
Rifaximin Improves Spatial Learning and Memory Impairment in Rats with Liver Damage-Associated Neuroinflammation

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Model and Treatment with Rifaximin
2.2. Immunohistochemistry
2.3. Analysis of IL-1β, TNFα and CCL2 Expression in the CA1-Region of Hippocampus
2.4. Analysis of Astrocytes and Microglia Activation
2.5. Membrane Surface Expression of Subunits of AMPA and NMDA Glutamate Receptors
2.6. Novel Object Recognition (NOR) and Novel Object Location (NOL) Memory Tests
2.7. Radial Maze
2.8. Statistical Analysis
3. Results
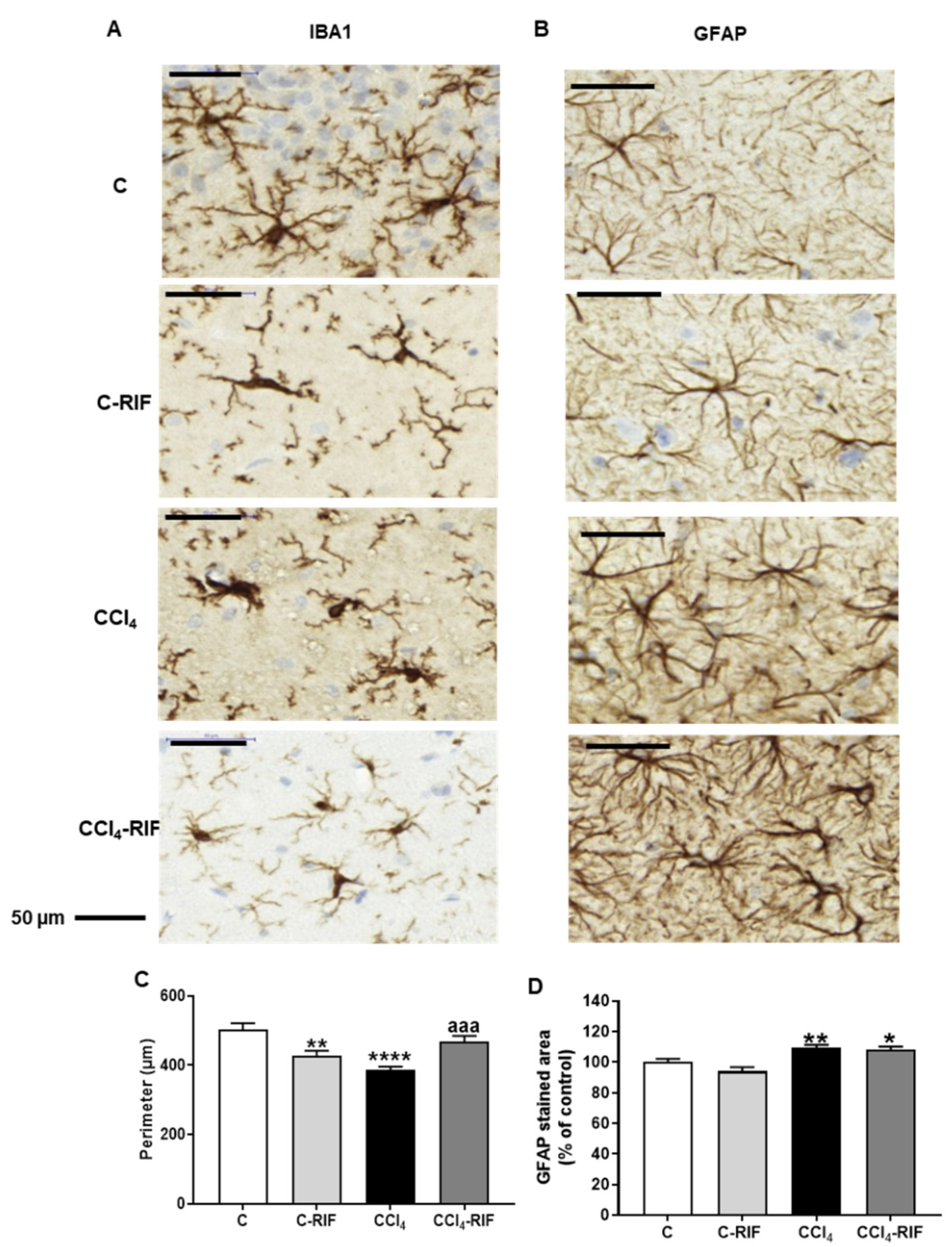
3.1. Rats with Mild Liver Damage Show Microglia and Astrocyte Activation in the Hippocampus Rifaximin Reverses Microglia but Not Astrocyte Activation
3.2. Hippocampus of Rats with Mild Liver Damage Shows an Increase in Infiltrated Macrophages and Lumphocytes
3.3. Membrane Expression of NMDA and AMPA Receptor Subunits Is Altered in the Hippocampus of Rats with Mild Liver Damage
3.4. Spatial Learning and Memory Are Impaired in Rats with Mild Liver Damage While Non-Spatial Memory and Working Memory Are Not Altered
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Felipo, V. Hepatic encephalopathy: Effects of liver failure on brain function. Nat. Rev. Neurosci. 2013, 14, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Felipo, V.; Urios, A.; Montesinos, E.; Molina, I.; Garcia-Torres, M.L.; Civera, M.; Del Olmo, J.A.; Ortega, J.; Martinez-Valls, J.; Serra, M.A.; et al. Contribution of hyperammonemia and inflammatory factors to cognitive impairment in minimal hepatic encephalopathy. Metab. Brain Dis. 2012, 27, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.W.; Gottesman, R.F.; Clark, J.M.; Hernaez, R.; Chang, Y.; Kim, C.; Ha, K.H.; Guallar, E.; Lazo, M. Nonalcoholic fatty liver disease is associated with cognitive function in adults. Neutology 2016, 86, 1136–1142. [Google Scholar] [CrossRef] [Green Version]
- Giménez-Garzó, C.; Fiorillo, A.; Ballester-Ferré, M.P.; Gallego, J.J.; Casanova-Ferrer, F.; Urios, A.; Benlloch, S.; Martí-Aguado, D.; San-Miguel, T.; Tosca, J.; et al. A new score unveils a high prevalence of mild cognitive impairment in patients with nonalcoholic fatty liver disease. J. Clin. Med. 2021, 10, 2806. [Google Scholar] [CrossRef] [PubMed]
- Cauli, O.; Rodrigo, R.; Piedrafita, B.; Boix, J.; Felipo, V. Inflammation and hepatic encephalopathy: Ibuprofen restores learning ability in rats with portacaval shunts. Hepatology 2007, 46, 514–519. [Google Scholar] [CrossRef]
- Rodrigo, R.; Cauli, O.; Gomez-Pinedo, U.; Agusti, A.; Hernandez-Rabaza, V.; Garcia-Verdugo, J.M.; Felipo, V. Hyperammonemia induces neuroinflammation that contributes to cognitive impairment in rats with hepatic encephalopathy. Gastroenterology 2010, 139, 675–684. [Google Scholar] [CrossRef]
- Agusti, A.; Cauli, O.; Rodrigo, R.; Llansola, M.; Hernández-Rabaza, V.; Felipo, V. p38 MAP kinase is a therapeutic target for hepatic encephalopathy in rats with portacaval shunts. Gut 2011, 60, 1572–1579. [Google Scholar] [CrossRef]
- Mumby, D.G.; Gaskin, S.; Glenn, M.J.; Schramek, T.E.; Lehmann, H. Hippocampal damage and exploratory preferences in rats: Memory for objects, places, and contexts. Learn. Mem. 2002, 9, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Barker, G.R.I.; Warburton, E.C. When is the hippocampus involved in recognition memory? J. Neurosci. 2011, 31, 10721–10731. [Google Scholar] [CrossRef] [Green Version]
- Broadbent, N.J.; Squire, L.R.; Clark, R.E. Spatial memory, recognition memory, and the hippocampus. Proc. Natl. Acad. Sci. USA 2004, 101, 14515–14520. [Google Scholar] [CrossRef] [Green Version]
- Hammond, R.S.; Tull, L.E.; Stackman, R.W. On the delay-dependent involvement of the hippocampus in object recognition memory. Neurobiol. Learn. Mem. 2004, 82, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Hyman, J.M.; Zilli, E.A.; Paley, A.M.; Hasselmo, M.E. Working memory performance correlates with prefrontal-hippocampal theta interactions but not with prefrontal neuron firing rates. Front. Integr. Neurosci. 2010, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, Y.; Mikami, A.; Fujii, Y.; Kamei, C. Effect of histamine on muscimol-induced working memory deficits in radial maze performance. J. Pharmacol. Sci. 2007, 104, 252–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, K.; Shimizu, M.; Kawabe, K.; Ichitani, Y. Hippocampal AP5 treatment impairs both spatial working and reference memory in radial maze performance in rats. Eur. J. Pharmacol. 2015, 758, 137–141. [Google Scholar] [CrossRef]
- Hernandez-Rabaza, V.; Agusti, A.; Cabrera-Pastor, A.; Fustero, S.; Delgado, O.; Taoro-Gonzalez, L.; Montoliu, C.; Llansola, M.; Felipo, V. Sildenafil reduces neuroinflammation and restores spatial learning in rats with hepatic encephalopathy: Underlying mechanisms. J. Neuroinflamm. 2015, 12, 195. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Rabaza, V.; Cabrera-Pastor, A.; Taoro-González, L.; Malaguarnera, M.; Agustí, A.; Llansola, M.; Felipo, V. Hyperammonemia induces glial activation, neuroinflammation and alters neurotransmitter receptors in hippocampus, impairing spatial learning: Reversal by sulforaphane. J. Neuroinflamm. 2016, 13, 41. [Google Scholar] [CrossRef] [Green Version]
- Dadsetan, S.; Balzano, T.; Forteza, J.; Cabrera-Pastor, A.; Taoro-Gonzalez, L.; Hernandez-Rabaza, V.; Gil-Perotín, S.; Cubas-Núñez, L.; García-Verdugo, J.M.; Agusti, A.; et al. Reducing peripheral inflammation with infliximab reduces neuroinflammation and improves cognition in rats with hepatic encephalopathy. Front. Mol. Neurosci. 2016, 9, 106. [Google Scholar] [CrossRef]
- Cabrera-Pastor, A.; Hernandez-Rabaza, V.; Taoro-Gonzalez, L.; Balzano, T.; Llansola, M.; Felipo, V. In vivo administration of extracellular cGMP normalizes TNF-α and membrane expression of AMPA receptors in hippocampus and spatial reference memory but not IL-1β, NMDA receptors in membrane and working memory in hyperammonemic rats. Brain Behav. Immun. 2016, 57, 360–370. [Google Scholar] [CrossRef]
- Taoro-González, L.; Cabrera-Pastor, A.; Sancho-Alonso, M.; Arenas, Y.M.; Meseguer-Estornell, F.; Balzano, T.; ElMlili, N.; Felipo, V. Differential role of interleukin-1β in neuroinflammation-induced impairment of spatial and nonspatial memory in hyperammonemic rats. FASEB J. 2019, 33, 9913–9928. [Google Scholar] [CrossRef]
- Balzano, T.; Dadsetan, S.; Forteza, J.; Cabrera-Pastor, A.; Taoro-Gonzalez, L.; Malaguarnera, M.; Gil-Perotin, S.; Cubas-Nuñez, L.; Casanova, B.; Castro-Quintas, A.; et al. Chronic hyperammonemia induces peripheral inflammation that leads to cognitive impairment in rats: Reversed by anti-TNF-α treatment. J. Hepatol. 2020, 73, 582–592. [Google Scholar] [CrossRef]
- Balzano, T.; Forteza, J.; Molina, P.; Giner, J.; Monzó, A.; Sancho-Jiménez, J.; Urios, A.; Montoliu, C.; Felipo, V. The Cerebellum of Patients with Steatohepatitis Shows Lymphocyte Infiltration, Microglial Activation and Loss of Purkinje and Granular Neurons. Sci. Rep. 2018, 8, 3004. [Google Scholar] [CrossRef] [PubMed]
- Zemtsova, I.; Görg, B.; Keitel, V.; Bidmon, H.J.; Schrör, K.; Häussinger, D. Microglia activation in hepatic encephalopathy in rats and humans. Hepatology 2011, 54, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Mangas-Losada, A.; García-García, R.; Urios, A.; Escudero-García, D.; Tosca, J.; Giner-Durán, R.; Serra, M.A.; Montoliu, C.; Felipo, V. Minimal hepatic encephalopathy is associated with expansion and activation of CD4+CD28−, Th22 and Tfh and B lymphocytes. Sci. Rep. 2017, 7, 1–13. [Google Scholar]
- Dadsetan, S.; Balzano, T.; Forteza, J.; Agusti, A.; Cabrera-Pastor, A.; Taoro-Gonzalez, L.; Hernandez-Rabaza, V.; Gomez-Gimenez, B.; ElMlili, N.; Llansola, M.; et al. Infliximab reduces peripheral inflammation, neuroinflammation, and extracellular GABA in the cerebellum and improves learning and motor coordination in rats with hepatic encephalopathy. J. Neuroinflamm. 2016, 13, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Carroll, S.J.; Kho, D.T.; Wiltshire, R.; Nelson, V.; Rotimi, O.; Johnson, R.; Angel, C.E.; Graham, E.S. Pro-inflammatory TNFaα and IL-1β differentially regulate the inflammatory phenotype of brain microvascular endothelial cells. J. Neuroinflamm. 2015, 12, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rummel, C.; Sachot, C.; Poole, S.; Luheshi, G.N. Circulating interleukin-6 induces fever through a STAT3-linked activation of COX-2 in the brain. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2006, 291, 1316–1326. [Google Scholar] [CrossRef]
- Krasnow, S.M.; Knoll, J.G.; Verghese, S.C.; Levasseur, P.R.; Marks, D.L. Amplification and propagation of interleukin-1β signaling by murine brain endothelial and glial cells. J. Neuroinflamm. 2017, 14, 133. [Google Scholar] [CrossRef]
- Izquierdo-Altarejos, P.; Cabrera-Pastor, A.; Gonzalez-King, H.; Montoliu, C.; Felipo, V. Extracellular Vesicles from Hyperammonemic Rats Induce Neuroinflammation and Motor Incoordination in Control Rats. Cells 2020, 9, 572. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Liao, S.; Wei, C.; Jia, D.; Wood, K.; Liu, Q.; Wang, X.; Shi, F.D.; Jin, W.N. Infiltration and persistence of lymphocytes during late-stage cerebral ischemia in middle cerebral artery occlusion and photothrombotic stroke models. J. Neuroinflamm. 2017, 14, 248. [Google Scholar] [CrossRef] [Green Version]
- Beuker, C.; Strecker, J.K.; Rawal, R.; Schmidt-Pogoda, A.; Ruck, T.; Wiendl, H.; Klotz, L.; Schäbitz, W.R.; Sommer, C.J.; Minnerup, H.; et al. Immune Cell Infiltration into the Brain after Ischemic Stroke in Humans Compared to Mice and Rats: A Systematic Review and Meta-Analysis. Transl. Stroke Res. 2021, 12, 976–990. [Google Scholar] [CrossRef]
- Arima, Y.; Kamimura, D.; Sabharwal, L.; Yamada, M.; Bando, H.; Ogura, H.; Atsumi, T.; Murakami, M. Regulation of immune cell infiltration into the CNS by regional neural inputs explained by the gate theory. Mediat. Inflamm. 2013, 2013, 898165. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, G.G.; Pacheco-Moisés, F.P.; Bitzer-Quintero, O.K.; Ramírez-Anguiano, A.C.; Flores-Alvarado, L.J.; Ramírez-Ramírez, V.; Macias-Islas, M.A.; Torres-Sánchez, E.D. Immunology and oxidative stress in multiple sclerosis: Clinical and basic approach. Clin. Dev. Immunol. 2013, 2013, 708659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belarbi, K.; Jopson, T.; Arellano, C.; Fike, J.R.; Rosi, S. CCR2 deficiency prevents neuronal dysfunction and cognitive impairments induced by cranial irradiation. Cancer Res. 2013, 73, 1201–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varvel, N.H.; Neher, J.J.; Bosch, A.; Wang, W.; Ransohoff, R.M.; Miller, R.J.; Dingledine, R. Infiltrating monocytes promote brain inflammation and exacerbate neuronal damage after status epilepticus. Proc. Natl. Acad. Sci. USA 2016, 113, E5665–E5674. [Google Scholar] [CrossRef] [Green Version]
- Chompre, G.; Cruz, E.; Maldonado, L.; Rivera-Amill, V.; Porter, J.T.; Noel, R.J. Astrocytic expression of HIV-1 Nef impairs spatial and recognition memory. Neurobiol. Dis. 2013, 49, 128–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balzano, T.; Leone, P.; Ivaylova, G.; Castro, M.C.; Reyes, L.; Ramón, C.; Malaguarnera, M.; Llansola, M.; Felipo, V. Rifaximin prevents T-lymphocytes and macrophages infiltration in cerebellum and restores motor incoordination in rats with mild liver damage. Biomedicines 2021, 9, 1002. [Google Scholar] [CrossRef]
- Mangas-Losada, A.; García-García, R.; Leone, P.; Ballester, M.P.; Cabrera-Pastor, A.; Urios, A.; Gallego, J.J.; Martínez-Pretel, J.J.; Giménez-Garzó, C.; Revert, F.; et al. Selective improvement by rifaximin of changes in the immunophenotype in patients who improve minimal hepatic encephalopathy. J. Transl. Med. 2019, 17, 293. [Google Scholar] [CrossRef] [Green Version]
- Bajaj, J.S. Review article: Potential mechanisms of action of rifaximin in the management of hepatic encephalopathy and other complications of cirrhosis. Aliment. Pharmacol. Ther. 2016, 43 (Suppl. S1), 11–26. [Google Scholar] [CrossRef] [Green Version]
- Bajaj, J.S.; Kakiyama, G.; Savidge, T.; Takei, H.; Kassam, Z.A.; Fagan, A.; Gavis, E.A.; Pandak, W.M.; Nittono, H.; Hylemon, P.B.; et al. Antibiotic-Associated Disruption of Microbiota Composition and Function in Cirrhosis Is Restored by Fecal Transplant. Hepatology 2018, 68, 1549–1558. [Google Scholar] [CrossRef]
- Kalambokis, G.N.; Mouzaki, A.; Rodi, M.; Tsianos, E.V. Rifaximin for the prevention of spontaneous bacterial peritonitis. World J. Gastroenterol. 2012, 18, 1700–1702. [Google Scholar] [CrossRef]
- Lee, G.-P.; Jeong, W.-I.; Jeong, D.-H.; Do, S.-H.; Kim, T.-H.; Jeong, K.-S. Diagnostic evaluation of carbon tetrachloride-induced rat hepatic cirrhosis model. Anticancer Res. 2005, 25, 1029–1038. [Google Scholar] [PubMed]
- Boudreau, A.C.; Milovanovic, M.; Conrad, K.L.; Nelson, C.; Ferrario, C.R.; Wolf, M.E. A protein cross-linking assay for measuring cell surface expression of glutamate receptor subunits in the rodent brain after in vivo treatments. Curr. Protoc. Neurosci. 2012, 59, 5–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Mello, C.; Le, T.; Swain, M.G. Cerebral microglia recruit monocytes into the brain in response to tumor necrosis factora signaling during peripheral organ inflammation. J. Neurosci. 2009, 29, 2089–2102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ní Chasaide, C.; Lynch, M.A. The role of the immune system in driving neuroinflammation. Brain Neurosci. Adv. 2020, 4, 239821281990108. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, H.; Minogue, A.M.; Rooney, S.; Lynch, M.A. Infiltrating macrophages contribute to age-related neuroinflammation in C57/BL6 mice. Mech. Ageing Dev. 2018, 173, 84–91. [Google Scholar] [CrossRef]
- Zhang, L.; Tan, J.; Jiang, X.; Qian, W.; Yang, T.; Sun, X.; Chen, Z.; Zhu, Q. Neuron-derived CCL2 contributes to microglia activation and neurological decline in hepatic encephalopathy. Biol. Res. 2017, 50, 26. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Dong, H.; Qian, Q.; Zhang, X.; Wang, Y.; Jin, W.; Qian, Y. Astrocyte-derived CCL2 participates in surgery-induced cognitive dysfunction and neuroinflammation via evoking microglia activation. Behav. Brain Res. 2017, 332, 145–153. [Google Scholar] [CrossRef]
- Zhang, K.; Wang, H.; Xu, M.; Frank, J.A.; Luo, J. Role of MCP-1 and CCR2 in ethanol-induced neuroinflammation and neurodegeneration in the developing brain. J. Neuroinflamm. 2018, 15, 197. [Google Scholar] [CrossRef]
- Hernandez-Rabaza, V.; Cabrera-Pastor, A.; Taoro-Gonzalez, L.; Gonzalez-Usano, A.; Agusti, A.; Balzano, T.; Llansola, M.; Felipo, V. Neuroinflammation increases GABAergic tone and impairs cognitive and motor function in hyperammonemia by increasing GAT-3 membrane expression. Reversal by sulforaphane by promoting M2 polarization of microglia. J. Neuroinflamm. 2016, 13, 83. [Google Scholar] [CrossRef] [Green Version]
- Balzano, T.; Arenas, Y.M.; Dadsetan, S.; Forteza, J.; Gil-Perotin, S.; Cubas-Nuñez, L.; Casanova, B.; Gracià, F.; Varela-Andrés, N.; Montoliu, C.; et al. Sustained hyperammonemia induces TNF-A in Purkinje neurons by activating the TNFR1-NF-κB pathway. J. Neuroinflamm. 2020, 17, 70. [Google Scholar] [CrossRef] [Green Version]
- Izadpanah, K.; Freyer, D.; Weber, J.R.; Braun, J.S. Brain parenchymal TNF-α and IL-1β induction in experimental pneumococcal meningitis. J. Neuroimmunol. 2014, 276, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Bai, L.; Zhou, W.; Yang, Y.; Zhang, J.; Li, L.; Jiang, M.; Mi, Y.; Li, T.T.; Zhang, X.; et al. PARP-1-regulated TNF-α expression in the dorsal root ganglia and spinal dorsal horn contributes to the pathogenesis of neuropathic pain in rats. Brain Behav. Immun. 2020, 88, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Villarroya, H.; Marie, Y.; Ouallet, J.C.; Le Saux, F.; Tchélingérian, J.L.; Baumann, N. Expression of TNFα in central neurons of lewis rat spinal cord after EAE induction. J. Neurosci. Res. 1997, 49, 592–599. [Google Scholar] [CrossRef]
- Chen, B.; Deng, X.; Wang, B.; Liu, H. Etanercept, an inhibitor of TNF-a, prevents propofol-induced neurotoxicity in the developing brain. Int. J. Dev. Neurosci. 2016, 55, 91–100. [Google Scholar] [CrossRef]
- Balosso, S.; Ravizza, T.; Pierucci, M.; Calcagno, E.; Invernizzi, R.; Di Giovanni, G.; Esposito, E.; Vezzani, A. Molecular and functional interactions between tumor necrosis factor-alpha receptors and the glutamatergic system in the mouse hippocampus: Implications for seizure susceptibility. Neuroscience 2009, 161, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Stellwagen, D.; Beattie, E.C.; Seo, J.Y.; Malenka, R.C. Differential regulation of AMPA receptor and GABA receptor trafficking by tumor necrosis factor-α. J. Neurosci. 2005, 25, 3219–3228. [Google Scholar] [CrossRef] [Green Version]
- Lai, A.Y.; Swayze, R.D.; El-Husseini, A.; Song, C. Interleukin-1 beta modulates AMPA receptor expression and phosphorylation in hippocampal neurons. J. Neuroimmunol. 2006, 175, 97–106. [Google Scholar] [CrossRef]
- Wheeler, D.; Knapp, E.; Bandaru, V.V.; Wang, Y.; Knorr, D.; Poirier, C.; Mattson, M.P.; Geiger, J.D.; Haughey, N.J. Tumor necrosis factor-α-induced neutral sphingomyelinase-2 modulates synaptic plasticity by controlling the membrane insertion of NMDA receptors. J. Neurochem. 2009, 109, 1237–1249. [Google Scholar] [CrossRef] [Green Version]
- He, P.; Liu, Q.; Wu, J.; Shen, Y. Genetic deletion of TNF receptor suppresses excitatory synaptic transmission via reducing AMPA receptor synaptic localization in cortical neurons. FASEB J. 2012, 26, 334–345. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, A.R.; Christensen, R.N.; Gensel, J.C.; Miller, B.A.; Sun, F.; Beattie, E.C.; Bresnahan, J.C.; Beattie, M.S. Cell death after spinal cord injury is exacerbated by rapid TNFα-induced trafficking of GluR2-lacking AMPARs to the plasma membrane. J. Neurosci. 2008, 28, 11391–11400. [Google Scholar]
- Ogoshi, F.; Yin, H.Z.; Kuppumbatti, Y.; Song, B.; Amindari, S.; Weiss, J.H. Tumor necrosis-factor-alpha (TNF-α) induces rapid insertion of Ca2+-permeable α-amino-3-hydroxyl-5-methyl-4-isoxazole-propionate (AMPA)/kainate (Ca-A/K) channels in a subset of hippocampal pyramidal neurons. Exp. Neurol. 2005, 193, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Agusti, A.; Hernández-Rabaza, V.; Balzano, T.; Taoro-Gonzalez, L.; Ibañez-Grau, A.; Cabrera-Pastor, A.; Fustero, S.; Llansola, M.; Montoliu, C.; Felipo, V. Sildenafil reduces neuroinflammation in cerebellum, restores GABAergic tone, and improves motor in-coordination in rats with hepatic encephalopathy. CNS Neurosci. Ther. 2017, 23, 386–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taoro-Gonzalez, L.; Arenas, Y.M.; Cabrera-Pastor, A.; Felipo, V. Hyperammonemia alters membrane expression of GluA1 and GluA2 subunits of AMPA receptors in hippocampus by enhancing activation of the IL-1 receptor: Underlying mechanisms. J. Neuroinflamm. 2018, 15, 36. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Prieto, G.A.; Cotman, C.W. IL-1β suppresses cLTP-induced surface expression of GluA1 and actin polymerization via ceramide-mediated Src activation. J. Neuroinflamm. 2018, 15, 127. [Google Scholar] [CrossRef]
- Machado, I.; Schiöth, H.B.; Lasaga, M.; Scimonelli, T. IL-1β reduces GluA1 phosphorylation and its surface expression during memory reconsolidation and α-melanocyte-stimulating hormone can modulate these effects. Neuropharmacology 2018, 128, 314–323. [Google Scholar] [CrossRef]
- Gu, X.; Wu, H.; Xie, Y.; Xu, L.; Liu, X.; Wang, W. Caspase-1/IL-1β represses membrane transport of GluA1 by inhibiting the interaction between Stargazin and GluA1 in Alzheimer’s disease. Mol. Med. 2021, 27, 8. [Google Scholar] [CrossRef]
- Viviani, B.; Bartesaghi, S.; Gardoni, F.; Vezzani, A.; Behrens, M.M.; Bartfai, T.; Binaglia, M.; Corsini, E.; Di Luca, M.; Galli, C.L.; et al. Interleukin-1β enhances NMDA receptor-mediated intracellular calcium increase through activation of the Src family of kinases. J. Neurosci. 2003, 23, 8692–8700. [Google Scholar] [CrossRef]
- Viviani, B.; Gardoni, F.; Bartesaghi, S.; Corsini, E.; Facchi, A.; Galli, C.L.; Di Luca, M.; Marinovich, M. Interleukin-1β released by gp120 drives neural death through tyrosine phosphorylation and trafficking of NMDA receptors. J. Biol. Chem. 2006, 281, 30212–30222. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Adams, M.M.; Long, A.; Carter, C.C.; Bennett, C.; Sonntag, W.E.; Nicolle, M.M.; Robbins, M.; D’Agostino, R., Jr.; Brunso-Bechtold, J.K. Spatial learning and memory deficits after whole-brain irradiation are associated with changes in NMDA receptor subunits in the hippocampus. Radiat. Res. 2006, 166, 892–899. [Google Scholar] [CrossRef] [Green Version]
- Collingridge, G.L.; Volianskis, A.; Bannister, N.; France, G.; Hanna, L.; Mercier, M.; Tidball, P.; Fang, G.; Irvine, M.W.; Costa, B.M. The NMDA receptor as a target for cognitive enhancement. Neuropharmacology 2015, 512, 49–53. [Google Scholar] [CrossRef] [Green Version]
- Sanz-Clemente, A.; Nicoll, R.A.; Roche, K.W. Diversity in NMDA receptor composition: Many regulators, many consequences. Neuroscientist 2013, 19, 62–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, L.; Chu, P.; Huang, P.; Guo, Y.; Lv, Y. Effect of vitamin D on the learning and memory ability of FGR rat and NMDA receptor expression in hippocampus. Exp. Ther. Med. 2017, 14, 581–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Yang, L.; Bu, X.; Tang, J.; Fan, X. NMDA receptor subunit and CaMKII changes in rat hippocampus by congenital HCMV infection: A mechanism for learning and memory impairment. Neuroreport 2017, 28, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Shipton, O.A.; Paulsen, O. NMDA receptors in hippocampal plasticity. Philos. Trans. R. Soc. 2014, 369, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Cercato, M.C.; Colettis, N.; Snitcofsky, M.; Aguirre, A.I.; Kornisiuk, E.E.; Baez, M.V.; Jerusalinsky, D.A. Hippocampal NMDA receptors and the previous experience effect on memory. J. Physiol.-Paris 2014, 108, 263–269. [Google Scholar] [CrossRef]
- Louveau, A.; Angibaud, J.; Haspot, F.; Opazo, M.C.; Thinard, R.; Thepenier, V.; Baudouin, S.J.; Lescaudron, L.; Hulin, P.; Riedel, C.A.; et al. Impaired spatial memory in mice lacking CD3ζ is associated with altered NMDA and AMPA receptors signaling independent of T-Cell deficiency. J. Neurosci. 2013, 33, 18672–18685. [Google Scholar] [CrossRef] [Green Version]
- Monfort, P.; Erceg, S.; Piedrafita, B.; Llansola, M.; Felipo, V. Chronic liver failure in rats impairs glutamatergic synaptic transmission and long-term potentiation in hippocampus and learning ability. Eur. J. Neurosci. 2007, 25, 2103–2111. [Google Scholar] [CrossRef]
- Ferro, D.; Baratta, F.; Pastori, D.; Cocomello, N.; Colantoni, A.; Angelico, F.; Del Ben, M. New Insights into the Pathogenesis of Non-Alcoholic Fatty Liver Disease: Gut-Derived Lipopolysaccharides and Oxidative Stress. Nutrients 2020, 12, 2762. [Google Scholar] [CrossRef]
- Brown, G.C. The endotoxin hypothesis of neurodegeneration. J. Neuroinflamm. 2019, 16, 180. [Google Scholar] [CrossRef] [Green Version]
- Zakaria, R.; Wan Yaacob, W.M.; Othman, Z.; Long, I.; Ahmad, A.H.; Al-Rahbi, B. Lipopolysaccharide-induced memory impairment in rats: A model of Alzheimer’s disease. Physiol. Res. 2017, 66, 553–565. [Google Scholar] [CrossRef]
- Zhao, H.-W.; Zhang, Z.-F.; Chai, X.; Li, G.-Q.; Cui, H.-R.; Wang, H.-B.; Meng, Y.-K.; Liu, H.-M.; Wang, J.-B.; Li, R.-S.; et al. Oxymatrine attenuates CCl4-induced hepatic fibrosis via modulation of TLR4-dependent inflammatory and TGF-β1 signaling pathways. Int. Immunopharmacol. 2016, 36, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Schippers, M.; Post, E.; Eichhorn, I.; Langeland, J.; Beljaars, L.; Malo, M.S.; Hodin, R.A.; Millán, J.L.; Popov, Y.; Schuppan, D.; et al. Phosphate Groups in the Lipid A Moiety Determine the Effects of LPS on Hepatic Stellate Cells: A Role for LPS Dephosphorylating Activity in Liver Fibrosis. Cells 2020, 9, 2708. [Google Scholar] [CrossRef] [PubMed]
- Lieber, C.S. Role of oxidative stress and antioxidant therapy in alcoholic and nonalcoholic liver diseases. Adv. Pharmacol. 1997, 38, 601–628. [Google Scholar] [CrossRef] [PubMed]
- Ionescu-Tucker, A.; Cotman, C.W. Emerging roles of oxidative stress in brain aging and Alzheimer’s disease. Neurobiol. Aging 2021, 107, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.T.; Leu, D.; Zou, Y. Oxidative stress and redox regulation on hippocampal-dependent cognitive functions. Arch. Biochem. Biophys. 2015, 576, 2–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montgomery, J.Y.; Bajaj, J.S. Advances in the evaluation and management of minimal hepatic encephalopathy. Curr. Gastroenterol. Rep. 2011, 13, 26–33. [Google Scholar] [CrossRef]
- Gimenez-Garzó, C.; Urios, A.; Agustí, A.; González-López, O.; Escudero-García, D.; Escudero-Sanchis, A.; Serra, M.A.; Giner-Durán, R.; Montoliu, C.; Felipo, V. Is cognitive impairment in cirrhotic patients due to increased peroxynitrite and oxidative stress? Antioxid. Redox Signal. 2015, 22, 871–877. [Google Scholar] [CrossRef]
- Giménez-Garzó, C.; Urios, A.; Agustí, A.; Mangas-Losada, A.; García-García, R.; Escudero-García, D.; Kosenko, E.; Ordoño, J.F.; Tosca, J.; Giner-Durán, R.; et al. Cirrhotic patients with minimal hepatic encephalopathy have increased capacity to eliminate superoxide and peroxynitrite in lymphocytes, associated with cognitive impairment. Free Radic. Res. 2018, 52, 118–133. [Google Scholar] [CrossRef]
- Mousa, N.; Abdel-Razik, A.; Zaher, A.; Hamed, M.; Shiha, G.; Effat, N.; Elbaz, S.; Elhelaly, R.; Hafez, M.; El-Wakeel, N.; et al. The role of antioxidants and zinc in minimal hepatic encephalopathy: A randomized trial. Ther. Adv. Gastroenterol. 2016, 9, 684–691. [Google Scholar] [CrossRef] [Green Version]
- Ahluwalia, V.; Wade, J.B.; Heuman, D.M.; Hammeke, T.A.; Sanyal, A.J.; Sterling, R.K.; Stravitz, R.T.; Luketic, V.; Siddiqui, M.S.; Puri, P.; et al. Enhancement of functional connectivity, working memory and inhibitory control on multi-modal brain MR imaging with Rifaximin in Cirrhosis: Implications for the gut-liver-brain axis. Metab. Brain Dis. 2014, 29, 1017–1025. [Google Scholar] [CrossRef]
- Rai, R.; Saraswat, V.A.; Dhiman, R.K. Gut microbiota: Its role in hepatic encephalopathy. J. Clin. Exp. Hepatol. 2015, 5 (Suppl. S1), S29–S36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Jin, Y.; Zhou, W.; Xiao, T.; Wu, Z.; Su, J.; Gao, H.; Shen, P.; Zheng, B.; Luo, Q.; et al. Rifaximin Modulates the Gut Microbiota to Prevent Hepatic Encephalopathy in Liver Cirrhosis without Impacting the Resistome. Front. Cell. Infect. Microbiol. 2022, 11, 761192. [Google Scholar] [CrossRef] [PubMed]
- Kaji, K.; Takaya, H.; Saikawa, S.; Furukawa, M.; Sato, S.; Kawaratani, H.; Kitade, M.; Moriya, K.; Namisaki, T.; Akahane, T.; et al. Rifaximin ameliorates hepatic encephalopathy and endotoxemia without affecting the gut microbiome diversity. World J. Gastroenterol. 2017, 23, 8355–8366. [Google Scholar] [CrossRef]
- Kaji, K.; Saikawa, S.; Takaya, H.; Fujinaga, Y.; Furukawa, M.; Kitagawa, K.; Ozutsumi, T.; Kaya, D.; Tsuji, Y.; Sawada, Y.; et al. Rifaximin Alleviates Endotoxemia with Decreased Serum Levels of Soluble CD163 and Mannose Receptor and Partial Modification of Gut Microbiota in Cirrhotic Patients. Antibiotics 2020, 9, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odena, G.; Miquel, M.; Serafín, A.; Galan, A.; Morillas, R.; Planas, R.; Bartolí, R. Rifaximin, but not growth factor 1, reduces brain edema in cirrhotic rats. World J. Gastroenterol. 2012, 18, 2084–2091. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Zou, L.; Jagavelu, K.; Simonetto, D.A.; Huebert, R.C.; Jiang, Z.D.; DuPont, H.L.; Shah, V.H. Intestinal decontamination inhibits TLR4 dependent fibronectin-mediated cross-talk between stellate cells and endothelial cells in liver fibrosis in mice. J. Hepatol. 2012, 56, 893–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangarapu, V.; Ince, A.T.; Baysal, B.; Kayar, Y.; Kılıç, U.; Gök, Ö.; Uysal, Ö.; Şenturk, H. Efficacy of rifaximin on circulating endotoxins and cytokines in patients with nonalcoholic fatty liver disease. Eur. J. Gastroenterol. Hepatol. 2015, 27, 840–845. [Google Scholar] [CrossRef]
- Abdel-Razik, A.; Mousa, N.; Shabana, W.; Refaey, M.; Elzehery, R.; Elhelaly, R.; Zalata, K.; Abdelsalam, M.; Eldeeb, A.A.; Awad, M.; et al. Rifaximin in nonalcoholic fatty liver disease: Hit multiple targets with a single shot. Eur. J. Gastroenterol. Hepatol. 2018, 30, 1237–1246. [Google Scholar] [CrossRef]
- Cobbold, J.F.L.; Atkinson, S.; Marchesi, J.R.; Smith, A.; Wai, S.N.; Stove, J.; Shojaee-Moradie, F.; Jackson, N.; Umpleby, A.M.; Fitzpatrick, J.; et al. Rifaximin in non-alcoholic steatohepatitis: An open-label pilot study. Hepatol. Res. 2018, 48, 69–77. [Google Scholar] [CrossRef]
- Fujinaga, Y.; Kawaratani, H.; Kaya, D.; Tsuji, Y.; Ozutsumi, T.; Furukawa, M.; Kitagawa, K.; Sato, S.; Nishimura, N.; Sawada, Y.; et al. Effective Combination Therapy of Angiotensin-II Receptor Blocker and Rifaximin for Hepatic Fibrosis in Rat Model of Nonalcoholic Steatohepatitis. Int. J. Mol. Sci. 2020, 21, 5589. [Google Scholar] [CrossRef]
- Ian, J.; Nie, M.T.; Xiang, B.; Qian, H.; Yin, C.; Zhang, X.; Zhang, M.; Zhu, X.; Xie, W.F. Rifaximin Ameliorates Non-Alcoholic Steatohepatitis in Mice through Regulating gut Microbiome-Related Bile Acids. Front. Pharmacol. 2022, 13, 841132. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CCL4 | C-RIF | CCL4-RIF | |
|---|---|---|---|
| Microglia activation | ↑ | ↑ | Normalized |
| Astrocyte activation | ↑ | Not affected | ↑ |
| Content of IL-1β | ↑ | ↑ | ↑ |
| Content of TNFα | ↑ | Not affected | Normalized |
| Content of CCL2 | ↑ | ↑ | Normalized |
| Macrophages in meninges | ↑ | Not affected | Normalized |
| CD4+ Lymphocytes in meninges | ↑ | Not affected | ↑ |
| Membrane expression of GluA1 | Not affected | Not affected | Not affected |
| Membrane expression of GluA2 | ↑ | ↑ | ↑ |
| Membrane expression of NR1 | ↓ | Not affected | Normalized |
| Membrane expression of NR2A | ↓ | Not affected | Normalized |
| Membrane expression of NR2B | Not affected | Not affected | Not affected |
| Reference Errors | Not affected | Not affected | Not affected |
| Working Errors | Not affected | Not affected | Not affected |
| Learning Index | ↓ | Not affected | Normalized |
| NOL | ↓ | Not affected | Normalized |
| NOR | Not affected | Not affected | Not affected |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leone, P.; Mincheva, G.; Balzano, T.; Malaguarnera, M.; Felipo, V.; Llansola, M. Rifaximin Improves Spatial Learning and Memory Impairment in Rats with Liver Damage-Associated Neuroinflammation. Biomedicines 2022, 10, 1263. https://doi.org/10.3390/biomedicines10061263
Leone P, Mincheva G, Balzano T, Malaguarnera M, Felipo V, Llansola M. Rifaximin Improves Spatial Learning and Memory Impairment in Rats with Liver Damage-Associated Neuroinflammation. Biomedicines. 2022; 10(6):1263. https://doi.org/10.3390/biomedicines10061263
Chicago/Turabian StyleLeone, Paola, Gergana Mincheva, Tiziano Balzano, Michele Malaguarnera, Vicente Felipo, and Marta Llansola. 2022. "Rifaximin Improves Spatial Learning and Memory Impairment in Rats with Liver Damage-Associated Neuroinflammation" Biomedicines 10, no. 6: 1263. https://doi.org/10.3390/biomedicines10061263
APA StyleLeone, P., Mincheva, G., Balzano, T., Malaguarnera, M., Felipo, V., & Llansola, M. (2022). Rifaximin Improves Spatial Learning and Memory Impairment in Rats with Liver Damage-Associated Neuroinflammation. Biomedicines, 10(6), 1263. https://doi.org/10.3390/biomedicines10061263