
N-3 PUFA Ameliorates the Gut Microbiota, Bile Acid Profiles, and Neuropsychiatric Behaviours in a Rat Model of Geriatric Depression





Abstract
:1. Introduction
2. Experimental Section
2.1. Animals, Diets, and Experimental Design
2.2. CUMS Model
2.3. Detection of Biochemical Parameters and Inflammatory Cytokines
2.4. Behavioural Tests
2.5. Faecal Microbiota Analysis
2.6. Microbiota-Derived Bile Acid Analysis
2.7. Statistical Analysis
3. Results
3.1. Biochemical and Inflammatory Status
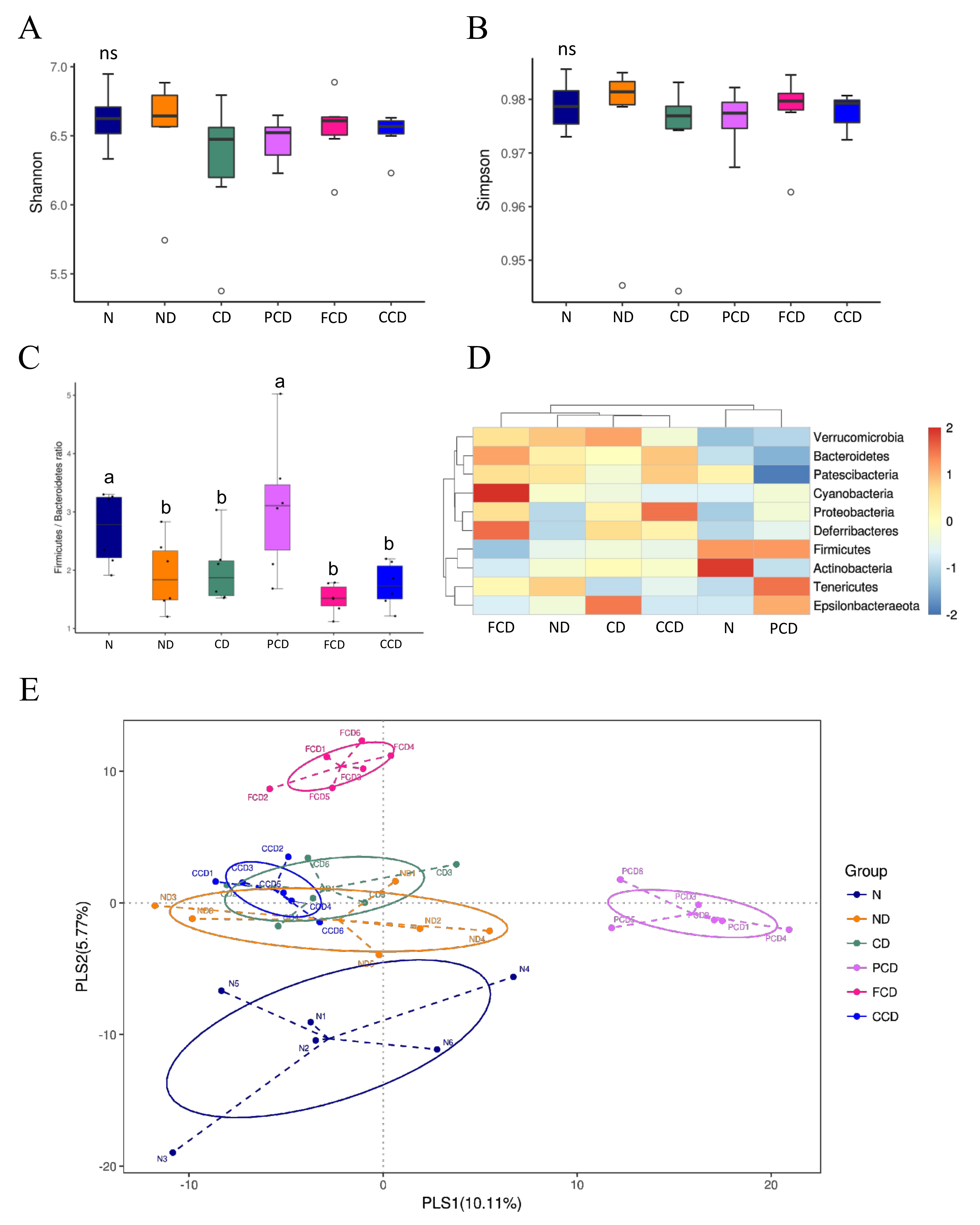
3.2. Between-Group Comparison of Microbial Diversity
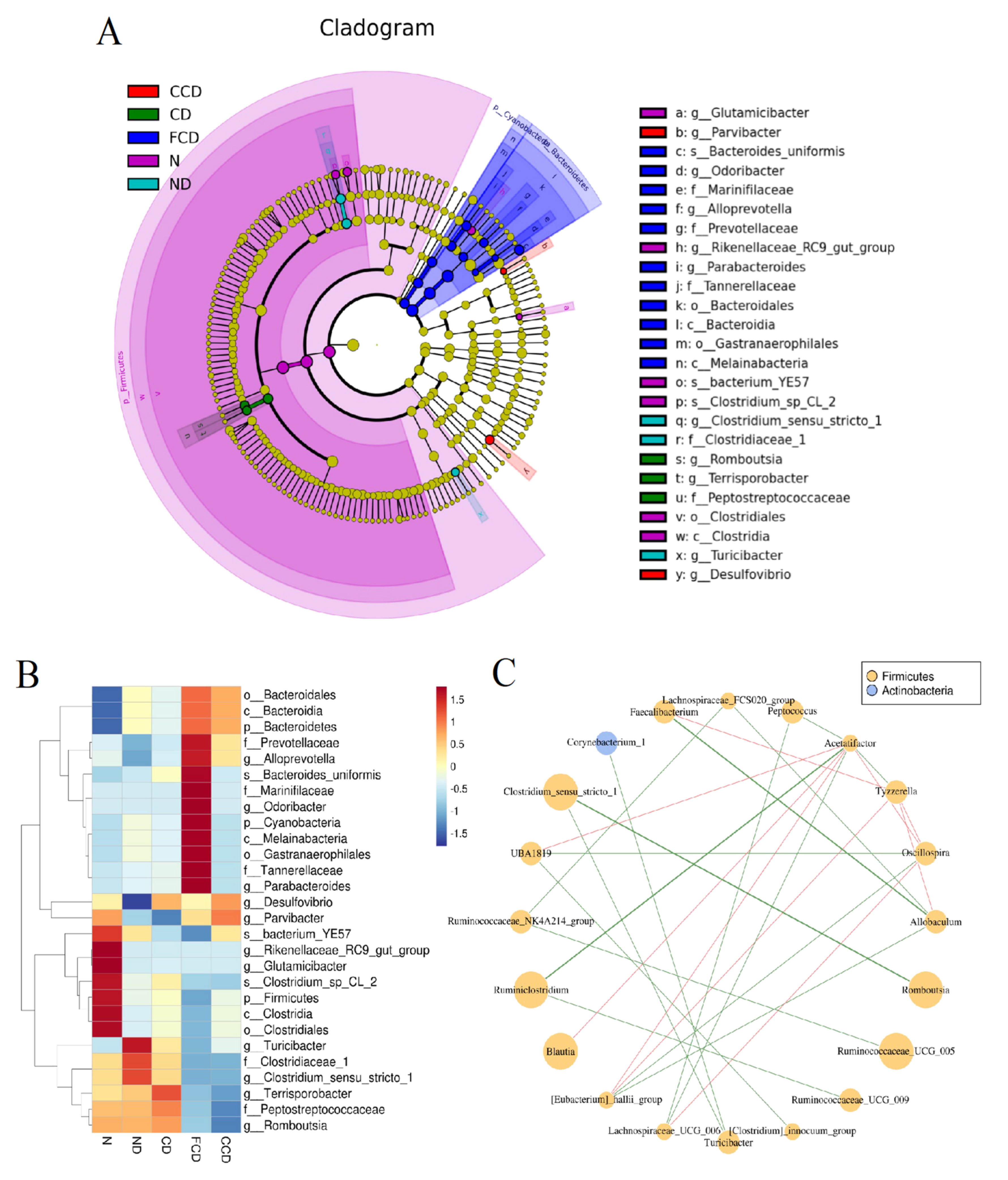
3.3. Between-Group Comparison of Microbiota Composition
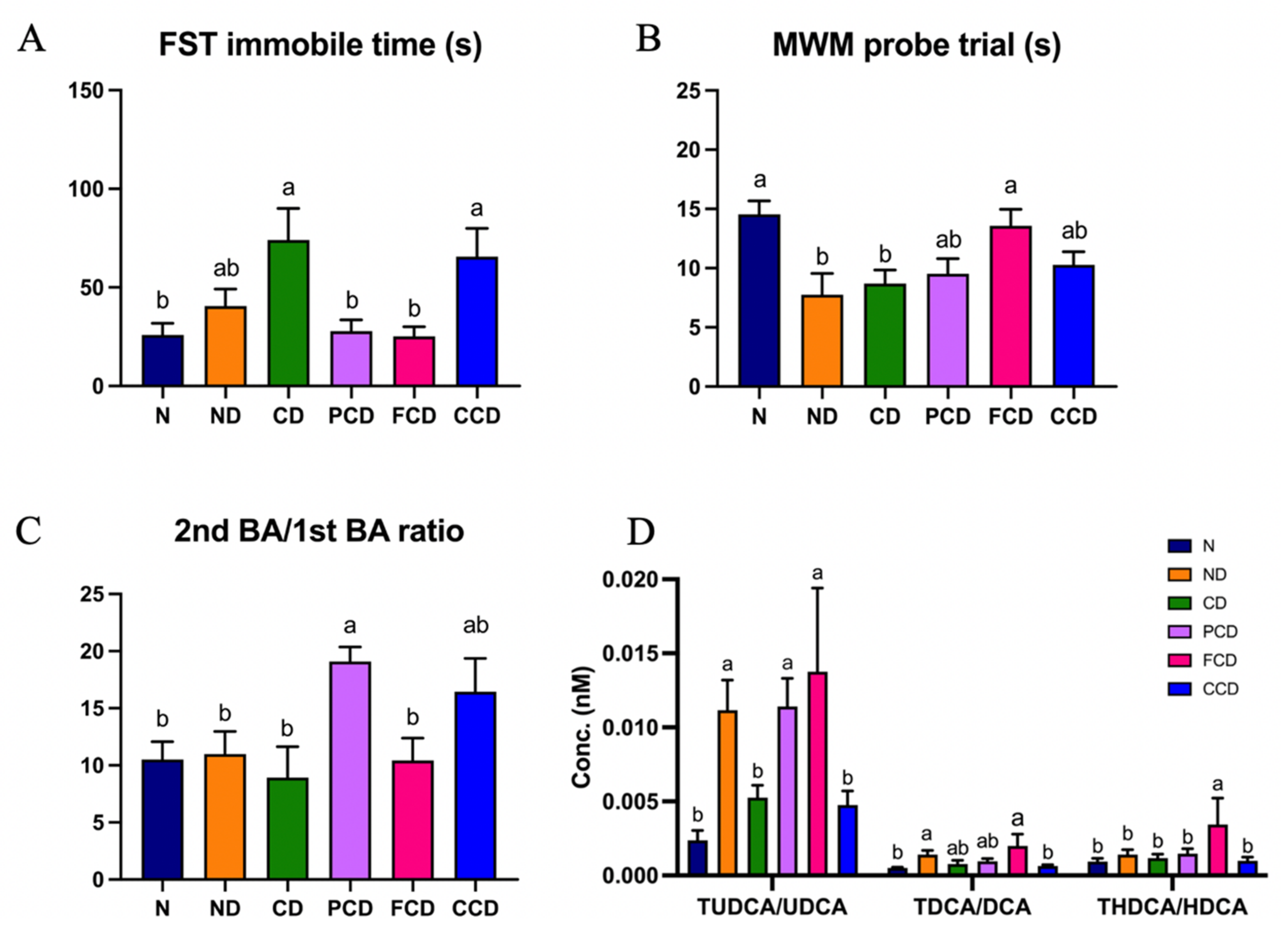
3.4. Bile Acid Profile Changes Induced by Fish Oil
3.5. Neuropsychiatric Behavioural Outcomes and Bile Acid Metabolites
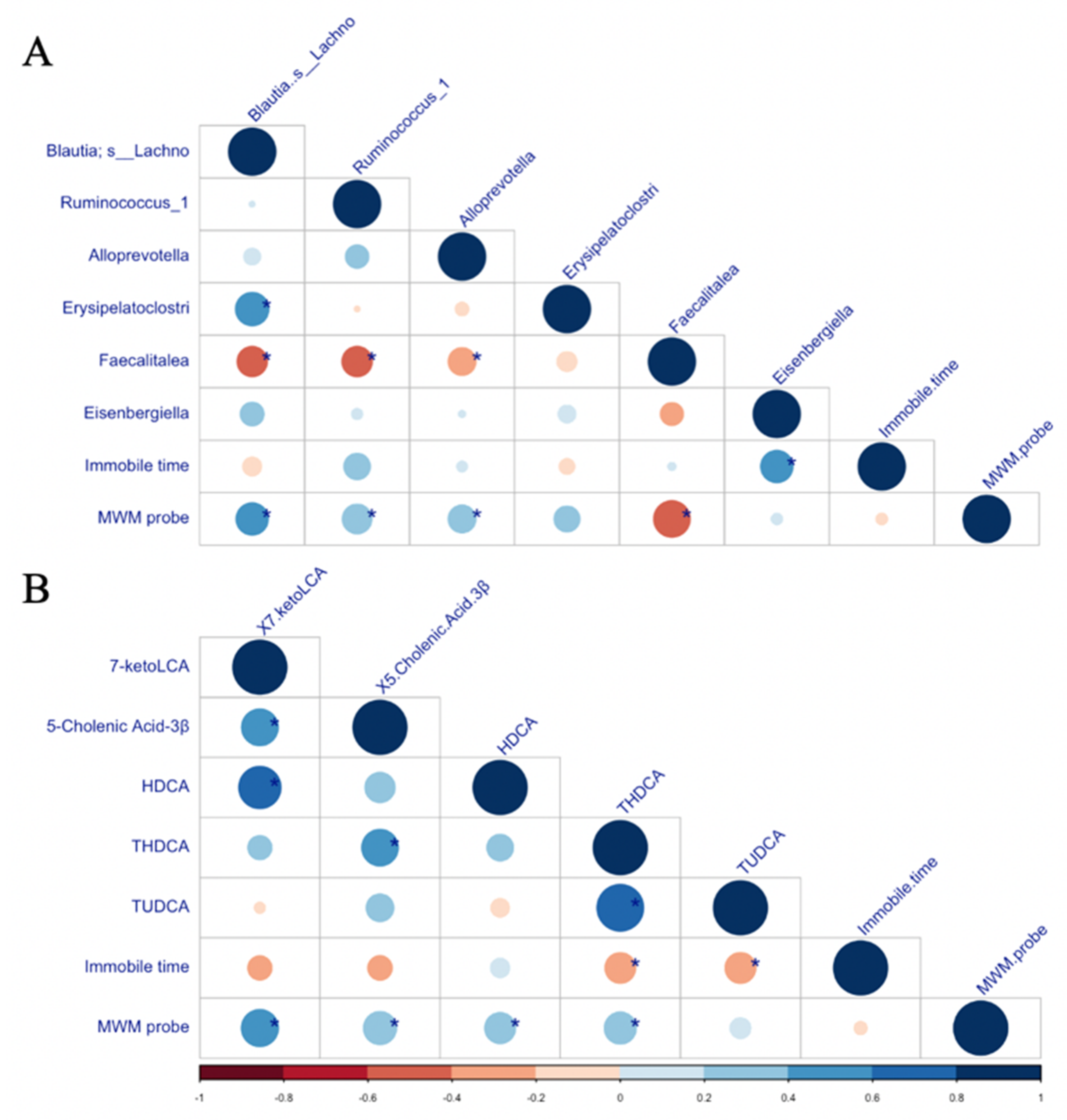
3.6. Correlations between Neurobehavioural Outcomes and Microbiota/Bile Acid Profile Characteristics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gentile, C.L.; Weir, T.L. The gut microbiota at the intersection of diet and human health. Science 2018, 362, 776–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grüner, N.; Mattner, J. Bile Acids and Microbiota: Multifaceted and Versatile Regulators of the Liver–Gut Axis. Int. J. Mol. Sci. 2021, 22, 1397. [Google Scholar] [CrossRef] [PubMed]
- Rudzki, L.; Maes, M. The Microbiota-Gut-Immune-Glia (MGIG) Axis in Major Depression. Mol. Neurobiol. 2020, 57, 4269–4295. [Google Scholar] [CrossRef] [PubMed]
- Wiley, N.C.; Dinan, T.G.; Ross, R.P.; Stanton, C.; Clarke, G.; Cryan, J.F. The microbiota-gut-brain axis as a key regulator of neural function and the stress response: Implications for human and animal health. J. Anim. Sci. 2017, 95, 3225–3246. [Google Scholar] [PubMed]
- Lai, W.-D.; Tung, T.-H.; Teng, C.-Y.; Chang, C.-H.; Chen, Y.-C.; Huang, H.-Y.; Lee, H.-C.; Huang, S.-Y. Fish oil ameliorates neuropsychiatric behaviors and gut dysbiosis by elevating selected microbiota–derived metabolites and tissue tight junctions in rats under chronic sleep deprivation. Food Funct. 2022, 13, 2662–2680. [Google Scholar] [CrossRef]
- Braniste, V.; Al-Asmakh, M.; Kowal, C.; Anuar, F.; Abbaspour, A.; Tóth, M.; Korecka, A.; Bakocevic, N.; Ng, L.G.; Kundu, P.; et al. The gut microbiota influences blood-brain barrier permeability in mice. Sci. Transl. Med. 2014, 6, 263ra158. [Google Scholar] [CrossRef] [Green Version]
- Kealy, J.; Greene, C.; Campbell, M. Blood-brain barrier regulation in psychiatric disorders. Neurosci. Lett. 2018, 726, 133664. [Google Scholar] [CrossRef] [Green Version]
- Baier, J.; Gänsbauer, M.; Giessler, C.; Arnold, H.; Muske, M.; Schleicher, U.; Lukassen, S.; Ekici, A.B.; Rauh, M.; Daniel, C.; et al. Arginase impedes the resolution of colitis by altering the microbiome and metabolome. J. Clin. Investig. 2020, 130, 5703–5720. [Google Scholar] [CrossRef]
- Selwyn, F.P.; Csanaky, I.L.; Zhang, Y.; Klaassen, C.D. Importance of Large Intestine in Regulating Bile Acids and Glucagon-Like Peptide-1 in Germ-Free Mice. Drug Metab. Dispos. 2015, 43, 1544–1556. [Google Scholar] [CrossRef] [Green Version]
- Kimmel, M.; Jin, W.; Xia, K.; Lun, K.; Azcarate-Peril, A.; Plantinga, A.; Wu, M.; Ataei, S.; Rackers, H.; Carroll, I.; et al. Metabolite trajectories across the perinatal period and mental health: A preliminary study of tryptophan-related metabolites, bile acids and microbial composition. Behav. Brain Res. 2021, 418, 113635. [Google Scholar] [CrossRef]
- Li, Z.; Lai, J.; Zhang, P.; Ding, J.; Jiang, J.; Liu, C.; Huang, H.; Zhen, H.; Xi, C.; Sun, Y.; et al. Multi-omics analyses of serum metabolome, gut microbiome and brain function reveal dysregulated microbiota-gut-brain axis in bipolar depression. Mol. Psychiatry 2022, 1–13. [Google Scholar] [CrossRef]
- Yoshimoto, S.; Loo, T.M.; Atarashi, K.; Kanda, H.; Sato, S.; Oyadomari, S.; Iwakura, Y.; Oshima, K.; Morita, H.; Hattori, M.; et al. Obesity-induced gut microbial metabolite promotes liver cancer through senescence secretome. Nature 2013, 499, 97–101. [Google Scholar] [CrossRef]
- Wahlström, A.; Sayin, S.I.; Marschall, H.-U.; Bäckhed, F. Intestinal Crosstalk between Bile Acids and Microbiota and Its Impact on Host Metabolism. Cell Metab. 2016, 24, 41–50. [Google Scholar] [CrossRef] [Green Version]
- MahmoudianDehkordi, S.; Arnold, M.; Nho, K.; Ahmad, S.; Jia, W.; Xie, G.; Louie, G.; Kueider-Paisley, A.; Moseley, M.A.; Thompson, J.W.; et al. Altered bile acid profile associates with cognitive impairment in Alzheimer’s disease—An emerging role for gut microbiome. Alzheimer’s Dement. 2019, 15, 76–92. [Google Scholar] [CrossRef]
- Abdelkader, N.F.; Safar, M.M.; Salem, H.A. Ursodeoxycholic Acid Ameliorates Apoptotic Cascade in the Rotenone Model of Parkinson’s Disease: Modulation of Mitochondrial Perturbations. Mol. Neurobiol. 2014, 53, 810–817. [Google Scholar] [CrossRef]
- Su, H.-M. Mechanisms of n-3 fatty acid-mediated development and maintenance of learning memory performance. J. Nutr. Biochem. 2010, 21, 364–373. [Google Scholar] [CrossRef]
- Chen, C.; Liao, J.; Xia, Y.; Liu, X.; Jones, R.; Haran, J.; McCormick, B.; Sampson, T.R.; Alam, A.; Ye, K. Gut microbiota regulate Alzheimer’s disease pathologies and cognitive disorders via PUFA-associated neuroinflammation. Gut 2022. [Google Scholar] [CrossRef]
- Su, K.-P.; Lai, H.-C.; Yang, H.-T.; Su, W.-P.; Peng, C.-Y.; Chang, J.P.-C.; Chang, H.-C.; Pariante, C.M. Omega-3 Fatty Acids in the Prevention of Interferon-Alpha-Induced Depression: Results from a Randomized, Controlled Trial. Biol. Psychiatry 2014, 76, 559–566. [Google Scholar] [CrossRef]
- Saraswathi, V.; Heineman, R.; Alnouti, Y.; Shivaswamy, V.; DeSouza, C.V. A combination of Omega-3 PUFAs and COX inhibitors: A novel strategy to manage obesity-linked dyslipidemia and adipose tissue inflammation. J. Diabetes its Complicat. 2019, 34, 107494. [Google Scholar] [CrossRef]
- García-Díaz, D.F.; Campion, J.; Milagro, F.I.; Lomba, A.; Marzo, F.; Martínez, J.A. Chronic mild stress induces variations in locomotive behavior and metabolic rates in high fat fed rats. J. Physiol. Biochem. 2007, 63, 337–346. [Google Scholar] [CrossRef]
- Willner, P. The chronic mild stress (CMS) model of depression: History, evaluation and usage. Neurobiol. Stress 2017, 6, 78–93. [Google Scholar] [CrossRef] [Green Version]
- Contreras, C.M.; Chacón, L.; Rodríguez-Landa, J.F.; Bernal-Morales, B.; Gutiérrez-García, A.G.; Saavedra, M. Spontaneous firing rate of lateral septal neurons decreases after forced swimming test in Wistar rat. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2004, 28, 343–348. [Google Scholar] [CrossRef]
- Vorhees, C.V.; Williams, M. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [Green Version]
- Yanar, K.; Simsek, B.; Atukeren, P.; Aydin, S.; Cakatay, U. Is D-Galactose a Useful Agent for Accelerated Aging Model of Gastrocnemius and Soleus Muscle of Sprague-Dawley Rats? Rejuvenation Res. 2019, 22, 521–528. [Google Scholar] [CrossRef]
- Azman, K.F.; Zakaria, R. d-Galactose-induced accelerated aging model: An overview. Biogerontology 2019, 20, 763–782. [Google Scholar] [CrossRef]
- Bruce-Keller, A.J.; Salbaum, J.M.; Luo, M.; Blanchard, E.; Taylor, C.M.; Welsh, D.A.; Berthoud, H.-R. Obese-type Gut Microbiota Induce Neurobehavioral Changes in the Absence of Obesity. Biol. Psychiatry 2014, 77, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Dutheil, S.; Ota, K.T.; Wohleb, E.S.; Rasmussen, K.; Duman, R.S. High-fat diet induced anxiety and anhedonia: Impact on brain homeostasis and inflammation. Neuropsychopharmacology 2016, 47, 1874–1887. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Zhang, Q.; Ma, W.; Tian, F.; Shen, H.; Zhou, M. A combination of quercetin and resveratrol reduces obesity in high-fat diet-fed rats by modulation of gut microbiota. Food Funct. 2017, 8, 4644–4656. [Google Scholar] [CrossRef]
- Jiang, H.; Ling, Z.; Zhang, Y.; Mao, H.; Ma, Z.; Yin, Y.; Wang, W.; Tang, W.; Tan, Z.; Shi, J.; et al. Altered fecal microbiota composition in patients with major depressive disorder. Brain Behav. Immun. 2015, 48, 186–194. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, P.W.; Jeffery, I.B. Gut microbiota and aging. Science 2015, 350, 1214–1215. [Google Scholar] [CrossRef]
- Kumar, M.; Babaei, P.; Ji, B.; Nielsen, J. Human gut microbiota and healthy aging: Recent developments and future prospective. Nutr. Heal. Aging 2016, 4, 3–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.M.; Milillo, M.M.; Krause-Sorio, B.; Siddarth, P.; Kilpatrick, L.; Narr, K.L.; Jacobs, J.P.; Lavretsky, H. Gut Microbiome Diversity and Abundance Correlate with Gray Matter Volume (GMV) in Older Adults with Depression. Int. J. Environ. Res. Public Heal. 2022, 19, 2405. [Google Scholar] [CrossRef]
- Scott, K.A.; Ida, M.; Peterson, V.L.; Prenderville, J.A.; Moloney, G.M.; Izumo, T.; Murphy, K.; Murphy, A.; Ross, R.P.; Stanton, C.; et al. Revisiting Metchnikoff: Age-related alterations in microbiota-gut-brain axis in the mouse. Brain Behav. Immun. 2017, 65, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Parker, B.J.; Wearsch, P.A.; Veloo, A.C.M.; Rodriguez-Palacios, A. The Genus Alistipes: Gut Bacteria with Emerging Implications to Inflammation, Cancer, and Mental Health. Front. Immunol. 2020, 11, 906. [Google Scholar] [CrossRef] [PubMed]
- Naseribafrouei, A.; Hestad, K.; Avershina, E.; Sekelja, M.; Linløkken, A.; Wilson, R.; Rudi, K. Correlation between the human fecal microbiota and depression. Neurogastroenterol. Motil. 2014, 26, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Simpson, C.A.; Diaz-Arteche, C.; Eliby, D.; Schwartz, O.S.; Simmons, J.G.; Cowan, C.S. The gut microbiota in anxiety and depression – A systematic review. Clin. Psychol. Rev. 2020, 83, 101943. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.-S.; Cha, L.; Sim, M.; Jung, S.; Chun, W.Y.; Baik, H.W.; Shin, D.-M. Probiotic Supplementation Improves Cognitive Function and Mood with Changes in Gut Microbiota in Community-Dwelling Older Adults: A Randomized, Double-Blind, Placebo-Controlled, Multicenter Trial. J. Gerontol. Ser. A 2020, 76, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Dong, T.S.; Krause-Sorio, B.; Siddarth, P.; Milillo, M.M.; Lagishetty, V.; Datta, T.; Aguilar-Faustino, Y.; Jacobs, J.P.; Lavretsky, H. The intestinal microbiota as a predictor for antidepressant treatment outcome in geriatric depression: A prospective pilot study. Int. Psychogeriatr. 2021, 34, 33–45. [Google Scholar] [CrossRef]
- Tavella, T.; Rampelli, S.; Guidarelli, G.; Bazzocchi, A.; Gasperini, C.; Pujos-Guillot, E.; Comte, B.; Barone, M.; Biagi, E.; Candela, M.; et al. Elevated gut microbiome abundance of Christensenellaceae, Porphyromonadaceae and Rikenellaceae is associated with reduced visceral adipose tissue and healthier metabolic profile in Italian elderly. Gut Microbes. 2021, 13, 1–19. [Google Scholar] [CrossRef]
- Tung, T.-H.; Nguyen, N.T.K.; Huang, S.-Y. New Insights into Depressive Disorder with Respect to Low-Grade Inflammation and Fish Oil Intake. J. Oleo Sci. 2021, 70, 1539–1550. [Google Scholar] [CrossRef]
- Joffre, C.; Rey, C.; Layé, S. N-3 Polyunsaturated Fatty Acids and the Resolution of Neuroinflammation. Front. Pharmacol. 2019, 10, 1022. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.R.; Lee, H.C.; Lo, Y.C.; Yu, S.C.; Huang, S.Y. n-3 polyunsaturated fatty acids prevent d-galactose-induced cognitive deficits in prediabetic rats. Food Funct. 2018, 9, 2228–2239. [Google Scholar] [CrossRef]
- Su, K.-P.; Huang, S.-Y.; Chiu, C.-C.; Shen, W.W. Omega-3 fatty acids in major depressive disorder: A preliminary double-blind, placebo-controlled trial. Eur. Neuropsychopharmacol. 2003, 13, 267–271. [Google Scholar] [CrossRef]
- Chen, Y.-L.; Shirakawa, H.; Lu, N.-S.; Peng, H.-C.; Xiao, Q.; Yang, S.-C. Impacts of fish oil on the gut microbiota of rats with alcoholic liver damage. J. Nutr. Biochem. 2020, 86, 108491. [Google Scholar] [CrossRef]
- Qin, N.; Song, G.; Ren, X.; Zhang, L.; Gao, J.; Xia, X.; Zhu, B. Fish oil extracted from Coregonus peled improves obese phenotype and changes gut microbiota in a high-fat diet-induced mouse model of recurrent obesity. Food Funct. 2020, 11, 6158–6169. [Google Scholar] [CrossRef]
- Fabersani, E.; Portune, K.; Campillo, I.; López-Almela, I.; la Paz, S.M.-D.; Romaní-Pérez, M.; Benítez-Páez, A.; Sanz, Y. Bacteroides uniformis CECT 7771 alleviates inflammation within the gut-adipose tissue axis involving TLR5 signaling in obese mice. Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef]
- Zhao, Y.; Jaber, V.; Lukiw, W.J. Secretory Products of the Human GI Tract Microbiome and Their Potential Impact on Alzheimer’s Disease (AD): Detection of Lipopolysaccharide (LPS) in AD Hippocampus. Front. Cell. Infect. Microbiol. 2017, 7, 318. [Google Scholar] [CrossRef]
- Nunes, A.F.; Amaral, J.D.; Lo, A.C.; Fonseca, M.B.; Viana, R.J.; Callaerts-Vegh, Z.; D’Hooge, R.; Rodrigues, C.M. TUDCA, a bile acid, attenuates amyloid precursor protein processing and amyloid-β deposition in APP/PS1 mice. Mol. Neurobiol. 2012, 45, 440–454. [Google Scholar] [CrossRef]
- Yanguas-Casás, N.; Barreda-Manso, M.A.; Pérez-Rial, S.; Nieto–Sampedro, M.; Romero-Ramírez, L. TGFβ Contributes to the Anti-inflammatory Effects of Tauroursodeoxycholic Acid on an Animal Model of Acute Neuroinflammation. Mol. Neurobiol. 2016, 54, 6737–6749. [Google Scholar] [CrossRef]
- Huang, B.; Zhao, Q.; Zhou, J.-H.; Xu, G. Enhanced activity and substrate tolerance of 7α-hydroxysteroid dehydrogenase by directed evolution for 7-ketolithocholic acid production. Appl. Microbiol. Biotechnol. 2019, 103, 2665–2674. [Google Scholar] [CrossRef]
- McMillin, M.; Frampton, G.; Quinn, M.; Ashfaq, S.; de los Santos, M., 3rd; Grant, S.; DeMorrow, S. Bile Acid Signaling Is Involved in the Neurological Decline in a Murine Model of Acute Liver Failure. Am. J. Pathol. 2016, 186, 312–323. [Google Scholar] [CrossRef]
- Mortiboys, H.; Furmston, R.; Bronstad, G.; Aasly, J.; Elliott, C.; Bandmann, O. UDCA exerts beneficial effect on mitochondrial dysfunction in LRRK2(G2019S) carriers and in vivo. Neurology 2015, 85, 846–852. [Google Scholar] [CrossRef] [Green Version]
- Ramalho, R.M.; Ribeiro, P.S.; Solá, S.; Castro, R.E.; Steer, C.J.; Rodrigues, C.M. Inhibition of the E2F-1/p53/Bax pathway by tauroursodeoxycholic acid in amyloid beta-peptide-induced apoptosis of PC12 cells. J. Neurochemistry. 2004, 90, 567–575. [Google Scholar] [CrossRef]
- Dionísio, P.; Amaral, J.D.; Ribeiro, M.F.; Lo, A.C.; D’Hooge, R.; Rodrigues, C.M. Amyloid-β pathology is attenuated by tauroursodeoxycholic acid treatment in APP/PS1 mice after disease onset. Neurobiol. Aging 2015, 36, 228–240. [Google Scholar] [CrossRef] [Green Version]
- Yanguas-Casás, N.; Barreda-Manso, M.A.; Nieto-Sampedro, M.; Romero-Ramírez, L. Tauroursodeoxycholic acid reduces glial cell activation in an animal model of acute neuroinflammation. J. Neuroinflammation 2014, 11, 50. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (g/kg Dry Matter) | Group | ||
|---|---|---|---|
| N, ND, CD, PCD | FCD | CCD | |
| Casein | 140.00 | 140.00 | 140.00 |
| L-Cystine | 1.80 | 1.80 | 1.80 |
| Corn starch | 465.69 | 465.69 | 465.69 |
| Dextrin | 155.00 | 155.00 | 155.00 |
| Sucrose | 100.00 | 100.00 | 100.00 |
| Cellulose | 50.00 | 50.00 | 50.00 |
| Soybean oil | 40.00 | 20.00 | 20.00 |
| Corn oil | 0 | 0 | 20.00 |
| LA | 0 | 0 | 11.2 |
| Fish oil | 0 | 20.00 | 0 |
| Eicosapentaenoic acid (EPA) | 0 | 3.60 | 0 |
| Docosahexaenoic acid (DHA) | 0 | 2.40 | 0 |
| AIN-93M Mineral mix | 35.00 | 35.00 | 35.00 |
| AIN-93M Vitamin mix | 10.00 | 10.00 | 10.00 |
| Choline bitartrate | 2.50 | 2.50 | 2.50 |
| t-butylhydroquinone | 0.01 | 0.01 | 0.01 |
| Fatty acid profiles (%) | |||
| C14:0 | 1.42 | 5.63 | 1.23 |
| C16:0 | 13.05 | 18.02 | 10.41 |
| C16:1 (n-7) | 0.07 | 0.04 | 0.03 |
| C18:0 | 4.87 | 7.97 | 6.22 |
| C18:1 (n-9) | 17.79 | 9.95 | 19.01 |
| C18:2 (n-6) | 52.71 | 28.74 | 59.52 |
| C18:3 (n-3) | 9.88 | 6.19 | 3.26 |
| C20:1 (n-9) | N.D. | 0.27 | N.D. |
| C20:2 (n-6) | N.D. | 0.38 | N.D. |
| C20:4 (n-6) | 0.01 | 4.08 | 0.01 |
| C20:5 (n-3) | N.D. | 9.63 | N.D. |
| C22:5 (n-3) | 0.18 | 1.46 | 0.08 |
| C22:6 (n-3) | N.D. | 5.98 | N.D. |
| n-6/n-3 ratio | 5.24 | 1.30 | 17.82 |
| Biochemical Characteristics | Group | |||||
|---|---|---|---|---|---|---|
| N | ND | CD | PCD | FCD | CCD | |
| Body weight | 702 ± 48.3 a | 639 ± 73.8 ab | 583 ± 71.3 bc | 522 ± 62.3 c | 585 ± 79.1 bc | 622 ± 56.2 ab |
| FBG (mg/dL) | 99 ± 6 ns | 105 ± 7 | 109 ± 5 | 103 ± 8 | 96 ± 7 | 102 ± 5 |
| ALB (g/dL) | 4.14 ± 0.07 ns | 4.03 ± 0.10 | 4.00 ± 0.08 | 4.13 ± 0.04 | 4.11 ± 0.06 | 4.02 ± 0.09 |
| TG (mg/dL) | 71.3 ± 2.8 a | 80.7 ± 7.3 a | 69.2 ± 5.9 a | 73.9 ± 3.9 a | 60.2 ± 3.6 b | 78.8 ± 5.5 a |
| TC (mg/dL) | 66.6 ± 3.2 b | 70.4 ± 4.3 b | 81.6 ± 5.7 a | 74.5 ± 6.5 b | 68.8 ± 7.1 b | 72.9 ± 10.5 b |
| LDLc (mg/dL) | 6.89 ± 0.91 a | 7.11 ± 0.58 a | 7.94 ± 1.12 a | 5.82 ± 0.55 ab | 5.52 ± 0.31 b | 6.56 ± 0.72 a |
| HDLc (mg/dL) | 23.1 ± 1.4 ns | 24.5 ± 2.2 | 25.0 ± 1.7 | 24.2 ± 1.5 | 19.8 ± 1.8 | 23.6 ± 1.4 |
| AST (U/L) | 87.2 ± 7.2 ns | 76.9 ± 7.1 | 77.4 ± 6.1 | 81.8 ± 5.2 | 79.6 ± 2.9 | 85.2 ± 5.3 |
| BUN (mg/dL) | 20.8 ± 1.6 a | 19.7 ± 1.3 a | 19.9 ± 0.8 a | 16.1 ± 1.0 b | 15.8 ± 1.3 b | 19.1 ± 0.8 a |
| CRE (mg/dL) | 0.60 ± 0.04 b | 0.66 ± 0.06 ab | 0.59 ± 0.05 b | 0.72 ± 0.06 a | 0.68 ± 0.10 a | 0.69 ± 0.06 a |
| TNF-α (pg/mL) | 41.9 ± 3.1 b | 56.3 ± 6.2 a | 69.3 ± 3.7 a | 40.7 ± 5.1 b | 44.3 ± 2.6 b | 61.7 ± 4.2 a |
| IL-1β (pg/mL) | 20.2 ± 2.4 b | 48.6 ± 8.2 a | 49.4 ± 4.3 a | 32.3 ± 2.7 b | 24.1 ± 5.2 b | 43.4 ± 2.5 a |
| IL-6 (pg/mL) | 40.2 ± 1.9 ns | 41.8 ± 2.6 | 41.0 ± 2.1 | 39.4 ± 2.9 | 36.5 ± 2.8 | 38.2 ± 2.3 |
| Corticosterone (ng/mL) | 135 ± 14 b | 265 ± 27 a | 205 ± 19 a | 187 ± 11 a | 161 ± 13 b | 200 ± 16 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tung, T.-H.; Chen, Y.-C.; Lin, Y.-T.; Huang, S.-Y. N-3 PUFA Ameliorates the Gut Microbiota, Bile Acid Profiles, and Neuropsychiatric Behaviours in a Rat Model of Geriatric Depression. Biomedicines 2022, 10, 1594. https://doi.org/10.3390/biomedicines10071594
Tung T-H, Chen Y-C, Lin Y-T, Huang S-Y. N-3 PUFA Ameliorates the Gut Microbiota, Bile Acid Profiles, and Neuropsychiatric Behaviours in a Rat Model of Geriatric Depression. Biomedicines. 2022; 10(7):1594. https://doi.org/10.3390/biomedicines10071594
Chicago/Turabian StyleTung, Te-Hsuan, Yang-Ching Chen, Ya-Tin Lin, and Shih-Yi Huang. 2022. "N-3 PUFA Ameliorates the Gut Microbiota, Bile Acid Profiles, and Neuropsychiatric Behaviours in a Rat Model of Geriatric Depression" Biomedicines 10, no. 7: 1594. https://doi.org/10.3390/biomedicines10071594
APA StyleTung, T. -H., Chen, Y. -C., Lin, Y. -T., & Huang, S. -Y. (2022). N-3 PUFA Ameliorates the Gut Microbiota, Bile Acid Profiles, and Neuropsychiatric Behaviours in a Rat Model of Geriatric Depression. Biomedicines, 10(7), 1594. https://doi.org/10.3390/biomedicines10071594