. Stabilised biosensing using needle-based recess electrodes. Electroanalysis 2012, 24, 529–538. [Google Scholar] [CrossRef]
- Valenza, F.; Pizzocri, M.; Salice, V.; Chevallard, G.; Fossali, T.; Coppola, S.; Froio, S.; Polli, F.; Gatti, S.; Fortunato, F.; et al. Sodium bicarbonate treatment during transient or sustained lactic acidemia in normoxic and normotensive rats. PLoS ONE 2012, 7, e46035. [Google Scholar] [CrossRef]
- Adatia, K.; Raja, M.; Vadgama, P. An electrochemical study of microporous track-etched membrane permeability and the effect of surface protein layers. Colloid Surf. B 2017, 158, 84–92. [Google Scholar] [CrossRef]
- Kandimalla, K.; Kanikkannan, N.; Andega, S.; Singh, M. Effect of fatty acids on the permeation of melatonin across rat and pig skin in-vitro and on the transepidermal water loss in rats in-vivo. J. Pharm. Pharmacol. 1999, 51, 783–790. [Google Scholar] [CrossRef]
- Stace, T.M.; Damiano, E.R. An electrochemical model of the transport of charged molecules through the capillary glycocalyx. Biophys. J. 2001, 80, 1670–1690. [Google Scholar] [CrossRef] [Green Version]
- Rosengren, B.I.; Rippe, B.; Tenstad, O.; Wiig, H. Acute peritoneal dialysis in rats results in a marked reduction of interstitial colloid osmotic pressure. J. Am. Soc. Nephrol. 2004, 15, 3111–3116. [Google Scholar] [CrossRef] [Green Version]
- Ellmerer, M.; Haluzik, M.; Blaha, J.; Kremen, J.; Svacina, S.; Plasnik, A.; Ikeoka, D.; Bodenlenz, M.; Schaupp, L.; Plank, J.; et al. Clinical evaluation of subcutaneous lactate measurement in patients after major cardiac surgery. Int. J. Endocrinol. 2009, 2009, 390975. [Google Scholar] [CrossRef] [Green Version]
- Den Heuvel, I.; Vlasselaers, D.; Wouters, P.J.; Milants, I.; Ellger, B.; Vanhorebeek, I.; Van den Berghe, G. Serial lactate measurements using microdialysis of interstitial fluid do not correlate with plasma lactate in children after cardiac surgery. Pediatr. Crit. Care Med. 2009, 10, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Kopterides, P.; Theodorakopoulou, M.; Ilias, I.; Nikitas, N.; Frantzeskaki, F.; Vassiliadi, D.A.; Armaganidis, A.; Dimopoulou, I. Interrelationship between blood and tissue lactate in a general intensive care unit: A subcutaneous adipose tissue microdialysis study on 162 critically ill patients. J. Crit. Care 2012, 27, 742.e9–742.e18. [Google Scholar] [CrossRef]
- Kaptein, W.A.; Zwaagstra, J.J.; Venema, K.; Korf, J. Continuous ultraslow microdialysis and ultrafiltration for subcutaneous sampling as demonstrated by glucose and lactate measurements in rats. Anal. Chem. 1998, 70, 4696–4700. [Google Scholar] [CrossRef]
- Wren-Dail, M.A.; Dauchy, R.; Blask, D.E.; Hill, S.M.; Ooms, T.G.; Dupepe, M.; Bohm, J.; Rudolf, P. Effect of isoflurane anaesthesia on circadian metabolism and physiology in rats. Comp. Med. 2017, 67, 138–146. [Google Scholar] [PubMed]
- Oliveira, L.S.; Oliviera, S.F.; Manchado-Gobatto, F.B.; Costa, M.C. Salivary and blood lactate kinetics in response to maximal workload on cycle ergometer. Rev. Bras. Cineantropom. Desempenho Hum. 2015, 17, 565–574. [Google Scholar] [CrossRef]
- Tekus, E.; Kaj, M.; Szabo, E.; Szenasi, N.L.; Kerepesi, I.; Figler, M.; Gabriel, R.; Wilhelm, M. Comparison of blood and saliva lactate level after maximum intensity exercise. Acta Biol. Hung. 2012, 63, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Bocanegra, O.L.; Diaz, M.M.; Teixeira, R.R.; Soares, S.S.; Espindola, F.S. Determination of the lactate threshold by means of salivary biomarkers: Chromogranin A as novel marker of exercise intensity. Eur. J. Appl. Physiol. 2012, 112, 3195–3203. [Google Scholar] [CrossRef]
- Meleti, M.; Quartieri, E.; Antonelli, R.; Pezzi, M.E.; Ghezzi, B.; Viani, M.V.; Setti, G.; Casali, E.; Ferrari, E.; Ciociola, T.; et al. Metabolic profiles of whole, parotid and submandibular/sublingual saliva. Metabolites 2020, 10, 318. [Google Scholar] [CrossRef] [PubMed]
- Enomoto, K.; Shimizu, R.; Kudo, H. Real-time skin lactic acid monitoring system for assessment of training intensity. Electr. Commun. Jpn. 2018, 101, 41–46. [Google Scholar] [CrossRef]
- Gao, W.; Emaminejad, S.; Nyein, H.Y.Y.; Challa, S.; Chen, K.V.; Peck, A.; Fahad, H.M.; Ota, H.; Shiraki, H.; Kiriya, D.; et al. Fully integrated wearable sensor arrays for multiplexed in situ perspiration analysis. Nature 2016, 529, 509–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, W.Z.; Bandodkar, A.J.; Valdes-Ramirez, G.; Windmiller, J.R.; Yang, Z.J.; Ramirez, J.; Chan, G.; Wang, J. Electrochemical tattoo biosensors for real-time noninvasive lactate monitoring in human perspiration. Anal. Chem. 2013, 85, 6553–6560. [Google Scholar] [CrossRef] [PubMed]
- Baker, L.B.; Wolfe, A.S. Physiological mechanisms determining eccrine sweat composition. Eur. J. Appl. Physiol. 2020, 120, 719–752. [Google Scholar] [CrossRef] [Green Version]
- Green, J.M.; Bishop, P.A.; Muir, I.H.; McLester, J.R.; Heath, H.E. Effects of high and low blood lactate concentrations on sweat lactate response. Int. J. Sports Med. 2000, 21, 556–560. [Google Scholar] [CrossRef]
- Baker, L.B. Sweating Rate and Sweat Sodium Concentration in Athletes: A Review of Methodology and Intra/Interindividual Variability. Sports Med. 2017, 47, S111–S128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vairo, D.; Bruzzese, L.; Marlinge, M.; Fuster, L.; Adjriou, N.; Kipson, N.; Brunet, P.; Cautela, J.; Jammes, Y.; Mottola, G.; et al. Towards addressing the body electrolyte environment via sweat analysis: Pilocarpine iontophoresis supports assessment of plasma potassium concentration. Sci. Rep. 2017, 7, 11801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parilla, M.; Ortiz-Gomez, I.; Canovas, R.; Salinas-Castillo, A.; Cuartero, M.; Crespo, G.A. Potentiometric ion patch for on-body electrolyte monitoring in sweat: Toward a validation strategy to ensure physiological relevance. Anal. Chem. 2019, 91, 8644–8651. [Google Scholar] [CrossRef] [Green Version]
- Green, J.M.; Pritchett, R.C.; Tucker, D.C.; Crews, T.R.; McLester, J.R. Sweat lactate response during cycling at 30 °C and 18 °C WBGT. J. Sports Sci. 2004, 22, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Karpova, E.V.; Laptev, A.I.; Andreev, E.A.; Karyakina, E.E.; Karyakin, A.A. Relationship between sweat and blood lactate levels during exhaustive physical exercise. ChemElectroChem 2020, 7, 191–194. [Google Scholar] [CrossRef]
- Gordon, R.S.; Thompson, R.H.; Muenzer, J.; Thrasher, D. Sweat lactate in man is derived from blood glucose. J. Appl. Physiol. 1971, 31, 713–716. [Google Scholar] [CrossRef]
- Sakharov, D.A.; Shkurnikov, M.U.; Vagin, M.Y.; Yashina, E.I.; Karyakin, A.A.; Tonevitsky, A.G. Relationship between Lactate Concentrations in Active Muscle Sweat and Whole Blood. Bull. Exp. Biol. Med. 2010, 150, 83–85. [Google Scholar] [CrossRef] [PubMed]
- Gil, B.; Anastasova, S.; Yang, G.Z.A. Smart Wireless Ear-Worn Device for cardiovascular and sweat parameter monitoring during physical exercise: Design and performance results. Sensors 2019, 19, 1616. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurement Model | Baseline Lactate (mM) | Peak Lactate (mM) |
|---|---|---|
| Rat–IV infusion | 0.75–1.0 | 3.1–8.0 |
| Human-exercise | 1.6–5.2 | 6.0–12.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spehar-Délèze, A.-M.; Anastasova, S.; Vadgama, P. Monitoring of Lactate in Interstitial Fluid, Saliva and Sweat by Electrochemical Biosensor: The Uncertainties of Biological Interpretation. Chemosensors 2021, 9, 195. https://doi.org/10.3390/chemosensors9080195
Spehar-Délèze A-M, Anastasova S, Vadgama P. Monitoring of Lactate in Interstitial Fluid, Saliva and Sweat by Electrochemical Biosensor: The Uncertainties of Biological Interpretation. Chemosensors. 2021; 9(8):195. https://doi.org/10.3390/chemosensors9080195
Chicago/Turabian StyleSpehar-Délèze, Anna-Maria, Salzitsa Anastasova, and Pankaj Vadgama. 2021. "Monitoring of Lactate in Interstitial Fluid, Saliva and Sweat by Electrochemical Biosensor: The Uncertainties of Biological Interpretation" Chemosensors 9, no. 8: 195. https://doi.org/10.3390/chemosensors9080195
APA StyleSpehar-Délèze, A. -M., Anastasova, S., & Vadgama, P. (2021). Monitoring of Lactate in Interstitial Fluid, Saliva and Sweat by Electrochemical Biosensor: The Uncertainties of Biological Interpretation. Chemosensors, 9(8), 195. https://doi.org/10.3390/chemosensors9080195