
EGFR Amplification Is a Phenomenon of IDH Wildtype and TERT Mutated High-Grade Glioma: An Integrated Analysis Using Fluorescence In Situ Hybridization and DNA Methylome Profiling


Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Collection
2.2. Molecular Genetic Characterization of Gliomas
2.3. Fluorescence In Situ Hybridization (FISH) Analysis
2.4. Infinium Methylation EPIC Array Analysis
2.5. Computational Data Analysis
3. Results
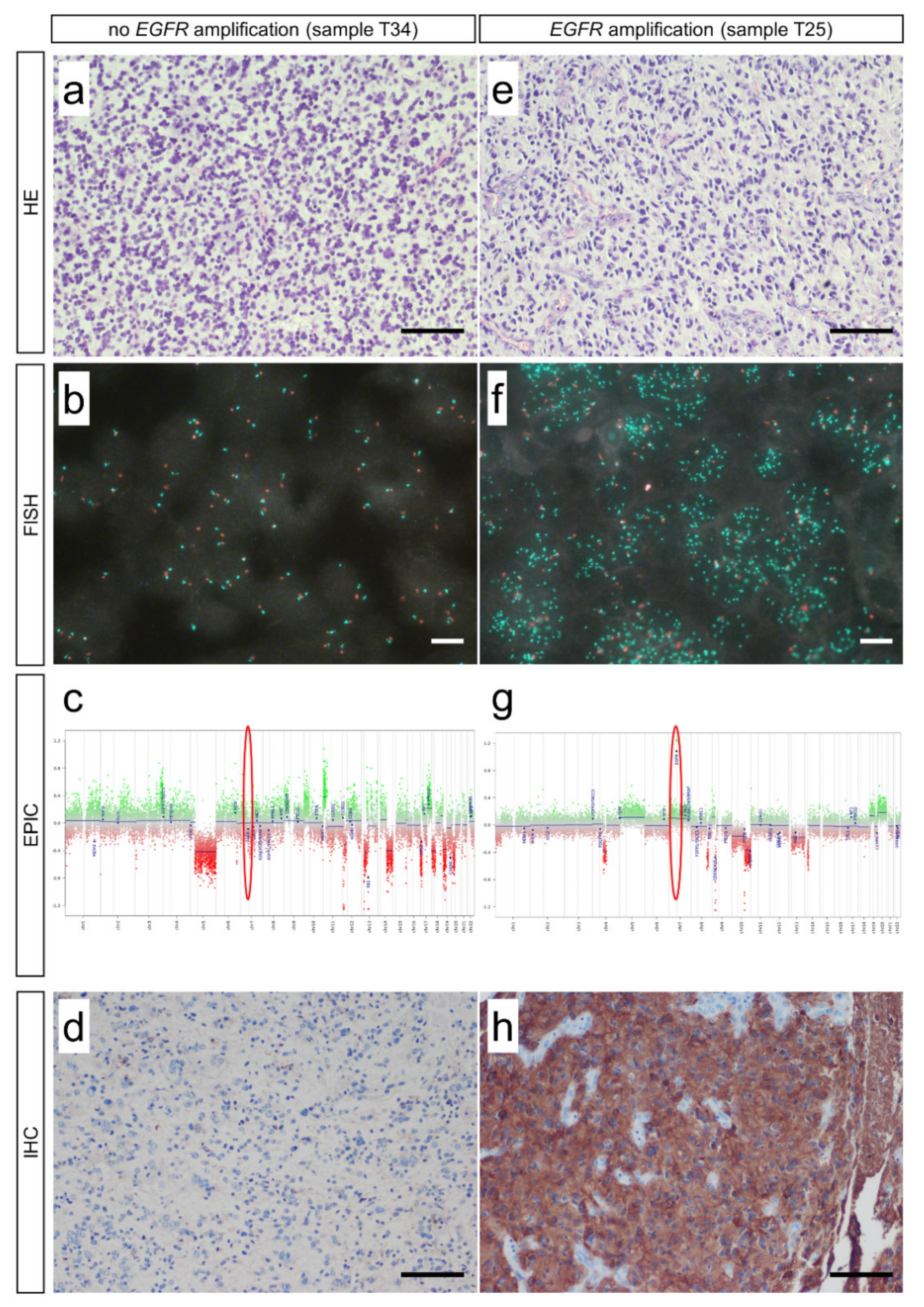
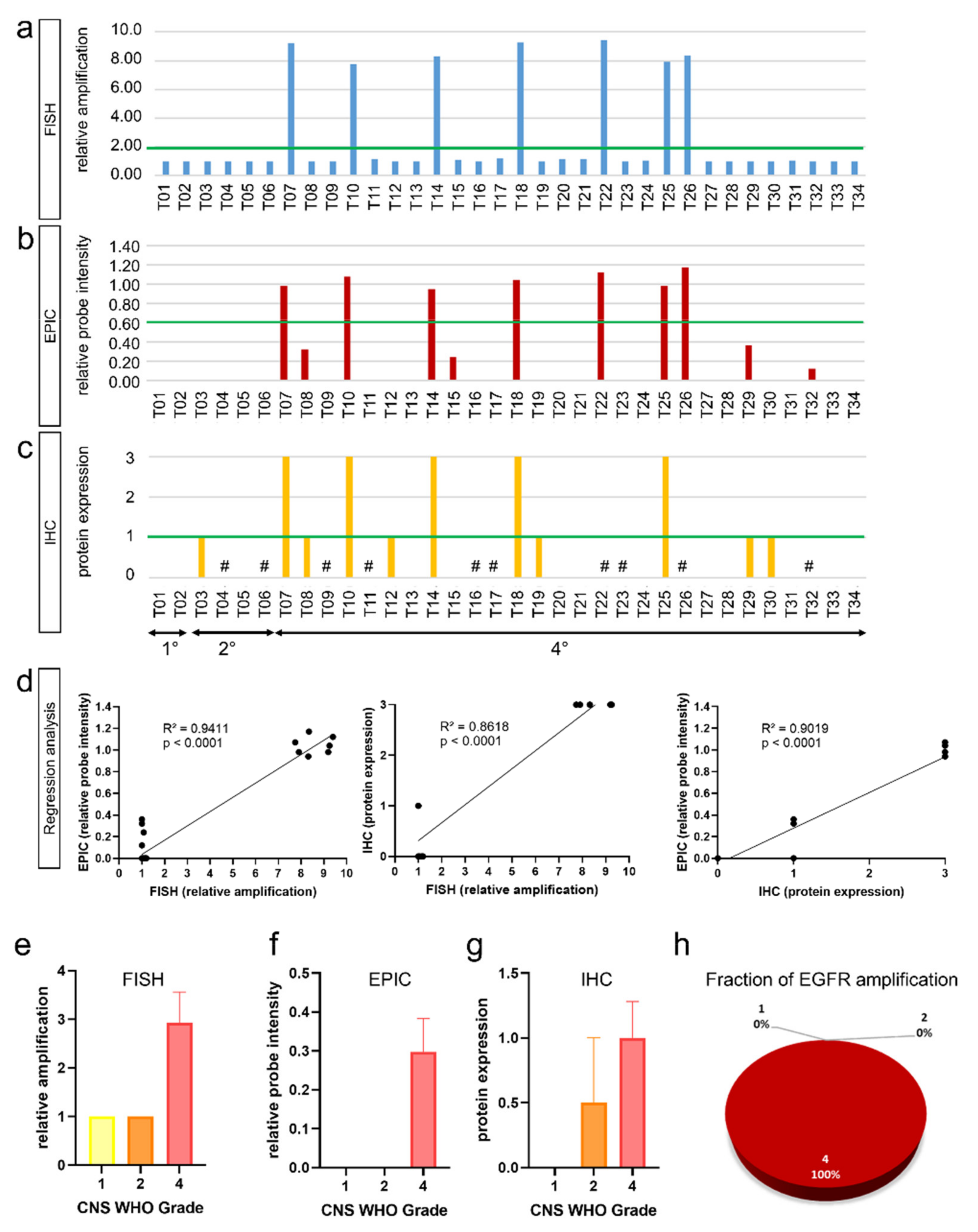
3.1. Fluorescence In Situ Hybridization (FISH) and Infinium EPIC Methylation Bead Chip Analysis Are Equally Valid Methods in Detection of EGFR Amplifications
3.2. EGFR Amplifications Predominantly Occur in High-Grade Glioma
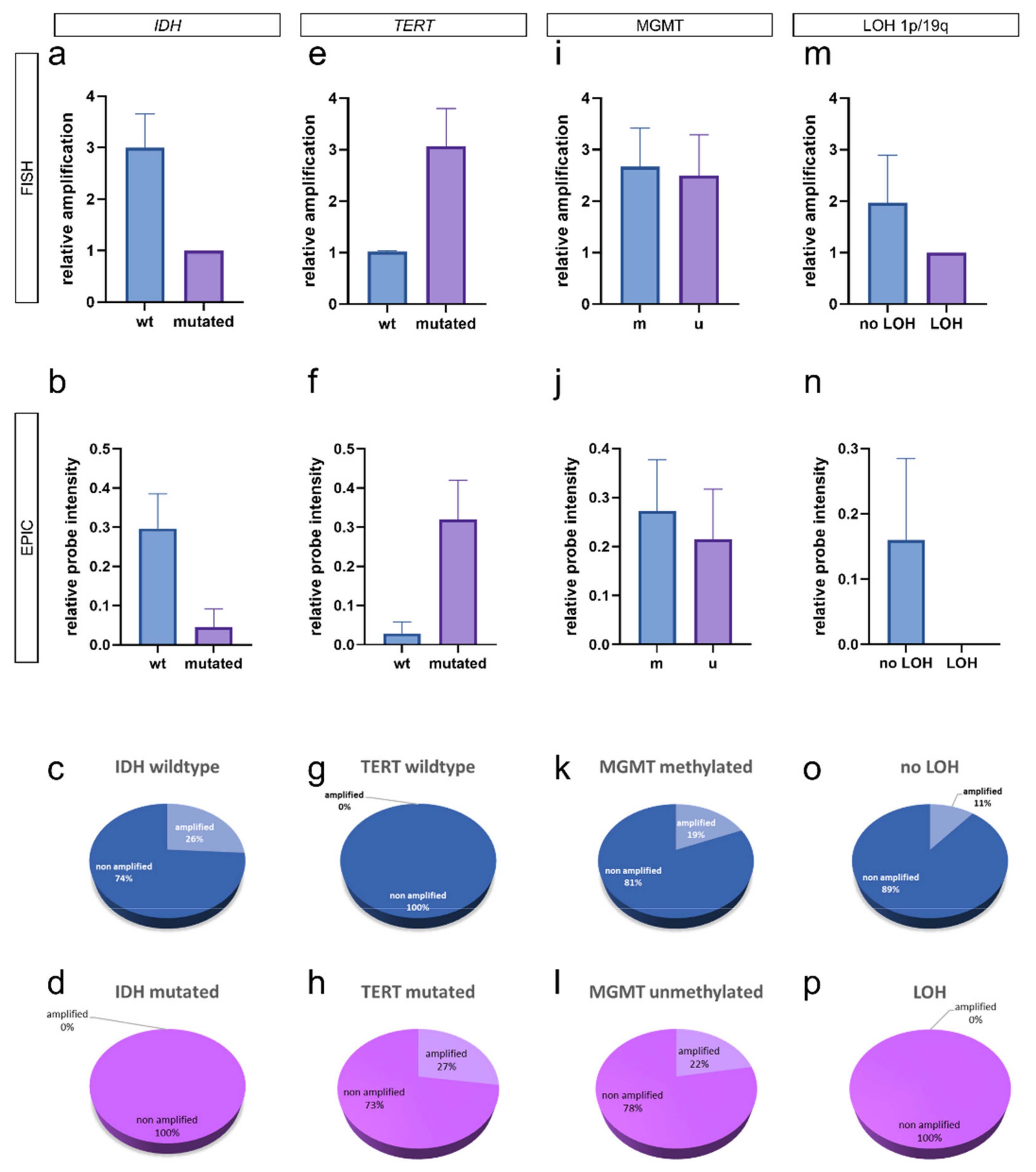
3.3. Integrated Analysis of EGFR Amplification and Molecular Glioma Hallmarks
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 who classification of tumors of the central nervous system: A summary. Neuro-Oncology 2021, 23, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Ameratunga, M.; Pavlakis, N.; Wheeler, H.; Grant, R.; Simes, J.; Khasraw, M. Anti-angiogenic therapy for high-grade glioma. Cochrane Database Syst. Rev. 2018, 11, Cd008218. [Google Scholar] [CrossRef] [PubMed]
- Gregory, T.A.; Chumbley, L.B.; Henson, J.W.; Theeler, B.J. Adult pilocytic astrocytoma in the molecular era: A comprehensive review. CNS Oncol. 2021, 10, CNS68. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, S.K.; Brothers, S.P.; Wahlestedt, C. Emerging treatment strategies for glioblastoma multiforme. EMBO Mol. Med. 2014, 6, 1359–1370. [Google Scholar] [CrossRef]
- Aldape, K.; Brindle, K.M.; Chesler, L.; Chopra, R.; Gajjar, A.; Gilbert, M.R.; Gottardo, N.; Gutmann, D.H.; Hargrave, D.; Holland, E.C.; et al. Challenges to curing primary brain tumours. Nat. Rev. Clin. Oncol. 2019, 16, 509–520. [Google Scholar] [CrossRef] [Green Version]
- Hegi, M.E.; Diserens, A.C.; Godard, S.; Dietrich, P.Y.; Regli, L.; Ostermann, S.; Otten, P.; Van Melle, G.; de Tribolet, N.; Stupp, R. Clinical trial substantiates the predictive value of o-6-methylguanine-DNA methyltransferase promoter methylation in glioblastoma patients treated with temozolomide. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2004, 10, 1871–1874. [Google Scholar] [CrossRef] [Green Version]
- Hegi, M.E.; Diserens, A.C.; Gorlia, T.; Hamou, M.F.; de Tribolet, N.; Weller, M.; Kros, J.M.; Hainfellner, J.A.; Mason, W.; Mariani, L.; et al. Mgmt gene silencing and benefit from temozolomide in glioblastoma. N. Engl. J. Med. 2005, 352, 997–1003. [Google Scholar] [CrossRef] [Green Version]
- Hegi, M.E.; Liu, L.; Herman, J.G.; Stupp, R.; Wick, W.; Weller, M.; Mehta, M.P.; Gilbert, M.R. Correlation of o6-methylguanine methyltransferase (mgmt) promoter methylation with clinical outcomes in glioblastoma and clinical strategies to modulate mgmt activity. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2008, 26, 4189–4199. [Google Scholar] [CrossRef] [Green Version]
- Hegi, M.E.; Janzer, R.C.; Lambiv, W.L.; Gorlia, T.; Kouwenhoven, M.C.; Hartmann, C.; von Deimling, A.; Martinet, D.; Besuchet Schmutz, N.; Diserens, A.C.; et al. Presence of an oligodendroglioma-like component in newly diagnosed glioblastoma identifies a pathogenetically heterogeneous subgroup and lacks prognostic value: Central pathology review of the eortc_26981/ncic_ce.3 trial. Acta Neuropathol. 2012, 123, 841–852. [Google Scholar] [CrossRef] [Green Version]
- Hau, P.; Stupp, R.; Hegi, M.E. Mgmt methylation status: The advent of stratified therapy in glioblastoma? Dis. Markers 2007, 23, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Rose, M.; Maurer, A.; Wirtz, J.; Bleilevens, A.; Waldmann, T.; Wenz, M.; Eyll, M.; Geelvink, M.; Gereitzig, M.; Rüchel, N.; et al. Egfr activity addiction facilitates anti-erbb based combination treatment of squamous bladder cancer. Oncogene 2020, 39, 6856–6870. [Google Scholar] [CrossRef]
- Slanina, H.; Mündlein, S.; Hebling, S.; Schubert-Unkmeir, A. Role of epidermal growth factor receptor signaling in the interaction of neisseria meningitidis with endothelial cells. Infect. Immun. 2014, 82, 1243–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Bent, M.J.; Gao, Y.; Kerkhof, M.; Kros, J.M.; Gorlia, T.; van Zwieten, K.; Prince, J.; van Duinen, S.; Sillevis Smitt, P.A.; Taphoorn, M.; et al. Changes in the egfr amplification and egfrviii expression between paired primary and recurrent glioblastomas. Neuro-Oncology 2015, 17, 935–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, F.; Harris, R.C. Epidermal growth factor, from gene organization to bedside. Semin. Cell Dev. Biol. 2014, 28, 2–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wee, P.; Wang, Z. Epidermal growth factor receptor cell proliferation signaling pathways. Cancers 2017, 9, 52. [Google Scholar] [CrossRef] [Green Version]
- Schaff, L.R.; Yan, D.; Thyparambil, S.; Tian, Y.; Cecchi, F.; Rosenblum, M.; Reiner, A.S.; Panageas, K.S.; Hembrough, T.; Lin, A.L. Characterization of mgmt and egfr protein expression in glioblastoma and association with survival. J. Neuro-Oncol. 2020, 146, 163–170. [Google Scholar] [CrossRef]
- Capper, D.; Jones, D.T.W.; Sill, M.; Hovestadt, V.; Schrimpf, D.; Sturm, D.; Koelsche, C.; Sahm, F.; Chavez, L.; Reuss, D.E.; et al. DNA methylation-based classification of central nervous system tumours. Nature 2018, 555, 469–474. [Google Scholar] [CrossRef]
- Capper, D.; Stichel, D.; Sahm, F.; Jones, D.T.W.; Schrimpf, D.; Sill, M.; Schmid, S.; Hovestadt, V.; Reuss, D.E.; Koelsche, C.; et al. Practical implementation of DNA methylation and copy-number-based cns tumor diagnostics: The heidelberg experience. Acta Neuropathol. 2018, 136, 181–210. [Google Scholar] [CrossRef] [Green Version]
- Assenov, Y.; Muller, F.; Lutsik, P.; Walter, J.; Lengauer, T.; Bock, C. Comprehensive analysis of DNA methylation data with rnbeads. Nat. Methods 2014, 11, 1138–1140. [Google Scholar] [CrossRef] [Green Version]
- Muller, F.; Scherer, M.; Assenov, Y.; Lutsik, P.; Walter, J.; Lengauer, T.; Bock, C. Rnbeads 2.0: Comprehensive analysis of DNA methylation data. Genome Biol. 2019, 20, 55. [Google Scholar] [CrossRef] [Green Version]
- Aviles-Salas, A.; Muniz-Hernandez, S.; Maldonado-Martinez, H.A.; Chanona-Vilchis, J.G.; Ramirez-Tirado, L.A.; HernaNdez-Pedro, N.; Dorantes-Heredia, R.; Rui, Z.M.J.M.; Motola-Kuba, D.; Arrieta, O. Reproducibility of the egfr immunohistochemistry scores for tumor samples from patients with advanced non-small cell lung cancer. Oncol. Lett. 2017, 13, 912–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraus, T.F.J.; Machegger, L.; Poppe, J.; Zellinger, B.; Dovjak, E.; Schlicker, H.U.; Schwartz, C.; Ladisich, B.; Spendel, M.; Kral, M.; et al. Diffuse midline glioma of the cervical spinal cord with h3 k27m genotype phenotypically mimicking anaplastic ganglioglioma: A case report and review of the literature. Brain Tumor Pathol. 2020, 37, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Holzl, D.; Hutarew, G.; Zellinger, B.; Schlicker, H.U.; Schwartz, C.; Winkler, P.A.; Sotlar, K.; Kraus, T.F.J. Integrated analysis of programmed cell death ligand 1 expression reveals increased levels in high-grade glioma. J. Cancer Res. Clin. Oncol. 2021, 147, 2271–2280. [Google Scholar] [CrossRef] [PubMed]
- Kraus, T.F.J.; Schwartz, C.; Machegger, L.; Zellinger, B.; Holzl, D.; Schlicker, H.U.; Poppe, J.; Ladisich, B.; Spendel, M.; Kral, M.; et al. A patient with two gliomas with independent oligodendroglioma and glioblastoma biology proved by DNA-methylation profiling: A case report and review of the literature. Brain Tumor Pathol. 2022. [Google Scholar] [CrossRef]
- French, P.J.; Eoli, M.; Sepulveda, J.M.; de Heer, I.; Kros, J.M.; Walenkamp, A.; Frenel, J.S.; Franceschi, E.; Clement, P.M.; Weller, M.; et al. Defining egfr amplification status for clinical trial inclusion. Neuro-Oncology 2019, 21, 1263–1272. [Google Scholar] [CrossRef] [Green Version]
- Stichel, D.; Ebrahimi, A.; Reuss, D.; Schrimpf, D.; Ono, T.; Shirahata, M.; Reifenberger, G.; Weller, M.; Hänggi, D.; Wick, W.; et al. Distribution of egfr amplification, combined chromosome 7 gain and chromosome 10 loss, and tert promoter mutation in brain tumors and their potential for the reclassification of idhwt astrocytoma to glioblastoma. Acta Neuropathol. 2018, 136, 793–803. [Google Scholar] [CrossRef] [Green Version]
- Wen, P.Y.; Weller, M.; Lee, E.Q.; Alexander, B.M.; Barnholtz-Sloan, J.S.; Barthel, F.P.; Batchelor, T.T.; Bindra, R.S.; Chang, S.M.; Chiocca, E.A.; et al. Glioblastoma in adults: A society for neuro-oncology (sno) and european society of neuro-oncology (eano) consensus review on current management and future directions. Neuro-Oncology 2020, 22, 1073–1113. [Google Scholar] [CrossRef]
- Hegi, M.E.; Sciuscio, D.; Murat, A.; Levivier, M.; Stupp, R. Epigenetic deregulation of DNA repair and its potential for therapy. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2009, 15, 5026–5031. [Google Scholar] [CrossRef] [Green Version]
- Kaina, B.; Christmann, M.; Naumann, S.; Roos, W.P. Mgmt: Key node in the battle against genotoxicity, carcinogenicity and apoptosis induced by alkylating agents. DNA Repair 2007, 6, 1079–1099. [Google Scholar] [CrossRef]
- Aquilanti, E.; Miller, J.; Santagata, S.; Cahill, D.P.; Brastianos, P.K. Updates in prognostic markers for gliomas. Neuro-Oncology 2018, 20, vii17–vii26. [Google Scholar] [CrossRef] [Green Version]
- De Witt Hamer, P.C. Small molecule kinase inhibitors in glioblastoma: A systematic review of clinical studies. Neuro-Oncology 2010, 12, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Bale, T.A.; Jordan, J.T.; Rapalino, O.; Ramamurthy, N.; Jessop, N.; DeWitt, J.C.; Nardi, V.; Alvarez, M.M.; Frosch, M.; Batchelor, T.T.; et al. Financially effective test algorithm to identify an aggressive, egfr-amplified variant of idh-wildtype, lower-grade diffuse glioma. Neuro-Oncology 2019, 21, 596–605. [Google Scholar] [CrossRef] [PubMed]
- Jaunmuktane, Z.; Capper, D.; Jones, D.T.W.; Schrimpf, D.; Sill, M.; Dutt, M.; Suraweera, N.; Pfister, S.M.; von Deimling, A.; Brandner, S. Methylation array profiling of adult brain tumours: Diagnostic outcomes in a large, single centre. Acta Neuropathol. Commun. 2019, 7, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| ID | Diagnosis | Grade | Age (y) | Sex | FISH | EPIC | IHC | IDH1 | IDH2 | 1p/19q | TERT | H3F3A | MGMT | CDKN2A/B |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T01 | Pilocytic astrocytoma | 1 | 38 | f | 1.00 | 0.00 | 0 | wt | wt | n.a. | wt | n.a. | u | n |
| T02 | Pilocytic astrocytoma | 1 | 16 | m | 1.00 | 0.00 | 0 | wt | wt | n.a. | wt | n.a. | u | n |
| T03 | Oligod. IDH mut. 1p/19q codel | 2 | 27 | m | 1.00 | 0.00 | 1 | R132H | wt | 1p/19q | C228T | n.a. | m | n |
| T04 | Oligod. IDH mut. 1p/19q codel. | 2 | 63 | f | 1.00 | 0.00 | n.a. | R132H | wt | 1p/19q | C250T | n.a. | m | n |
| T05 | Astrocytoma IDH mutant | 2 | 47 | m | 1.00 | 0.00 | 0 | R132S | wt | wt | wt | n.a. | u | n |
| T06 | Astrocytoma IDH mutant | 2 | 22 | m | 1.00 | 0.00 | n.a. | R132C | wt | wt | wt | n.a. | u | n |
| T07 | Glioblastoma IDH wildtype | 4 | 47 | m | 9.20 | 0.98 | 3 | wt | wt | n.a. | n.a. | n.a. | u | d |
| T08 | Astrocytoma IDH mutant | 4 | 37 | f | 1.00 | 0.32 | 1 | R132H | wt | wt | wt | n.a. | m | d |
| T09 | Glioblastoma IDH wildtype | 4 | 70 | f | 1.00 | 0.00 | n.a. | wt | wt | n.a. | C250T | n.a. | m | d |
| T10 | Glioblastoma IDH wildtype | 4 | 66 | m | 7.74 | 1.07 | 3 | wt | wt | n.a. | C228T | n.a. | m | d |
| T11 | Glioblastoma IDH wildtype | 4 | 62 | m | 1.13 | 0.00 | n.a. | wt | wt | n.a. | C250T | n.a. | m | n |
| T12 | Astrocytoma IDH mutant | 4 | 45 | f | 1.00 | 0.00 | 1 | R132H | wt | wt | wt | n.a. | m | n |
| T13 | Glioblastoma IDH wildtype | 4 | 77 | m | 1.00 | 0.00 | 0 | wt | wt | n.a. | C228T | n.a. | u | n |
| T14 | Glioblastoma IDH wildtype | 4 | 74 | m | 8.32 | 0.94 | 3 | wt | wt | n.a. | C250T | n.a. | m | n |
| T15 | Glioblastoma IDH wildtype | 4 | 43 | f | 1.08 | 0.24 | n.a. | wt | wt | n.a. | C250T | n.a. | u | n |
| T16 | Astrocytoma IDH mutant | 4 | 38 | m | 1.00 | 0.00 | n.a. | R132H | wt | wt | wt | n.a. | m | n |
| T17 | Glioblastoma IDH wildtype | 4 | 26 | m | 1.20 | 0.00 | 0 | wt | wt | n.a. | wt | n.a. | u | n |
| T18 | Glioblastoma IDH wildtype | 4 | 69 | m | 9.25 | 1.04 | 3 | wt | wt | n.a. | C228T | n.a. | m | d |
| T19 | Glioblastoma IDH wildtype | 4 | 63 | f | 1.00 | 0.00 | 1 | wt | wt | n.a. | C250T | n.a. | m | d |
| T20 | Glioblastoma IDH wildtype | 4 | 32 | m | 1.15 | 0.00 | 0 | wt | wt | 1p | C228T | n.a. | u | n |
| T21 | Glioblastoma IDH wildtype | 4 | 72 | m | 1.15 | 0.00 | 0 | wt | wt | wt | C228T | n.a. | m | d |
| T22 | Glioblastoma IDH wildtype | 4 | 79 | f | 9.4 | 1.12 | n.a. | wt | wt | wt | C228T | n.a. | u | d |
| T23 | Glioblastoma IDH wildtype | 4 | 75 | f | 1.00 | 0.00 | n.a. | wt | wt | 1p | C250T | n.a. | u | d |
| T24 | Glioblastoma IDH wildtype | 4 | 45 | f | 1.05 | 0.00 | 0 | wt | wt | n.a. | C228T | wt | m | n |
| T25 | Glioblastoma IDH wildtype | 4 | 77 | f | 7.90 | 0.98 | 3 | wt | wt | n.a. | C228T | n.a. | u | d |
| T26 | Glioblastoma IDH wildtype | 4 | 49 | m | 8.35 | 1.17 | n.a. | wt | wt | n.a. | C250T | n.a. | m | d |
| T27 | Glioblastoma IDH wildtype | 4 | 25 | m | 1.00 | 0.00 | 0 | wt | wt | n.a. | wt | wt | u | n |
| T28 | Glioblastoma IDH wildtype | 4 | 51 | m | 1.00 | 0.00 | 0 | wt | wt | n.a. | C250T | n.a. | m | d |
| T29 | Glioblastoma IDH wildtype | 4 | 63 | f | 1.00 | 0.36 | 1 | wt | wt | n.a. | C250T | n.a. | m | d |
| T30 | Glioblastoma IDH wildtype | 4 | 65 | f | 1.00 | 0.00 | 1 | wt | wt | n.a. | C228T | n.a. | m | d |
| T31 | Glioblastoma IDH wildtype | 4 | 76 | f | 1.05 | 0.00 | 0 | wt | wt | n.a. | wt | n.a. | m | n |
| T32 | Glioblastoma IDH wildtype | 4 | 72 | f | 1.00 | 0.12 | n.a. | wt | wt | n.a. | C250T | n.a. | u | d |
| T33 | Dif. midline glioma H3 K27 alt. | 4 | 38 | m | 1.00 | 0.00 | 0 | wt | wt | n.a. | C228T | K27M | u | n |
| T34 | Dif. midline glioma H3 K27 alt. | 4 | 33 | f | 1.00 | 0.00 | 0 | wt | wt | n.a. | wt | K27M | u | n |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hölzl, D.; Hutarew, G.; Zellinger, B.; Alinger-Scharinger, B.; Schlicker, H.U.; Schwartz, C.; Sotlar, K.; Kraus, T.F.J. EGFR Amplification Is a Phenomenon of IDH Wildtype and TERT Mutated High-Grade Glioma: An Integrated Analysis Using Fluorescence In Situ Hybridization and DNA Methylome Profiling. Biomedicines 2022, 10, 794. https://doi.org/10.3390/biomedicines10040794
Hölzl D, Hutarew G, Zellinger B, Alinger-Scharinger B, Schlicker HU, Schwartz C, Sotlar K, Kraus TFJ. EGFR Amplification Is a Phenomenon of IDH Wildtype and TERT Mutated High-Grade Glioma: An Integrated Analysis Using Fluorescence In Situ Hybridization and DNA Methylome Profiling. Biomedicines. 2022; 10(4):794. https://doi.org/10.3390/biomedicines10040794
Chicago/Turabian StyleHölzl, Dorothee, Georg Hutarew, Barbara Zellinger, Beate Alinger-Scharinger, Hans U. Schlicker, Christoph Schwartz, Karl Sotlar, and Theo F. J. Kraus. 2022. "EGFR Amplification Is a Phenomenon of IDH Wildtype and TERT Mutated High-Grade Glioma: An Integrated Analysis Using Fluorescence In Situ Hybridization and DNA Methylome Profiling" Biomedicines 10, no. 4: 794. https://doi.org/10.3390/biomedicines10040794
APA StyleHölzl, D., Hutarew, G., Zellinger, B., Alinger-Scharinger, B., Schlicker, H. U., Schwartz, C., Sotlar, K., & Kraus, T. F. J. (2022). EGFR Amplification Is a Phenomenon of IDH Wildtype and TERT Mutated High-Grade Glioma: An Integrated Analysis Using Fluorescence In Situ Hybridization and DNA Methylome Profiling. Biomedicines, 10(4), 794. https://doi.org/10.3390/biomedicines10040794