
Gene Expression-Based Functional Differences between the Bladder Body and Trigonal Urothelium in Adolescent Female Patients with Micturition Dysfunction

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
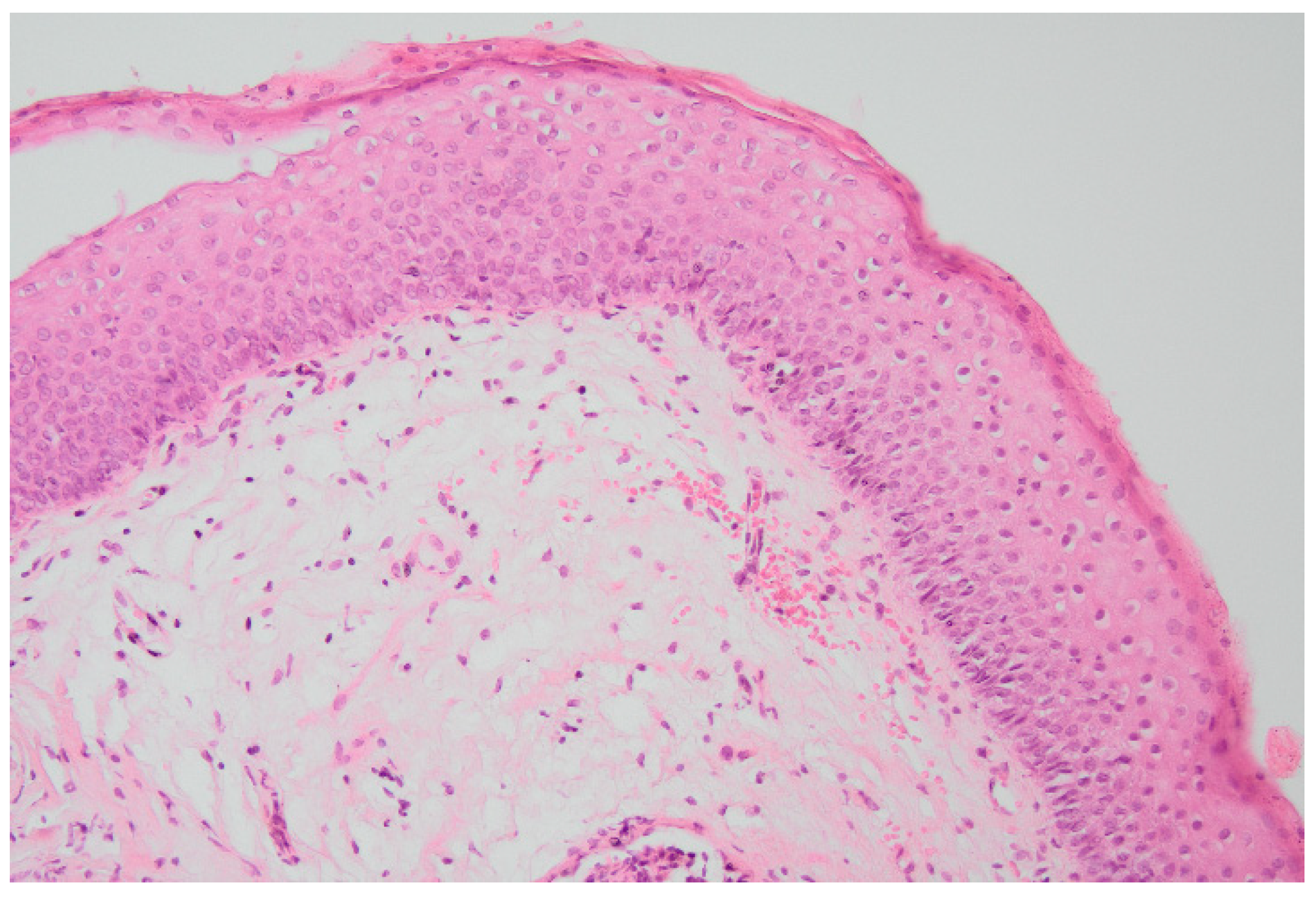
2.2. Subjects and Samples Collection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Age (Years) | Results of Urodynamic Studies | Results of Histopathological Examination |
|---|---|---|---|
| 1B | 12 | Increased sensation in the bladder. High maximal pressure in the bladder during micturition. | Reactive changes in the urothelial epithelium and underlying florid proliferation of von Brunn’s nests |
| 2B | 17 | Not tested | Segmental non-keratinized squamous metaplasia without inflammatory infiltrates |
| 3B | 14 | Mild detrusor overactivity with increased sensation in the bladder | Slight chronic inactive inflammation of the urothelial epithelium with reactive changes and focal squamous non-keratinized metaplasia |
| 4B | 13 | Normal | Moderate chronic active inflammation and swelling of the urothelial epithelium with reactive changes |
| 5B | 17 | Increased sensation in the bladder with reduced bladder capacity. Lower urinary tract dysfunction with urgency sensors. | Focal chronic active inflammation of the urothelial epithelium with reactive changes of the urothelial epithelium and focal non-keratinized squamous metaplasia |
| 6B | 14 | Increased sensation in the bladder with reduced bladder capacity. | Moderate chronic active inflammation of the urothelial epithelium and non-keratinized squamous metaplasia |
| 7B | 17 | Increased sensation in the bladder with reduced bladder capacity. | Chronic inactive inflammation of the urothelial epithelium and visible non-keratinized squamous metaplasia |
| 8B | 18 | Detrusor overactivity with increased sensation in the bladder. Reduced bladder capacity. | Non-keratinized squamous metaplasia * |
| 9B | 17 | Urethral flow with bladder obstruction features. | Non-keratinized squamous metaplasia |
| 10B | 13 | Urethral flow with bladder obstruction features. | Reactive changes of the urothelial epithelium and underlying florid proliferation of von Brunn’s nests |
| 11B | 16 | Detrusor overactivity with increased sensation in the bladder. Reduced bladder capacity. | Normotypic urothelial epithelium with swelling of the stroma and sparse infiltrates of lymphoid cells |
| 12B | 14 | Detrusor overactivity with increased sensation in the bladder. Decreased bladder capacity. | Normotypic urothelial epithelium with swollen and bloodshot stroma |
| 13B | 16 | Detrusor overactivity with increased sensation in the bladder. Reduced bladder capacity. | Non-keratinizing squamous metaplasia |
| 14B | 17 | Detrusor overactivity with increased sensation in the bladder. Reduced bladder capacity. | Non-keratinized squamous metaplasia with poor lymphocyte infiltrates |
| 15B | 13 | Detrusor overactivity with increased sensation in the bladder. Reduced bladder capacity. | Non-keratinized squamous metaplasia and cystitis glandularis |
| 16B | 14 | Detrusor overactivity with increased sensation in the bladder. | Focal active inflammation of the urothelial epithelium and focal non-keratinized squamous metaplasia |
| 17B | 17 | Detrusor overactivity with increased sensation in the bladder | Focal chronic inactive inflammation of the urothelial epithelium and squamous non-keratinized metaplasia |
2.3. RNA Isolation, Sequencing, and Data Analysis
2.4. DNA Extraction, 16S rRNA Sequencing, and Data Analysis
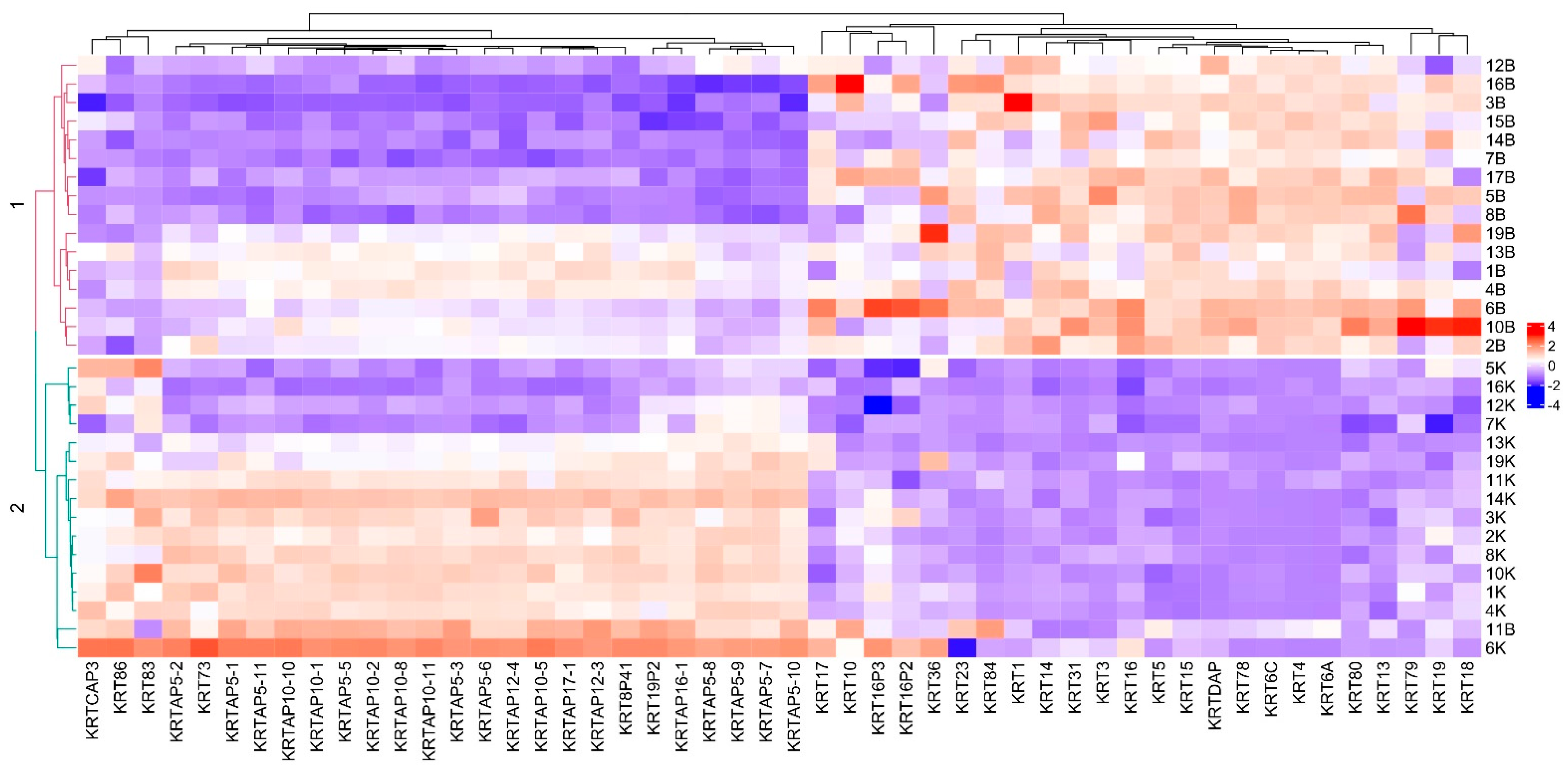
3. Results
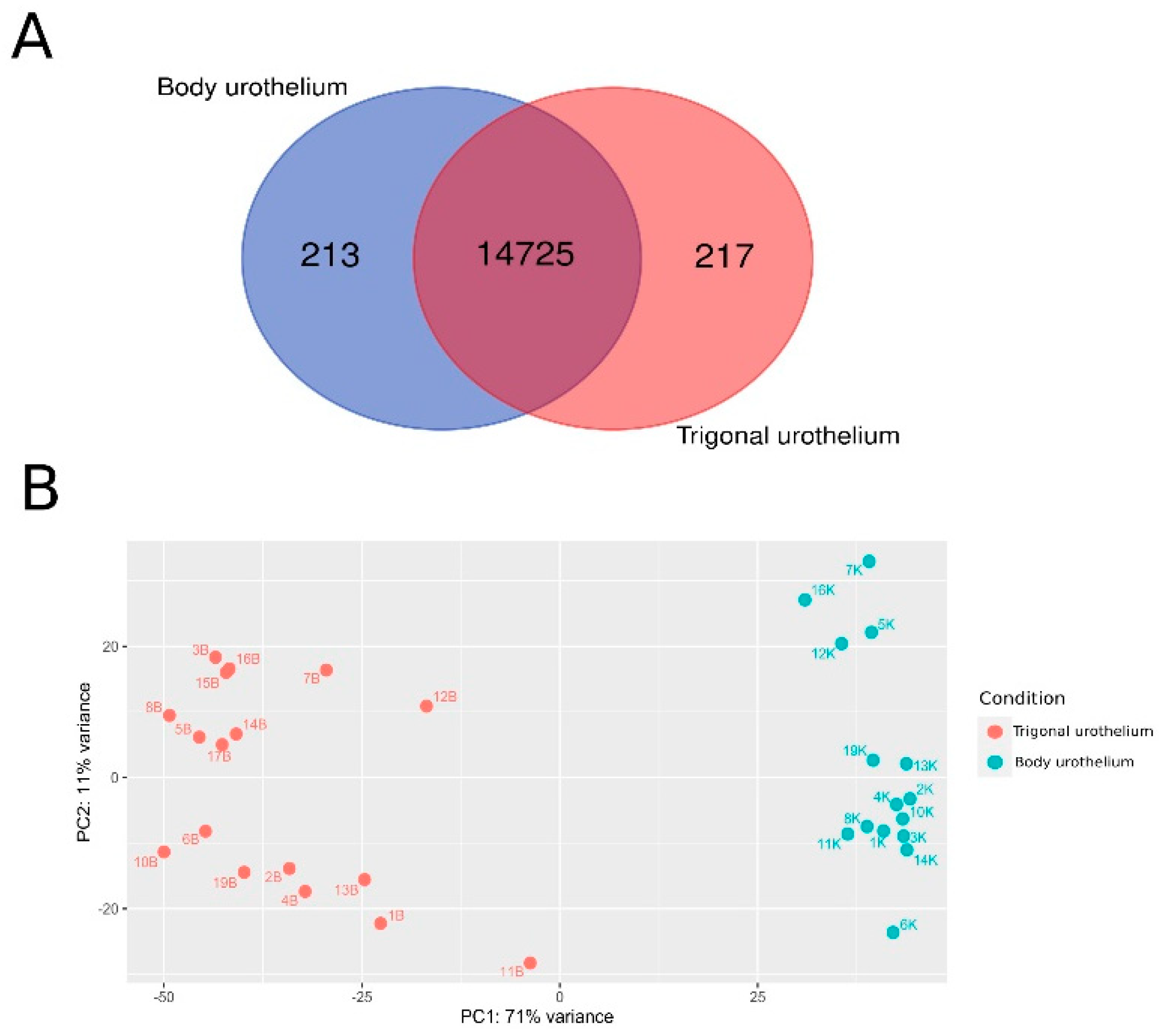
3.1. Transcriptomic Profiling
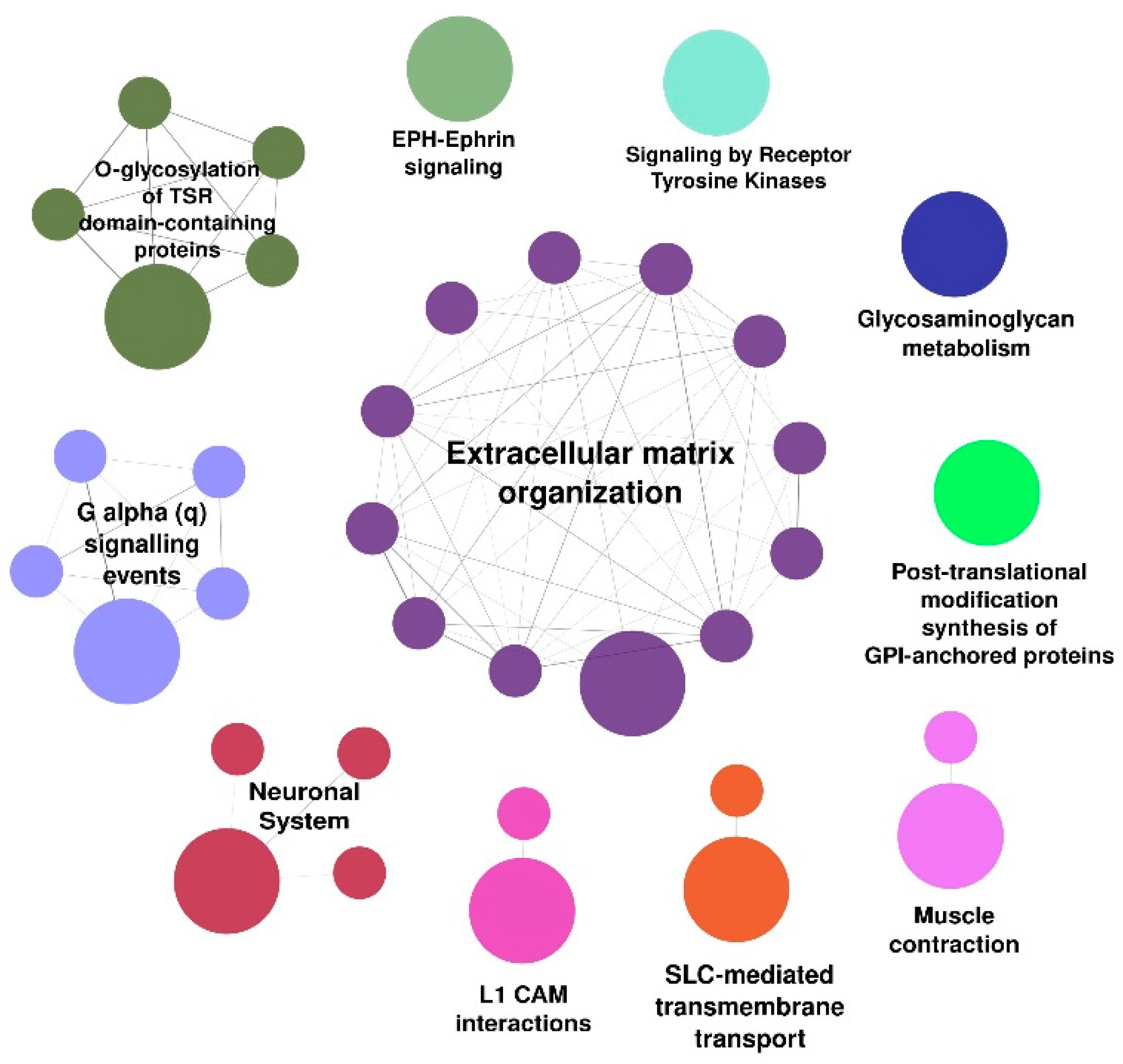
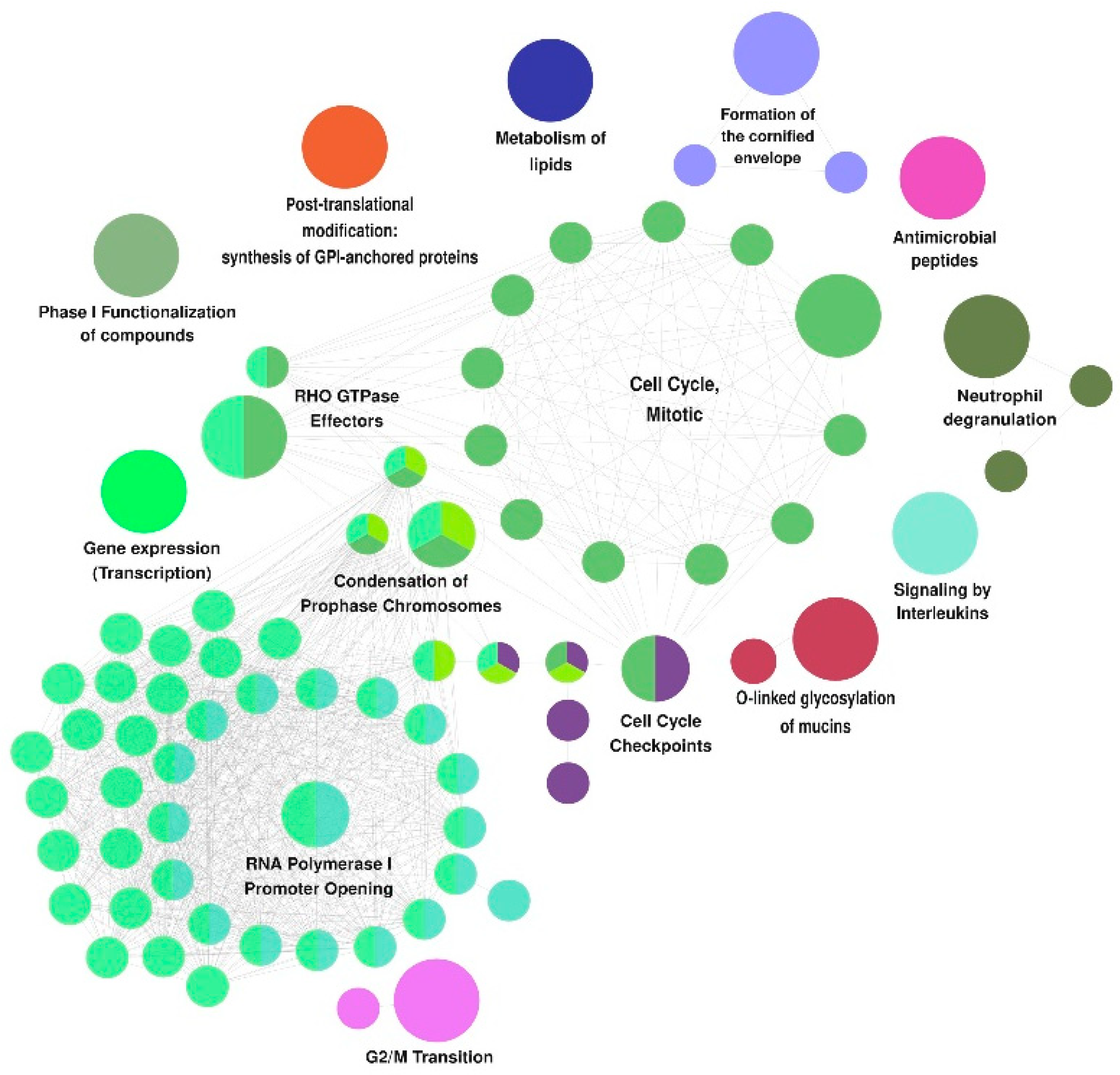
3.2. Functional Annotations in the Reactome Database
3.3. Differential Expression of Selected Genes
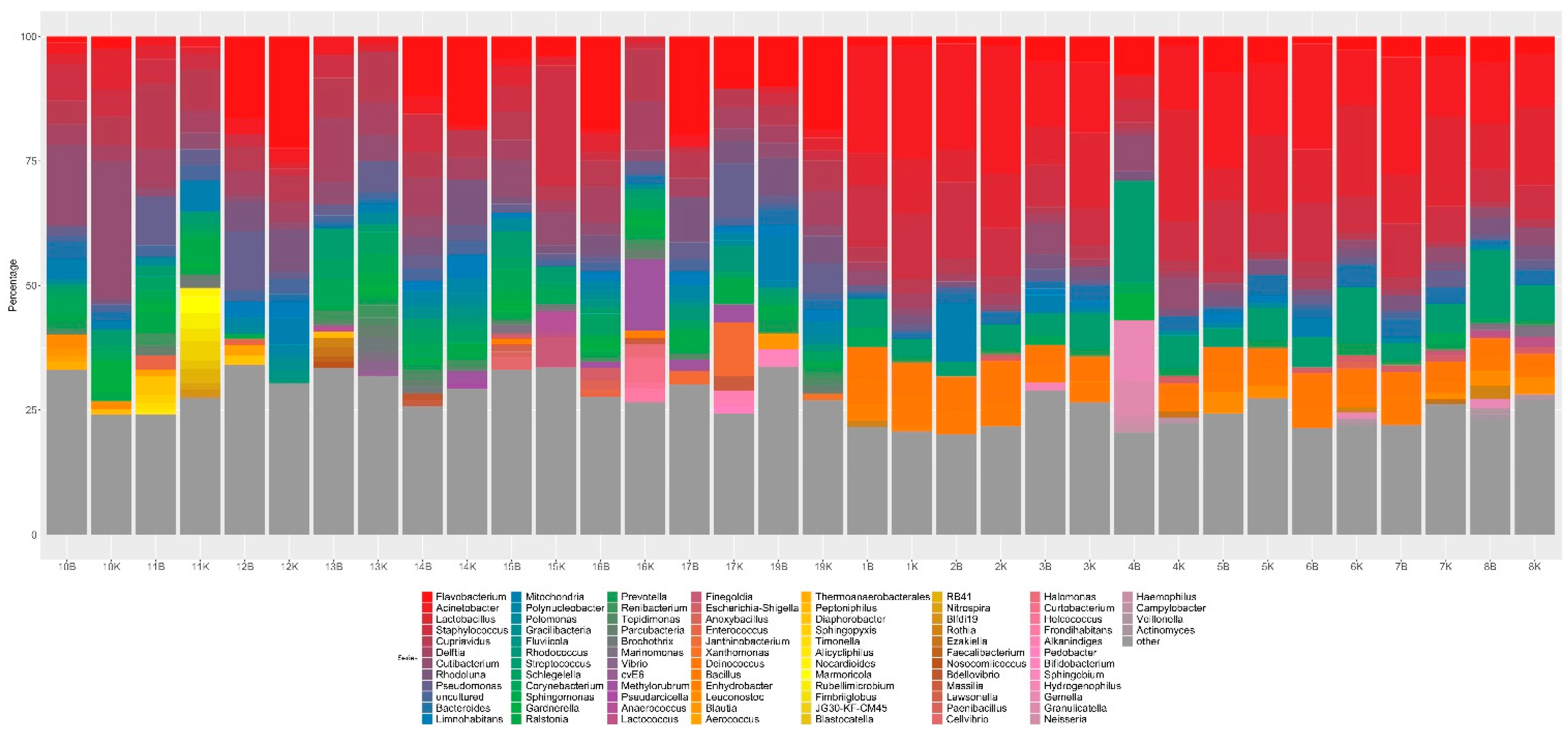
3.4. Mucosal Metagenomics
3.5. Urine Metagenomics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saban, M.R.; Hellmich, H.L.; Turner, M.; Nguyen, N.-B.; Vadigepalli, R.; Dyer, D.W.; Hurst, R.E.; Centola, M.; Saban, R. The Inflammatory and Normal Transcriptome of Mouse Bladder Detrusor and Mucosa. BMC Physiol. 2006, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logadottir, Y.; Delbro, D.; Fall, M.; Gjertsson, I.; Jirholt, P.; Lindholm, C.; Peeker, R. Cytokine Expression in Patients with Bladder Pain Syndrome/Interstitial Cystitis ESSIC Type 3C. J. Urol. 2014, 192, 1564–1568. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.; Han, J.-Y.; Shin, J.H.; Ryu, C.-M.; Yu, H.Y.; Kim, A.; Lee, S.; Lim, J.; Shin, D.-M.; Choo, M.-S. Downregulation of WNT11 Is Associated with Bladder Tissue Fibrosis in Patients with Interstitial Cystitis/Bladder Pain Syndrome without Hunner Lesion. Sci. Rep. 2018, 8, 9782. [Google Scholar] [CrossRef] [PubMed]
- Acharya, P.; Beckel, J.; Ruiz, W.G.; Wang, E.; Rojas, R.; Birder, L.; Apodaca, G. Distribution of the Tight Junction Proteins ZO-1, Occludin, and Claudin-4, -8, and -12 in Bladder Epithelium. Am. J. Physiol. Renal. Physiol. 2004, 287, F305–F318. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.A. Everything You Wanted to Know about the Bladder Epithelium but Were Afraid to Ask. Am. J. Physiol. Renal. Physiol. 2000, 278, F867–F874. [Google Scholar] [CrossRef]
- Fry, C.H.; Vahabi, B. The Role of the Mucosa in Normal and Abnormal Bladder Function. Basic Clin. Pharmacol. Toxicol. 2016, 119 (Suppl. 3), 57–62. [Google Scholar] [CrossRef] [Green Version]
- Merrill, L.; Gonzalez, E.J.; Girard, B.M.; Vizzard, M.A. Receptors, Channels, and Signalling in the Urothelial Sensory System in the Bladder. Nat. Rev. Urol. 2016, 13, 193–204. [Google Scholar] [CrossRef]
- Lv, Y.-S.; Gao, R.; Lin, Q.-M.; Jiang, T.; Chen, Q.; Tang, S.-X.; Mao, H.-P.; Zhou, H.-L.; Cao, L.-S. The Role of TSG-6 and Uroplakin III in Bladder Pain Syndrome/Interstitial Cystitis in Rats and Humans. Iran. J. Basic Med. Sci. 2017, 20, 1242–1249. [Google Scholar] [CrossRef]
- French, L.M.; Bhambore, N. Interstitial Cystitis/Painful Bladder Syndrome. Am. Fam. Physician 2011, 83, 1175–1181. [Google Scholar]
- Cortes, G.A.; Flores, J.L. Physiology, Urination. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Harik, L.R.; O’Toole, K.M. Nonneoplastic Lesions of the Prostate and Bladder. Arch. Pathol. Lab. Med. 2012, 136, 721–734. [Google Scholar] [CrossRef]
- Kyung, Y.S.; Park, H.Y.; Lee, G. Preservation of Uroplakins by 2-Mercaptoethanesulfonate in Cyclophosphamide-Induced Rat Cystitis. Arch. Toxicol. 2011, 85, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Zupancic, D.; Jezernik, K.; Vidmar, G. Effect of Melatonin on Apoptosis, Proliferation and Differentiation of Urothelial Cells after Cyclophosphamide Treatment. J. Pineal. Res. 2008, 44, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Romih, R.; Zupančič, D. Cystitis: From Urothelial Cell Biology to Clinical Applications. Biomed. Res. Int. 2014, 2014, 473536. [Google Scholar] [CrossRef] [Green Version]
- Long, E.D.; Shepherd, R.T. The Incidence and Significance of Vaginal Metaplasia of the Bladder Trigone in Adult Women. Br. J. Urol. 1983, 55, 189–194. [Google Scholar] [CrossRef] [PubMed]
- WebPathology. Available online: http://webpathology.com (accessed on 24 January 2022).
- Sánchez Freire, V.; Burkhard, F.C.; Schmitz, A.; Kessler, T.M.; Monastyrskaya, K. Structural Differences between the Bladder Dome and Trigone Revealed by MRNA Expression Analysis of Cold-Cut Biopsies. BJU Int. 2011, 108, E126–E135. [Google Scholar] [CrossRef] [PubMed]
- Khandelwal, P.; Abraham, S.N.; Apodaca, G. Cell Biology and Physiology of the Uroepithelium. Am. J. Physiol. Renal. Physiol. 2009, 297, F1477–F1501. [Google Scholar] [CrossRef] [Green Version]
- Roosen, A.; Wu, C.; Sui, G.; Chowdhury, R.A.; Patel, P.M.; Fry, C.H. Characteristics of Spontaneous Activity in the Bladder Trigone. Eur. Urol. 2009, 56, 346–353. [Google Scholar] [CrossRef]
- van Ophoven, A.; Oberpenning, F.; Hertle, L. Long-Term Results of Trigone-Preserving Orthotopic Substitution Enterocystoplasty for Interstitial Cystitis. J. Urol. 2002, 167, 603–607. [Google Scholar] [CrossRef]
- Ogawa, T.; Homma, T.; Igawa, Y.; Seki, S.; Ishizuka, O.; Imamura, T.; Akahane, S.; Homma, Y.; Nishizawa, O. CXCR3 Binding Chemokine and TNFSF14 over Expression in Bladder Urothelium of Patients with Ulcerative Interstitial Cystitis. J. Urol. 2010, 183, 1206–1212. [Google Scholar] [CrossRef]
- Grover, S.; Srivastava, A.; Lee, R.; Tewari, A.K.; Te, A.E. Role of Inflammation in Bladder Function and Interstitial Cystitis. Ther. Adv. Urol. 2011, 3, 19–33. [Google Scholar] [CrossRef]
- Offiah, I.; Didangelos, A.; Dawes, J.; Cartwright, R.; Khullar, V.; Bradbury, E.J.; O’Sullivan, S.; Williams, D.; Chessell, I.P.; Pallas, K.; et al. The Expression of Inflammatory Mediators in Bladder Pain Syndrome. Eur. Urol. 2016, 70, 283–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurkiewicz, B.; Załęska-Oracka, K.; Samotyjek, J.; Wajszczuk, E.; Szymanek-Szwed, M. Diagnostics and Symptomatic Treatment of Non-Keratinizing Squamous Cell Metaplasia of the Urinary Bladder in Children—Own Experience. Nephrol. Renal. Dis. 2019, 4, 1–4. [Google Scholar] [CrossRef]
- Lichawska-Cieslar, A.; Pietrzycka, R.; Ligeza, J.; Kulecka, M.; Paziewska, A.; Kalita, A.; Dolicka, D.D.; Wilamowski, M.; Miekus, K.; Ostrowski, J.; et al. RNA Sequencing Reveals Widespread Transcriptome Changes in a Renal Carcinoma Cell Line. Oncotarget 2018, 9, 8597–8613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeber-Lubecka, N.; Kulecka, M.; Ambrozkiewicz, F.; Paziewska, A.; Goryca, K.; Karczmarski, J.; Rubel, T.; Wojtowicz, W.; Mlynarz, P.; Marczak, L.; et al. Limited Prolonged Effects of Rifaximin Treatment on Irritable Bowel Syndrome-Related Differences in the Fecal Microbiome and Metabolome. Gut Microbes 2016, 7, 397–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picard Tools—By Broad Institute. Available online: https://broadinstitute.github.io/picard/ (accessed on 24 January 2022).
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A Versatile Open Source Tool for Metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Paulson, J.N. MetagenomeSeq: Statistical Analysis for Sparse High-Throughput Sequencing. Bioconduct. Package 2013, 1, 191. [Google Scholar]
- Huson, D.H.; Auch, A.F.; Qi, J.; Schuster, S.C. MEGAN Analysis of Metagenomic Data. Genome Res. 2007, 17, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- DeGraff, D.J.; Cates, J.M.; Mauney, J.R.; Clark, P.E.; Matusik, R.; Adam, R.M. When Urothelial Differentiation Pathways Go Wrong: Implications for Bladder Cancer Development and Progression. Urol. Oncol. 2013, 31, 802–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.-R.; Kong, X.-P.; Pellicer, A.; Kreibich, G.; Sun, T.-T. Uroplakins in Urothelial Biology, Function, and Disease. Kidney Int. 2009, 75, 1153–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephenson, T.J.; Henry, L.; Harris, S.C.; Giri, D.D.; Fox, M.; Underwood, J.C. Pseudomembranous Trigonitis of the Bladder: Hormonal Aetiology. J. Clin. Pathol. 1989, 42, 922–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadeghi, Z.; MacLennan, G.; Childs, S.J.; Zimmern, P.E. Is Trigonitis a Neglected, Imprecise, Misunderstood, or Forgotten Diagnosis? LUTS Low. Urin. Tract Symptoms 2019, 11, 182–188. [Google Scholar] [CrossRef] [PubMed]
- De Nisco, N.J.; Neugent, M.; Mull, J.; Chen, L.; Kuprasertkul, A.; de Souza Santos, M.; Palmer, K.L.; Zimmern, P.; Orth, K. Direct Detection of Tissue-Resident Bacteria and Chronic Inflammation in the Bladder Wall of Postmenopausal Women with Recurrent Urinary Tract Infection. J. Mol. Biol. 2019, 431, 4368–4379. [Google Scholar] [CrossRef]
- Haga, R.B.; Ridley, A.J. Rho GTPases: Regulation and Roles in Cancer Cell Biology. Small GTPases 2016, 7, 207–221. [Google Scholar] [CrossRef] [Green Version]
- Jurkiewicz, B.; Matuszewski, Ł.; Bokwa, T.; Gut, G.; Rybak, D. Bladder Squamous Metaplasia of the Urothelium—Introductory Report. Urol. Int. 2006, 77, 46–49. [Google Scholar] [CrossRef]
- Jurkiewicz, B.; Ząbkowski, T. Nonkeratinised Squamous Metaplasia of the Urinary Bladder in Children: A Report of Case Experiences. BioMed. Res. Int. 2014, 2014, e936970. [Google Scholar] [CrossRef]
- Zupančič, D.; Romih, R. Immunohistochemistry as a Paramount Tool in Research of Normal Urothelium, Bladder Cancer and Bladder Pain Syndrome. Eur. J. Histochem. 2021, 65, 3242. [Google Scholar] [CrossRef]
- Kreft, M.E.; Romih, R.; Kreft, M.; Jezernik, K. Endocytotic Activity of Bladder Superficial Urothelial Cells Is Inversely Related to Their Differentiation Stage. Differentiation 2009, 77, 48–59. [Google Scholar] [CrossRef]
- Tratnjek, L.; Romih, R.; Kreft, M. Differentiation-Dependent Rearrangements of Actin Filaments and Microtubules Hinder Apical Endocytosis in Urothelial Cells. Histochem. Cell Biol. 2017, 148, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Deo, P.N.; Deshmukh, R. Pathophysiology of Keratinization. J. Oral Maxillofac. Pathol. 2018, 22, 86. [Google Scholar] [CrossRef] [PubMed]
- Moll, R.; Divo, M.; Langbein, L. The Human Keratins: Biology and Pathology. Histochem. Cell Biol. 2008, 129, 705–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omary, M.B.; Ku, N.O.; Liao, J.; Price, D. Keratin Modifications and Solubility Properties in Epithelial Cells and in Vitro. Subcell. Biochem. 1998, 31, 105–140. [Google Scholar]
- Franke, W.W.; Schiller, D.L.; Moll, R.; Winter, S.; Schmid, E.; Engelbrecht, I.; Denk, H.; Krepler, R.; Platzer, B. Diversity of Cytokeratins. Differentiation Specific Expression of Cytokeratin Polypeptides in Epithelial Cells and Tissues. J. Mol. Biol. 1981, 153, 933–959. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Gao, Z.; Zhang, L.; Chen, L.; Wu, Z.; Liu, Q.; Wang, S.; Zhou, N.; Chai, T.C.; et al. Single-Cell Transcriptomes of Mouse Bladder Urothelium Uncover Novel Cell Type Markers and Urothelial Differentiation Characteristics. Cell Prolif. 2021, 54, e13007. [Google Scholar] [CrossRef]
- Birder, L.; Andersson, K.-E. Urothelial Signaling. Physiol. Rev. 2013, 93, 653–680. [Google Scholar] [CrossRef] [Green Version]
- Chai, T.C.; Russo, A.; Yu, S.; Lu, M. Mucosal Signaling in the Bladder. Auton. Neurosci 2016, 200, 49–56. [Google Scholar] [CrossRef]
- Kanai, A.; Andersson, K.-E. Bladder Afferent Signaling: Recent Findings. J. Urol. 2010, 183, 1288–1295. [Google Scholar] [CrossRef] [Green Version]
- Khakh, B.S.; North, R.A. P2X Receptors as Cell-Surface ATP Sensors in Health and Disease. Nature 2006, 442, 527–532. [Google Scholar] [CrossRef]
- Clapham, D.E.; Julius, D.; Montell, C.; Schultz, G. International Union of Pharmacology. XLIX. Nomenclature and Structure-Function Relationships of Transient Receptor Potential Channels. Pharmacol. Rev. 2005, 57, 427–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romagnani, P.; Lasagni, L.; Annunziato, F.; Serio, M.; Romagnani, S. CXC Chemokines: The Regulatory Link between Inflammation and Angiogenesis. Trends Immunol. 2004, 25, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Cabrero-de Las Heras, S.; Martínez-Balibrea, E. CXC Family of Chemokines as Prognostic or Predictive Biomarkers and Possible Drug Targets in Colorectal Cancer. World J. Gastroenterol. 2018, 24, 4738–4749. [Google Scholar] [CrossRef] [PubMed]
- Zasloff, M. Antimicrobial Peptides, Innate Immunity, and the Normally Sterile Urinary Tract. JASN 2007, 18, 2810–2816. [Google Scholar] [CrossRef] [Green Version]
- Birder, L.A. Urinary Bladder, Cystitis and Nerve/Urothelial Interactions. Auton. Neurosci. 2014, 182, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Graham, E.; Chai, T.C. Dysfunction of Bladder Urothelium and Bladder Urothelial Cells in Interstitial Cystitis. Curr. Urol. Rep. 2006, 7, 440–446. [Google Scholar] [CrossRef]
- Ganz, T. Defensins in the Urinary Tract and Other Tissues. J. Infect. Dis. 2001, 183 (Suppl. 1), S41–S42. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial Peptides of Multicellular Organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef]
- Hanno, P.M.; Fritz, R.W.; Mulholland, S.G.; Wein, A.J. Heparin—Examination of Its Antibacterial Adsorption Properties. Urology 1981, 18, 273–276. [Google Scholar] [CrossRef]
- Erickson, D.R.; Schwarze, S.R.; Dixon, J.K.; Clark, C.J.; Hersh, M.A. Differentiation Associated Changes in Gene Expression Profiles of Interstitial Cystitis and Control Urothelial Cells. J. Urol. 2008, 180, 2681–2687. [Google Scholar] [CrossRef]
- Neuhaus, J.; Berndt-Paetz, M.; Gonsior, A. Biomarkers in the Light of the Etiopathology of IC/BPS. Diagnostics 2021, 11, 2231. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, Y.; Luo, Y.; Hanno, P.M.; Maeda, D.; Homma, Y. Interstitial Cystitis/Bladder Pain Syndrome: The Evolving Landscape, Animal Models and Future Perspectives. Int. J. Urol. 2020, 27, 491–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inal-Gultekin, G.; Gormez, Z.; Mangir, N. Defining Molecular Treatment Targets for Bladder Pain Syndrome/Interstitial Cystitis: Uncovering Adhesion Molecules. Front. Pharmacol. 2022, 13, 780855. [Google Scholar] [CrossRef] [PubMed]






| Upregulated in Trigonal Urothelium | Upregulated in Body Urothelium | ||||
|---|---|---|---|---|---|
| Gene | padj-Value | FC | Gene | padj-Value | FC |
| CXCL1 | 1.57 × 10−49 | 386.85 | TAC1 | 5.49 × 10−17 | 47.38 |
| CXCL6 | 1.67 × 10−63 | 70.71 | CCL15 | 1.15 × 10−33 | 16.37 |
| CXCL3 | 1.00 × 10−15 | 34.22 | TH | 3.75 × 10−08 | 6.80 |
| CXCL2 | 1.02 × 10−15 | 28.83 | P2RX2 | 3.24 × 10−02 | 5.48 |
| CXCR1 | 4.20 × 10−05 | 9.46 | TRPV3 | 1.86 × 10−12 | 5.43 |
| CALCA | 1.93 × 10−02 | 8.08 | TRPA1 | 1.82 × 10−11 | 5.22 |
| CXCL17 | 1.69 × 10−25 | 7.51 | VIPR1 | 1.40 × 10−15 | 4.99 |
| CCL22 | 1.92 × 10−11 | 6.33 | CCL13 | 3.21 × 10−04 | 4.39 |
| CCL17 | 2.95 × 10−14 | 6.05 | CCL25 | 1.05 × 10−02 | 4.30 |
| CXCR2 | 4.18 × 10−27 | 5.77 | CCL16 | 4.54 × 10−02 | 3.96 |
| CCL7 | 1.12 × 10−02 | 5.12 | CCL27 | 8.98 × 10−04 | 3.71 |
| P2RY2 | 2.57 × 10−17 | 3.76 | CCL14 | 1.10 × 10−03 | 3.69 |
| TRPM6 | 7.46 × 10−04 | 2.95 | CHRM4 | 8.34 × 10−05 | 3.58 |
| CCL2 | 8.29 × 10−06 | 2.89 | P2RX6 | 3.51 × 10−05 | 3.40 |
| P2RY1 | 4.45 × 10−06 | 2.52 | CXCL12 | 4.10 × 10−07 | 3.20 |
| CXCL10 | 1.07 × 10−02 | 1.95 | CCL18 | 4.73 × 10−02 | 2.63 |
| CXCL16 | 4.22 × 10−04 | 1.74 | P2RX4 | 4.08 × 10−15 | 2.59 |
| CX3CL1 | 7.44 × 10−03 | 1.61 | CXCR2P1 | 1.26 × 10−02 | 2.41 |
| CXCR7 | 2.31 × 10−05 | 2.36 | |||
| P2RY4 | 1.01 × 10−02 | 2.27 | |||
| P2RX1 | 1.80 × 10−02 | 2.18 | |||
| P2RX5 | 3.50 × 10−02 | 2.15 | |||
| P2RY8 | 1.84 × 10−02 | 2.03 | |||
| TRPV2 | 8.50 × 10−04 | 1.81 | |||
| CXCR4 | 3.51 × 10-02 | 1.50 | |||
| TRPM7 | 5.16 × 10−05 | 1.40 | |||
| P2RX7 | 3.80 × 10−02 | 1.40 | |||
| TRPM4 | 4.04 × 10−02 | 1.38 | |||
| No. of Control Samples | No. of Patient Samples | FisherP | FisherAdjP | logFC | se | p-Value | padj-Value | Size | Taxonomic Classification |
|---|---|---|---|---|---|---|---|---|---|
| 0 | 11 | 1.83 × 10−08 | 1.66 × 10−05 | 3.81 | 0.40 | 0.00 | 0.00 | 60,407 | Aminobacter (100) |
| 12 | 10 | 1.99 × 10−01 | 1.00 × 1000 | 3.92 | 0.66 | 2.12 × 10−09 | 9.59 × 10−07 | 51,881 | Comamonadaceae_unclassified (100) |
| 0 | 7 | 1.62 × 10−04 | 2.45 × 10−02 | 1.29 | 0.29 | 5.94 × 10−06 | 1.79 × 10−03 | 2783 | Variovorax (100) |
| 13 | 4 | 1.32 × 10−01 | 1.00 × 1000 | −4.52 | 1.05 | 1.85 × 10−05 | 4.19 × 10−03 | 855 | Eukaryota_unclassified(100) |
| 16 | 1 | 9.04 × 10−05 | 2.45 × 10−02 | −1.53 | 0.39 | 9.09 × 10−05 | 1.65 × 10−02 | 1440 | Cupriavidus (100) |
| 15 | 0 | 5.00 × 10−05 | 2.27 × 10−02 | −1.82 | 0.53 | 5.58 × 10−04 | 7.22 × 10−02 | 22,283 | Mitochondria_ge (100) |
| 13 | 0 | 3.17 × 10−04 | 3.60 × 10−02 | −1.53 | 0.44 | 5.27 × 10−04 | 7.22 × 10−02 | 1344 | Mesorhizobium (100) |
| 13 | 0 | 3.17 × 10−04 | 3.60 × 10−02 | −1.44 | 0.42 | 6.66 × 10−04 | 7.53 × 10−02 | 1520 | Amaricoccus (100) |
| 19 | 4 | 1.62 × 10−04 | 2.45 × 10−02 | −0.45 | 443,343 | Gardnerella (100) | |||
| 19 | 4 | 1.62 × 10−04 | 2.45 × 10−02 | −2.99 | 8891 | Muribaculaceae_ge (100) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeber-Lubecka, N.; Kulecka, M.; Załęska-Oracka, K.; Dąbrowska, M.; Bałabas, A.; Hennig, E.E.; Szymanek-Szwed, M.; Mikula, M.; Jurkiewicz, B.; Ostrowski, J. Gene Expression-Based Functional Differences between the Bladder Body and Trigonal Urothelium in Adolescent Female Patients with Micturition Dysfunction. Biomedicines 2022, 10, 1435. https://doi.org/10.3390/biomedicines10061435
Zeber-Lubecka N, Kulecka M, Załęska-Oracka K, Dąbrowska M, Bałabas A, Hennig EE, Szymanek-Szwed M, Mikula M, Jurkiewicz B, Ostrowski J. Gene Expression-Based Functional Differences between the Bladder Body and Trigonal Urothelium in Adolescent Female Patients with Micturition Dysfunction. Biomedicines. 2022; 10(6):1435. https://doi.org/10.3390/biomedicines10061435
Chicago/Turabian StyleZeber-Lubecka, Natalia, Maria Kulecka, Katarzyna Załęska-Oracka, Michalina Dąbrowska, Aneta Bałabas, Ewa E. Hennig, Magdalena Szymanek-Szwed, Michał Mikula, Beata Jurkiewicz, and Jerzy Ostrowski. 2022. "Gene Expression-Based Functional Differences between the Bladder Body and Trigonal Urothelium in Adolescent Female Patients with Micturition Dysfunction" Biomedicines 10, no. 6: 1435. https://doi.org/10.3390/biomedicines10061435
APA StyleZeber-Lubecka, N., Kulecka, M., Załęska-Oracka, K., Dąbrowska, M., Bałabas, A., Hennig, E. E., Szymanek-Szwed, M., Mikula, M., Jurkiewicz, B., & Ostrowski, J. (2022). Gene Expression-Based Functional Differences between the Bladder Body and Trigonal Urothelium in Adolescent Female Patients with Micturition Dysfunction. Biomedicines, 10(6), 1435. https://doi.org/10.3390/biomedicines10061435