
Mechanisms Underlying Antipsychotic-Induced NAFLD and Iron Dysregulation: A Multi-Omic Approach



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Proteomic and Immunomic Changes Induced by Risperidone and Olanzapine in Non-Obese Mice
2.2. Transcriptomic Changes Induced by Risperidone Treatment of Human Neuroblastoma Cells In Vitro
2.3. Transcriptomic Changes Induced by Onset of Aripiprazole Treatment in Psychiatric Patients
2.4. Compilation of Multi-Omic DE Traits
2.5. Pathway Analysis to Detect Alterations in Traits Associated with NAFLD
2.6. Pathway Analysis to Detect Alterations in Traits Associated with Iron Homeostasis
2.7. Measurement of Iron Accumulation in Livers of OLAN-Treated Mice
3. Results
3.1. Parameters of Subjects Assessed for Differential Expression Induced by Atypical Antipsychotic Drugs
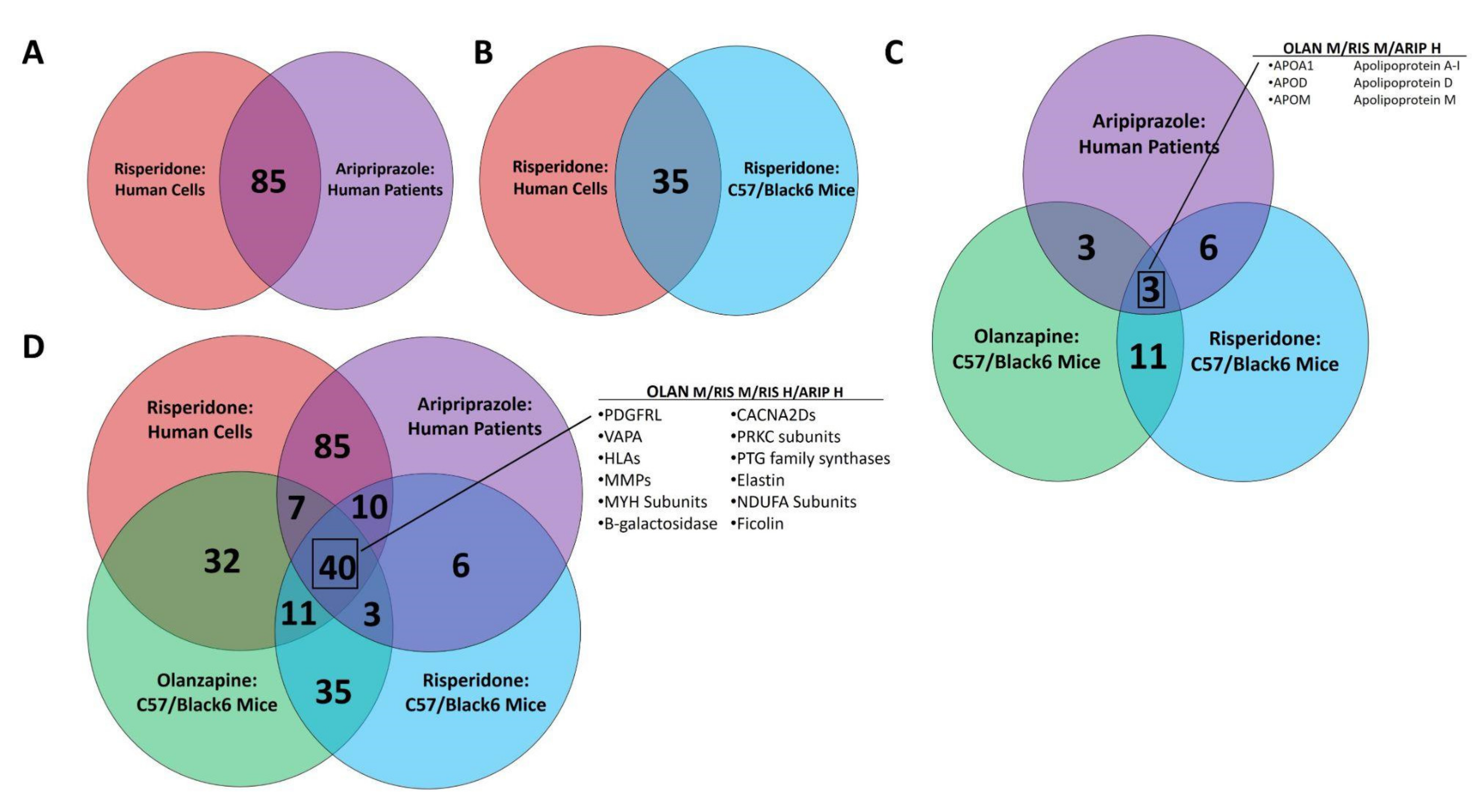
3.2. Multi-omic Analysis Predicts Impacts on NAFLD Development Following Atypical Antipsychotic Drug Exposure In Vitro and In Vivo
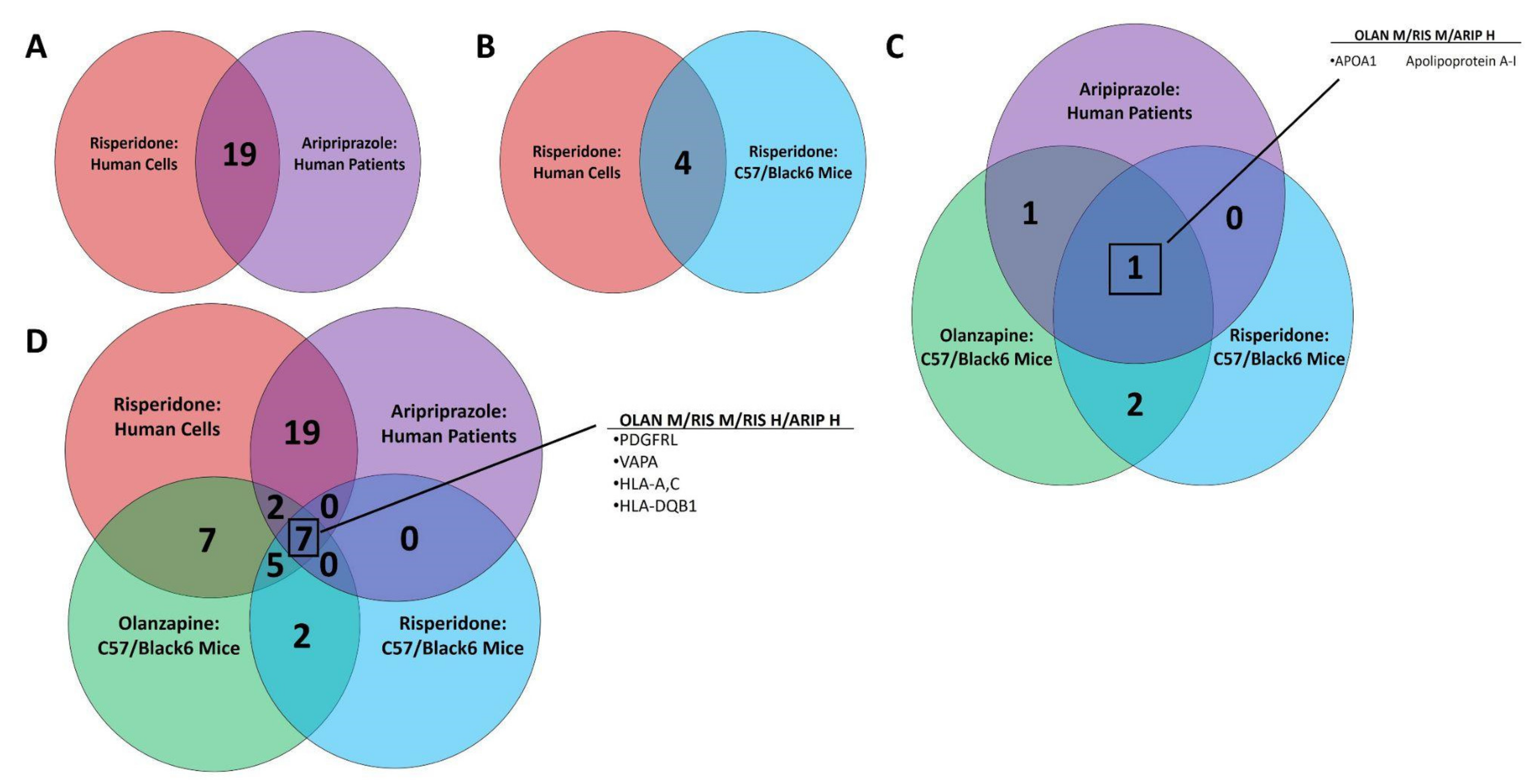
3.3. Multi-omic Analysis Predicts Impacts on Iron Homeostasis following Atypical Antipsychotic Drug Exposure In Vitro and In Vivo
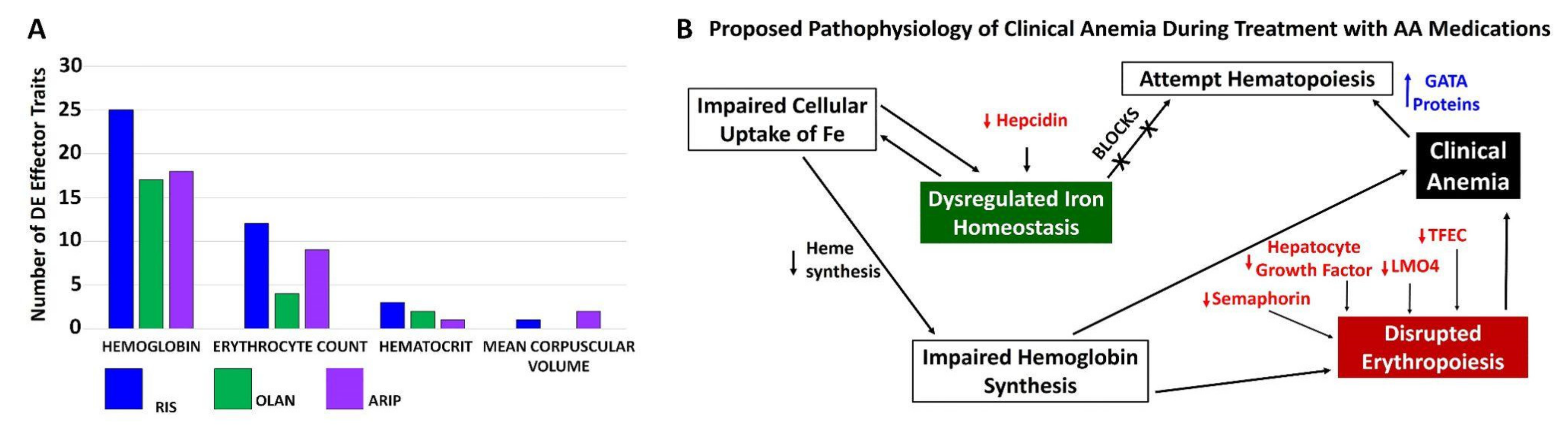
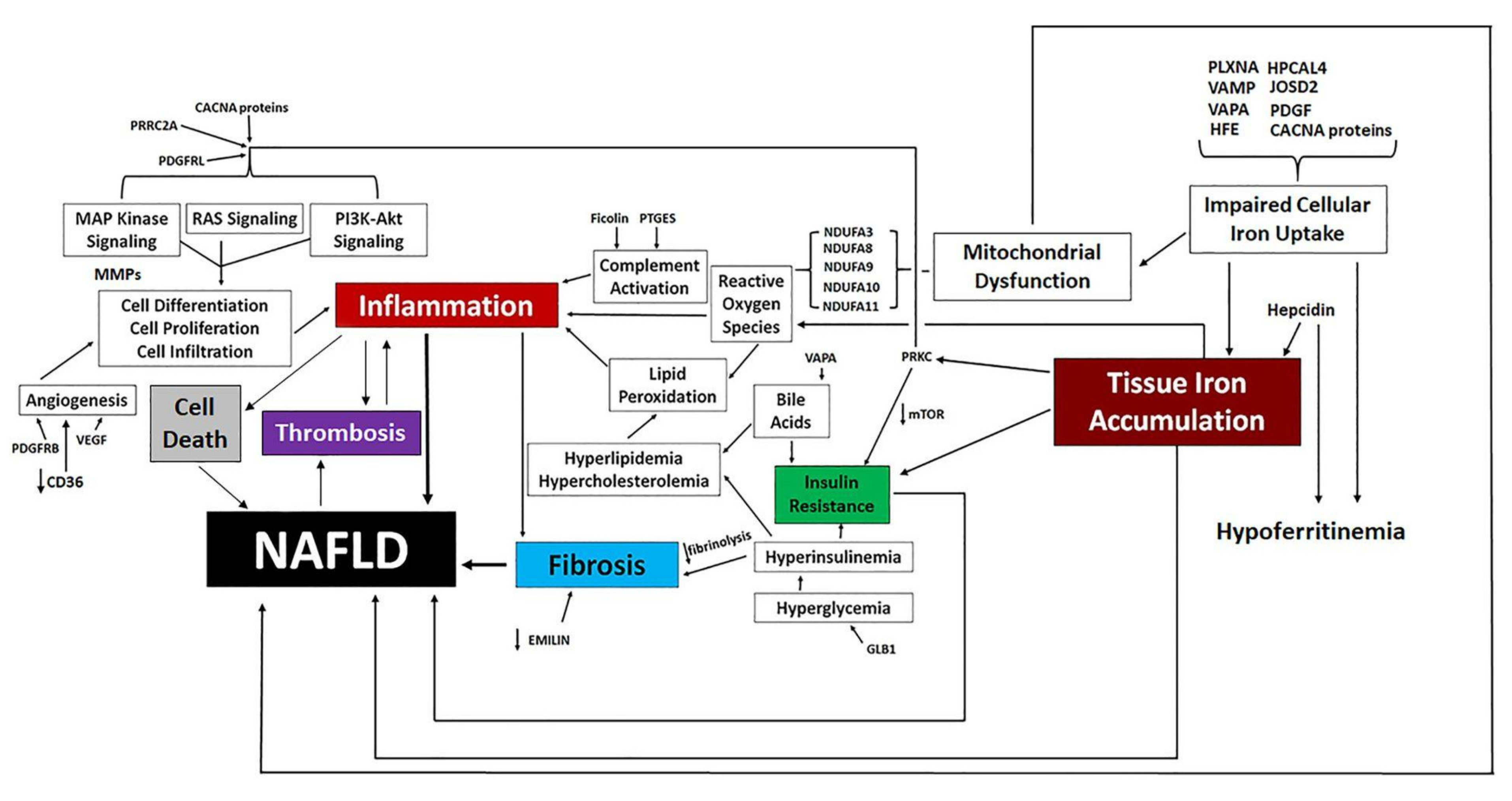
3.4. Proposed Mechanisms of Altered Iron Homeostasis Following Pathway Analysis of Traits Differentially Expressed by Atypical Antipsychotic Drugs
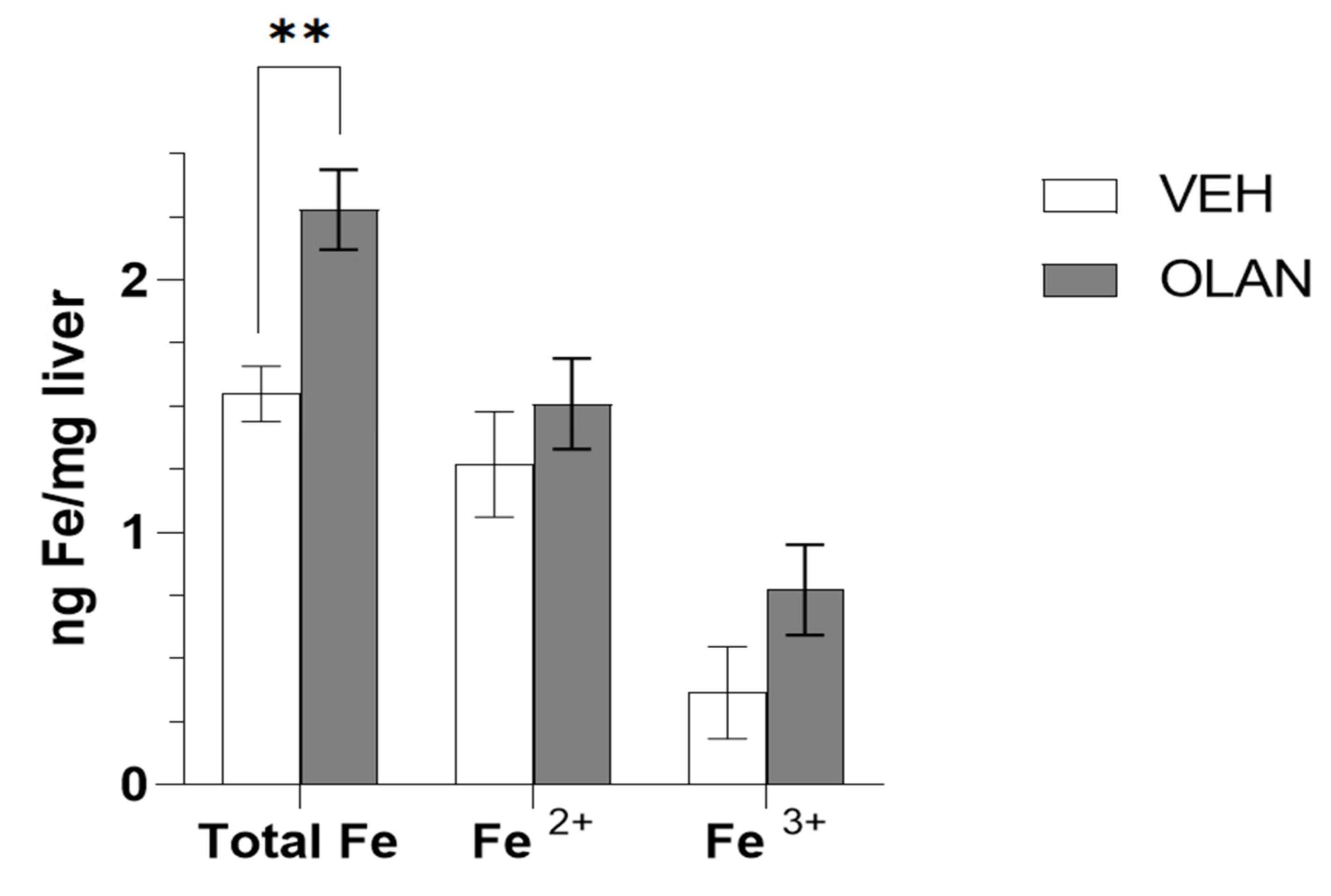
3.5. Measurement of Accumulated Iron in Mice Treated with OLAN
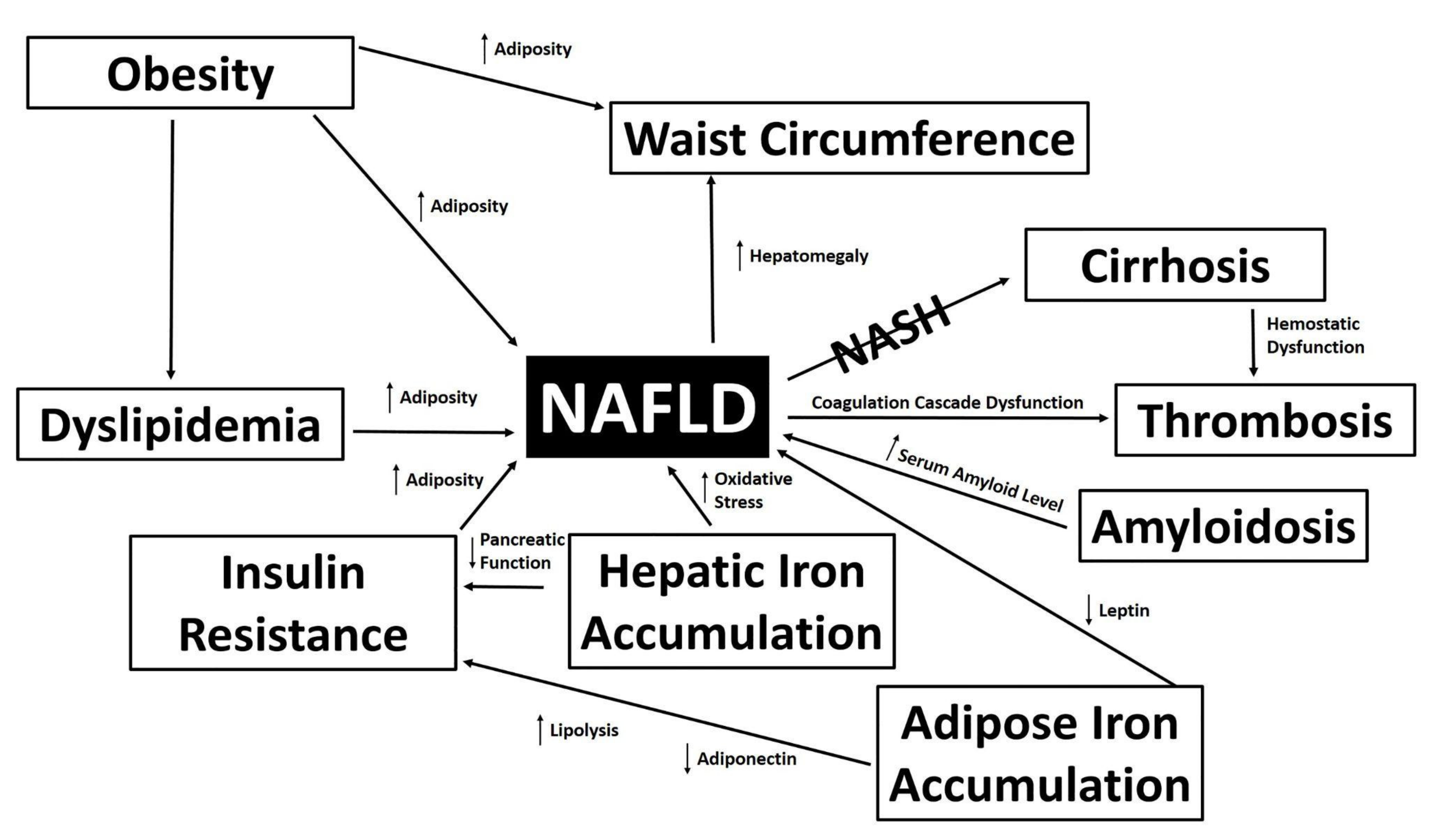
3.6. Proposed Mechanisms of NAFLD Development Following Pathway Analysis of Traits Differentially Expressed by Atypical Antipsychotic Drugs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Germani, G.; Becchetti, C. Liver transplantation for non-alcoholic fatty liver disease. Minerva Gastroenterol. Dietol. 2018, 64, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Benedict, M.; Zhang, X. Non-Alcoholic Fatty Liver Disease: An Expanded Review. World J. Hepatol. 2017, 9, 715–732. [Google Scholar] [CrossRef] [PubMed]
- Stefan, N.; Cusi, K. A global view of the interplay between non-alcoholic fatty liver disease and diabetes. Lancet Diabetes Endocrinol. 2022, 10, 284–296. [Google Scholar] [CrossRef]
- Anstee, Q.M.; Targher, G.; Day, C.P. Progression of NAFLD to diabetes mellitus, cardiovascular disease or cirrhosis. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 330–344. [Google Scholar] [CrossRef] [PubMed]
- Colognesi, M.; Gabbia, D.; De Martin, S. Depression and Cognitive Impairment—Extrahepatic Manifestations of NAFLD and NASH. Biomedicines 2020, 8, 229. [Google Scholar] [CrossRef]
- Soto-Angona, Ó.; Anmella, G.; Valdés-Florido, M.J.; De Uribe-Viloria, N.; Carvalho, A.F.; Penninx, B.W.J.H.; Berk, M. Non-alcoholic fatty liver disease (NAFLD) as a neglected metabolic companion of psychiatric disorders: Common pathways and future approaches. BMC Med. 2020, 18, 261. [Google Scholar] [CrossRef]
- Rostama, B.; Beauchemin, M.; Bouchard, C.; Bernier, E.; Vary, C.P.; May, M.; Houseknecht, K.L. Understanding Mechanisms Underlying Non-Alcoholic Fatty Liver Disease (NAFLD) in Mental Illness: Risperidone and Olanzapine Alter the Hepatic Proteomic Signature in Mice. Int. J. Mol. Sci. 2020, 21, 9362. [Google Scholar] [CrossRef]
- Xu, H.; Zhuang, X. Atypical antipsychotics-induced metabolic syndrome and nonalcoholic fatty liver disease: A critical review. Neuropsychiatr. Dis. Treat. 2019, 15, 2087–2099. [Google Scholar] [CrossRef] [Green Version]
- May, M.; Beauchemin, M.; Vary, C.; Barlow, D.; Houseknecht, K.L. The antipsychotic medication, risperidone, causes global immunosuppression in healthy mice. PLoS ONE 2019, 14, e0218937. [Google Scholar] [CrossRef]
- Freyberg, Z.; Aslanoglou, D.; Shah, R.; Ballon, J.S. Intrinsic and Antipsychotic Drug-Induced Metabolic Dysfunction in Schizophrenia. Front. Neurosci. 2017, 11, 432. [Google Scholar] [CrossRef]
- Bauer, M.E.; Teixeira, A.L. Inflammation in psychiatric disorders: What comes first? Ann. N. Y. Acad. Sci. USA 2019, 1437, 57–67. [Google Scholar] [CrossRef]
- Alvarez-Jiménez, M.; González-Blanch, C.; Crespo-Facorro, B.; Hetrick, S.; Rodríguez-Sánchez, J.M.; Pérez-Iglesias, R.; Vázquez-Barquero, J.L. Antipsychotic-Induced Weight Gain in Chronic and First-Episode Psychotic Disorders: A Systematic Critical Reappraisal. CNS Drugs 2008, 22, 547–562. [Google Scholar] [CrossRef]
- Pillinger, T.; McCutcheon, R.A.; Vano, L.; Mizuno, Y.; Arumuham, A.; Hindley, G.; Beck, K.; Natesan, S.; Efthimiou, O.; Cipriani, A.; et al. Comparative Effects of 18 Antipsychotics on Metabolic Function in Patients with Schizophrenia, Predictors of Metabolic Dysregulation, and Association with Psychopathology: A Systematic Review and Network Meta-Analysis. Lancet Psychiatry 2020, 7, 64–77. [Google Scholar] [CrossRef]
- Zipursky, R.B.; Gu, H.; Green, A.I.; Perkins, D.O.; Tohen, M.F.; McEvoy, J.P.; Strakowski, S.M.; Sharma, T.; Kahn, R.S.; Gur, R.E.; et al. Course and Predictors of Weight Gain in People with First-Episode Psychosis Treated with Olanzapine or Haloperidol. Br. J. Psychiatry 2005, 187, 537–543. [Google Scholar] [CrossRef] [Green Version]
- Annamalai, A.; Kosir, U.; Tek, C. Prevalence of Obesity and Diabetes in Patients with Schizophrenia. World J. Diabetes 2017, 15, 390–396. [Google Scholar] [CrossRef]
- Beauchemin, M.; Geguchadze, R.; Guntur, A.R.; Nevola, K.; Le, P.T.; Barlow, D.; Rue, M.; Vary, C.P.H.; Lary, C.W.; Motyl, K.J.; et al. Exploring mechanisms of increased cardiovascular disease risk with antipsychotic medications: Risperidone alters the cardiac proteomic signature in mice. Pharmacol. Res. 2020, 152, 104589. [Google Scholar] [CrossRef]
- Houseknecht, K.L.; Robertson, A.S.; Zavadoski, W.; Gibbs, E.M.; Johnson, D.E.; Rollema, H. Acute effects of atypical antipsychotics on whole-body insulin resistance in rats: Implications for adverse metabolic effects. Neuropsychopharmacology 2007, 32, 289–297. [Google Scholar] [CrossRef] [Green Version]
- Dongiovanni, P.; Fracanzani, A.L.; Fargion, S.; Valenti, L. Iron in fatty liver and in the metabolic syndrome: A promising therapeutic target. J. Hepatol. 2011, 55, 920–932. [Google Scholar] [CrossRef]
- Marmur, J.; Beshara, S.; Eggertsen, G.; Onelöv, L.; Albiin, N.; Danielsson, O.; Hultcrantz, R.; Stål, P. Hepcidin levels correlate to liver iron content, but not steatohepatitis, in non-alcoholic fatty liver disease. BMC Gastroenterol. 2018, 18, 78. [Google Scholar] [CrossRef]
- McCann, J.C.; Ames, B.N. An overview of evidence for a causal relation between iron deficiency during development and deficits in cognitive or behavioral function. Am. J. Clin. Nutr. 2007, 85, 931–945. [Google Scholar] [CrossRef] [Green Version]
- Calarge, C.A.; Murry, D.J.; Ziegler, E.E.; Arnold, L.E. Serum Ferritin, Weight Gain, Disruptive Behavior, and Extrapyramidal Symptoms in Risperidone-Treated Youth. J. Child Adolesc. Psychopharmacol. 2016, 26, 471–477. [Google Scholar] [CrossRef] [Green Version]
- Grantham-McGregor, S.; Ani, C. A review of studies on the effect of iron deficiency on cognitive development in children. J. Nutr. 2001, 131, 649S–666S, discussion 666S–668S. [Google Scholar] [CrossRef]
- Nelson, J.E.; Klintworth, H.; Kowdley, K.V. Iron metabolism in Nonalcoholic Fatty Liver Disease. Curr. Gastroenterol. Rep. 2012, 14, 8–16. [Google Scholar] [CrossRef]
- Schoretsanitis, G.; Nikolakopoulou, A.; Guinart, D.; Correll, C.U.; Kane, J.M. Iron homeostasis alterations and risk for akathisia in patients treated with antipsychotics: A systematic review and meta-analysis of cross-sectional studies. Eur. Neuropsychopharmacol. 2020, 35, 1–11. [Google Scholar] [CrossRef]
- Chong, S.A.; Mythily, M.B.B.S.; Remington, G. Tardive dyskinesia and iron status. J. Clin. Psychopharmacol. 2004, 24, 235–236. [Google Scholar] [CrossRef]
- Calarge, C.A.; Ziegler, E.E.; Del Castillo, N.; Aman, M.; McDougle, C.J.; Scahill, L.; McCracken, J.T.; Arnold, L.E. Iron homeostasis during risperidone treatment in children and adolescents. J. Clin. Psychiatry 2015, 76, 1500–1505. [Google Scholar] [CrossRef]
- Malekizadeh, Y.; Williams, G.; Kelson, M.; Whitfield, D.; Mill, J.; Collier, D.A.; Ballard, C.; Jeffries, A.R.; Creese, B. Whole transcriptome in silico screening implicates cardiovascular and infectious disease in the mechanism of action underlying atypical antipsychotic side effects. Alzheimers Dement. 2020, 6, e12078. [Google Scholar] [CrossRef]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets—Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [Green Version]
- Crespo-Facorro, B.; Ortiz-Garcia de la Foz, V.; Suarez-Pinilla, P.; Valdizan, E.M.; Pérez-Iglesias, R.; Amado-Señaris, J.A.; Teresa Garcia-Unzueta, M.; Labad, J.; Correll, C.; Ayesa-Arriola, R. Effects of aripiprazole, quetiapine and ziprasidone on plasma prolactin levels in individuals with first episode nonaffective psychosis: Analysis of a randomized open-label 1 year study. Schizophr. Res. 2017, 189, 134–141. [Google Scholar] [CrossRef]
- Crespo-Facorro, B.; Ruiz-Veguilla, M.; Vázquez-Bourgon, J.; Sánchez-Hidalgo, A.C.; Garrido-Torres, N.; Cisneros, J.M.; Prieto, C.; Sainz, J. Aripiprazole as a Candidate Treatment of COVID-19 Identified Through Genomic Analysis. Front. Pharmacol. 2021, 12, 646701. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.G.; Barnes, K.C.; Bright, T.J.; Wang, S.A. The genetic association database. Nat. Genet. 2004, 36, 431–432. [Google Scholar] [CrossRef] [Green Version]
- Soliman, H.M.; Wagih, H.M.; Algaidi, S.A.; Hafiz, A.H. Histological evaluation of the role of atypical antipsychotic drugs in inducing non-alcoholic fatty liver disease in adult male albino rats (light and electron microscopic study). Folia Biol. 2013, 59, 173–180. [Google Scholar]
- Morlán-Coarasa, M.J.; Arias-Loste, M.T.; Ortiz-García de la Foz, V.; Martínez-García, O.; Alonso-Martín, C.; Crespo, J.; Romero-Gómez, M.; Fábrega, E.; Crespo-Facorro, B. Incidence of non-alcoholic fatty liver disease and metabolic dysfunction in first episode schizophrenia and related psychotic disorders: A 3-year prospective randomized interventional study. Psychopharmacology 2016, 233, 3947–3952. [Google Scholar] [CrossRef]
- Koreki, A.; Mori, H.; Nozaki, S.; Koizumi, T.; Suzuki, H.; Onaya, M. Risk of Nonalcoholic Fatty Liver Disease in Patients With Schizophrenia Treated With Antipsychotic Drugs: A Cross-sectional Study. J. Clin. Psychopharmacol. 2021, 41, 474–477. [Google Scholar] [CrossRef]
- Li, R.; Zhu, W.; Huang, P.; Yang, Y.; Luo, F.; Dai, W.; Shen, L.; Pei, W.; Huang, X. Olanzapine leads to nonalcoholic fatty liver disease through the apolipoprotein A5 pathway. Biomed. Pharmacother. 2021, 141, 111803. [Google Scholar] [CrossRef]
- Chen, X.; Li, Y.; Zhang, T.; Yao, Y.; Shen, C.; Xue, Y. Association of Serum Trace Elements with Schizophrenia and Effects of Antipsychotic Treatment. Biol. Trace Elem. Res. 2018, 181, 22–30. [Google Scholar] [CrossRef]
- Calarge, C.A.; Ziegler, E.E. Iron deficiency in pediatric patients in long-term risperidone treatment. J. Child Adolesc. Psychopharmacol. 2013, 23, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Eleftheriou, G.; Butera, R.; Barcella, L.; Falanga, A. Clozapine-induced anemia: A case-report. Int. J. Clin. Pharmacol. Ther. 2020, 58, 289–292. [Google Scholar] [CrossRef]
- Kirpekar, V.C.; Faye, A.D.; Bhave, S.H.; Tadke, R.; Gawande, S. Lurasidone-induced anemia: Is there a need for hematological monitoring? Indian J. Pharmacol. 2019, 51, 276–278. [Google Scholar]
- Capeto Coelho, I.; Aleixo, A.; Linhares, L.; Martins, A.; Telles Correia, D. Hemolytic anemia potentially associated with aripiprazole. Psychiatry Clin. Neurosci. 2018, 72, 876. [Google Scholar] [CrossRef] [Green Version]
- Moulin, V.; Grandoni, F.; Castioni, J.; Lu, H. Pancytopenia in an adult patient with thiamine-responsive megaloblastic anaemia. BMJ Case Rep. 2018, 2018, bcr-2018. [Google Scholar] [CrossRef]
- Cusi, K.; Isaacs, S.; Barb, D.; Rinella, M.E.; Vos, M.B.; Younossi, Z. American Association of Clinical Endocrinology Clinical Practice Guideline for the Diagnosis and Management of Nonalcoholic Fatty Liver Disease in Primary Care and Endocrinology Clinical Settings Co-Sponsored by the American Association for the Study of Liver Diseases (AASLD). AACE Endocr. Pract. 2022, 28, 528–562. [Google Scholar]
- Milic, S.; Mikolasevic, I.; Orlic, L.; Devcic, E.; Starcevic-Cizmarevic, N.; Stimac, D.; Kapovic, M.; Ristic, S. The Role of Iron and Iron Overload in Chronic Liver Disease. Med. Sci. Monit. 2016, 22, 2144–2151. [Google Scholar] [CrossRef] [Green Version]
- Datz, C.; Müller, E.; Aigner, E. Iron overload and non-alcoholic fatty liver disease. Minerva Endocrinol. 2017, 42, 173–183. [Google Scholar] [CrossRef]
- Masarone, M.; Rosato, V.; Dallio, M.; Gravina, A.G.; Aglitti, A.; Loguercio, C.; Federico, A.; Persico, M. Role of Oxidative Stress in Pathophysiology of Nonalcoholic Fatty Liver Disease. Oxid. Med. Cell. Longev. 2018, 2018, 9547613. [Google Scholar] [CrossRef]
- Zhong, F.; Guan, L.; Lin, H.; Zhao, M.; Qin, Y.; Li, Q.; Yuan, Z.; Yang, G.; Gao, L.; Zhao, J. Red Blood Cell Count: An Unrecognized Risk Factor for Nonalcoholic Fatty Liver Disease. Front. Endocrinol. 2021, 12, 760981. [Google Scholar] [CrossRef]
- Jiang, Y.; Zeng, J.; Chen, B. Hemoglobin combined with triglyceride and ferritin in predicting non-alcoholic fatty liver. J. Gastroenterol. Hepatol. 2014, 29, 1508–1514. [Google Scholar] [CrossRef]
- Oudit, G.Y.; Trivieri, M.G.; Khaper, N.; Liu, P.P.; Backx, P.H. Role of L-type Ca2+ channels in iron transport and iron-overload cardiomyopathy. J. Mol. Med. 2006, 84, 349–364. [Google Scholar] [CrossRef]
- Del Campo, A.; Bustos, C.; Mascayano, C.; Acuña-Castillo, C.; Troncoso, R.; Rojo, L.E. Metabolic Syndrome and Antipsychotics: The Role of Mitochondrial Fission/Fusion Imbalance. Front. Endocrinol. 2018, 9, 144. [Google Scholar] [CrossRef] [Green Version]
- Bouvier, M.L.; Fehsel, K.; Schmitt, A.; Meisenzahl-Lechner, E.; Gaebel, W.; Von Wilmsdorff, M. Sex-dependent effects of long-term clozapine or haloperidol medication on red blood cells and liver iron metabolism in Sprague Dawley rats as a model of metabolic syndrome. BMC Pharmacol. Toxicol. 2022, 23, 8. [Google Scholar] [CrossRef]
- American Diabetes Association; American Psychiatric Association; American Association of Clinical Endocrinologists; North American Association for the Study of Obesity. Consensus Development Conference on Antipsychotic Drugs and Obesity and Diabetes. J. Clin. Psychiatry 2004, 65, 267–272. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
May, M.; Barlow, D.; Ibrahim, R.; Houseknecht, K.L. Mechanisms Underlying Antipsychotic-Induced NAFLD and Iron Dysregulation: A Multi-Omic Approach. Biomedicines 2022, 10, 1225. https://doi.org/10.3390/biomedicines10061225
May M, Barlow D, Ibrahim R, Houseknecht KL. Mechanisms Underlying Antipsychotic-Induced NAFLD and Iron Dysregulation: A Multi-Omic Approach. Biomedicines. 2022; 10(6):1225. https://doi.org/10.3390/biomedicines10061225
Chicago/Turabian StyleMay, Meghan, Deborah Barlow, Radwa Ibrahim, and Karen L. Houseknecht. 2022. "Mechanisms Underlying Antipsychotic-Induced NAFLD and Iron Dysregulation: A Multi-Omic Approach" Biomedicines 10, no. 6: 1225. https://doi.org/10.3390/biomedicines10061225
APA StyleMay, M., Barlow, D., Ibrahim, R., & Houseknecht, K. L. (2022). Mechanisms Underlying Antipsychotic-Induced NAFLD and Iron Dysregulation: A Multi-Omic Approach. Biomedicines, 10(6), 1225. https://doi.org/10.3390/biomedicines10061225