
NO-Dependent Mechanisms of p53 Expression and Cell Death in Rat’s Dorsal Root Ganglia after Sciatic-Nerve Transection

 , , , , and
, , , , and
Abstract
:
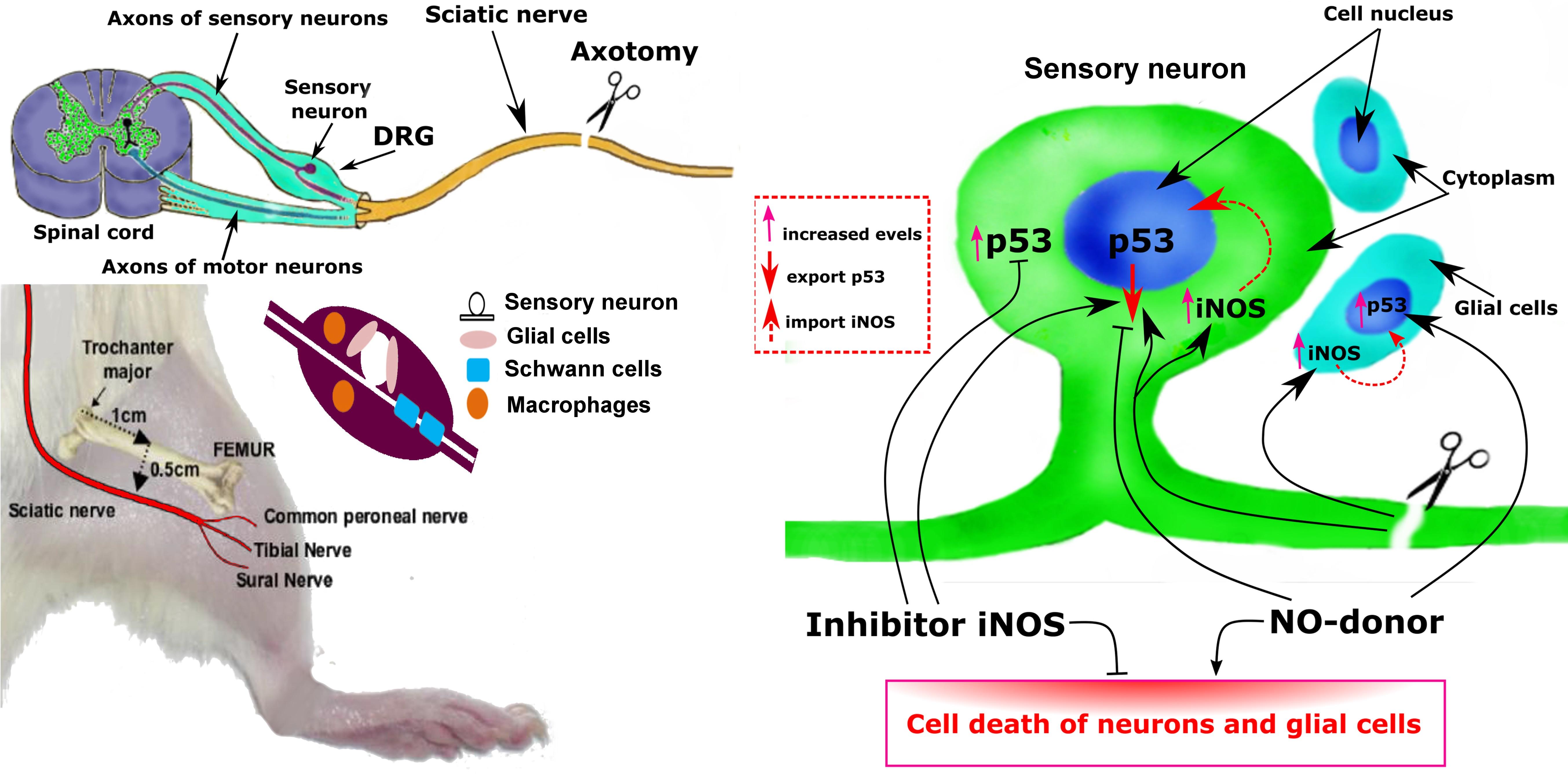{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals and Ethical Approval
2.2. Objects and Procedure of Axotomy
2.3. Immunofluorescent Microscopy
2.4. Western Blot
2.5. The Analysis of Cell Death
2.6. The Statistical Analysis
3. Results
3.1. Nuclear-Cytoplasmic Redistribution of p53 and NO-Dependent Deposition of p53 in Neurons and Glial Cells after Axotomy
3.2. Upregulation of iNOS in the Nuclear and Cytoplasmic Fractions of DRGs at Different Time Intervals after Sciatic-Nerve Transection
3.3. The NO-Dependent Activation of Neuron and Glial Cell Death by NO Donors and Cytoprotector Effect of the Selective Inhibitor of iNOS after Axotomy
4. Discussion
5. Conclusions and Future Decisions
- Axotomy induces nuclear–cytoplasmic redistribution of p53 in DRG neurons, 24 h after trauma.
- The NO donor (SNP) induces nuclear deposition of p53 in neurons and glial cells of DRGs, 4 h and 24 h after axotomy.
- The selective inhibitor of inducible NO-synthase (SMT) induces decrease in the level of p53 in DRG neurons, 4 h and 24 h after axotomy.
- Axotomy induces increase in the level of iNOS in both nucleus and cytoplasm of DRG cells, 4 h, 24 h, and 7 days after trauma.
- Axotomy increases the level of neural and glial-cell death, 24 h and 7 days after trauma.
- The NO donor SNP increases neural and glial-cell death, 24 h and 7 days after axotomy.
- The selective inhibitor of inducible NO-synthase SMT decreases neural and glial-cell death, 24 h and 7 days after trauma.
- Axotomy increases the Bax–Bcl-2 index in DRG cells, 24 h and 7 days after trauma.
- The NO donor (SNP) significantly increases the Bax–Bcl-2 index in DRG cells, 24 h and 7 days after axotomy.
- The selective inhibitor of inducible NO-synthase (SMT) decreases the Bax–Bcl-2 index in DRG cells, 24 h and 7 days after axotomy.
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Lepard, J.; Ammar, A.; Shlobin, N.A.; Boyke, A.E.; Berger, C.; Vital, A.; Rolle, M.; Corley, J.; Barthélemy, E.J.; Park, K.B. An Assessment of Global Neurotrauma Prevention and Care Delivery: The Provider Perspective. World Neurosurg. 2021, 156, e183–e191. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.S.; Coleman, M.P.; Menon, D.K. Traumatic Axonal Injury: Mechanisms and Translational Opportunities. Trends Neurosci. 2016, 39, 311–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laskowitz, D.; Grant, G.E. Translational Research in Traumatic Brain Injury; CRC Press/Taylor and Francis Group: Boca Raton, FL, USA, 2016. [Google Scholar]
- Khaitin, A.; Rudkovskii, M.; Uzdensky, A. Ca2+ mediates axotomy-induced necrosis and apoptosis of satellite glial cells remote from the transection site in the isolated crayfish mechanoreceptor. Mol. Cell. Neurosci. 2018, 88, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Cooke, R.M.; Mistry, R.; Challiss, R.A.J.; Straub, V.A. Nitric Oxide Synthesis and cGMP Production Is Important for Neurite Growth and Synapse Remodeling after Axotomy. J. Neurosci. 2013, 33, 5626–5637. [Google Scholar] [CrossRef] [Green Version]
- Koeberle, P.D.; Ball, A.K. Nitric Oxide Synthase Inhibition Delays Axonal Degeneration and Promotes the Survival of Axotomized Retinal Ganglion Cells. Exp. Neurol. 1999, 158, 366–381. [Google Scholar] [CrossRef]
- Lehmann, H.C.; Köhne, A.; Meyer zu Hörste, G.; Dehmel, T.; Kiehl, O.; Hartung, H.-P.; Kastenbauer, S.; Kieseier, B.C. Role of Nitric Oxide as Mediator of Nerve Injury in Inflammatory Neuropathies. J. Neuropathol. Exp. Neurol. 2007, 66, 305–312. [Google Scholar] [CrossRef]
- Wilkins, A.; Compston, A. Trophic factors attenuate nitric oxide mediated neuronal and axonal injury in vitro: Roles and interactions of mitogen-activated protein kinase signalling pathways. J. Neurochem. 2005, 92, 1487–1496. [Google Scholar] [CrossRef]
- González-Forero, D.; Moreno-López, B. Retrograde response in axotomized motoneurons: Nitric oxide as a key player in triggering reversion toward a dedifferentiated phenotype. Neuroscience 2014, 283, 138–165. [Google Scholar] [CrossRef] [Green Version]
- Thippeswamy, T.; Haddley, K.; Corness, J.D.; Howard, M.R.; McKay, J.S.; Beaucourt, S.M.; Pope, M.D.; Murphy, D.; Morris, R.; Hökfelt, T.; et al. NO-cGMP mediated galanin expression in NGF-deprived or axotomized sensory neurons. J. Neurochem. 2007, 100, 790–801. [Google Scholar] [CrossRef]
- Chu, T.-H.; Wu, W.-T. Nitric oxide synthase inhibitor attenuates number of regenerating spinal motoneurons in adult rats. Neuroreport 2006, 17, 969–973. [Google Scholar] [CrossRef]
- Forstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero, P.; Laforest, R.; Shoghi, K.; Zhou, D.; Ewald, G.; Pfeifer, J.; Duncavage, E.; Krupp, K.; Mach, R.; Gropler, R. Feasibility and Dosimetry Studies for 18 F-NOS as a Potential PET Radiopharmaceutical for Inducible Nitric Oxide Synthase in Humans. J. Nucl. Med. 2012, 53, 994–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiallos-Estrada, C.E.; Kummer, W.; Mayer, B.; Bravo, R.; Zimmermann, M.; Herdegen, T. Long-lasting increase of nitric oxide synthase immunoreactivity, NADPH-diaphorase reaction and c-JUN co-expression in rat dorsal root ganglion neurons following sciatic nerve transection. Neurosci. Lett. 1993, 150, 169–173. [Google Scholar] [CrossRef]
- Garthwaite, J. NO as a multimodal transmitter in the brain: Discovery and current status. Br. J. Pharmacol. 2019, 176, 197–211. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.K.; Yoon, C.S.; Kim, S.G.; Park, Y.W.; Lee, S.S.; Lee, S.K. Effects of 4-Hexylresorcinol on Protein Expressions in RAW 264.7 Cells as Determined by Immunoprecipitation High Performance Liquid Chromatography. Sci. Rep. 2019, 9, 3379. [Google Scholar] [CrossRef]
- Nakaya, N.; Lowe, S.W.; Taya, Y.; Chenchik, A.; Enikolopov, G. Specific pattern of p53 phosphorylation during nitric oxide-induced cell cycle arrest. Oncogene 2000, 19, 6369–6375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zalcenstein, A.; Oren, M. Nitric oxide promotes p53 nuclear retention and sensitizes neuroblastoma cells to apoptosis by ionizing radiation. Cell Death Differ. 2003, 10, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Aubrey, B.J.; Kelly, G.L.; Janic, A.; Herold, M.J.; Strasser, A. How does p53 induce apoptosis and how does this relate to p53-mediated tumour suppression? Cell Death Differ. 2018, 25, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Beyfuss, K.; Hood, D.A. A systematic review of p53 regulation of oxidative stress in skeletal muscle. Redox Rep. 2018, 23, 100–117. [Google Scholar] [CrossRef] [Green Version]
- Simabuco, F.M.; Morale, M.G.; Pavan, I.C.B.; Morelli, A.P.; Silva, F.R.; Tamura, R.E. p53 and metabolism: From mechanism to therapeutics. Oncotarget 2018, 9, 23780–23823. [Google Scholar] [CrossRef] [Green Version]
- Checler, F.; Alves da Costa, C. p53 in neurodegenerative diseases and brain cancers. Pharmacol. Ther. 2014, 142, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Jebelli, J.D.; Hooper, C.; Garden, G.A.; Pocock, J.M. Emerging roles of p53 in glial cell function in health and disease. Glia 2012, 60, 515–525. [Google Scholar] [CrossRef] [Green Version]
- Kiryu-Seo, S. Noxa Is a Critical Mediator of p53-Dependent Motor Neuron Death after Nerve Injury in Adult Mouse. J. Neurosci. 2005, 25, 1442–1447. [Google Scholar] [CrossRef] [PubMed]
- Culmsee, C.; Mattson, M.P. p53 in neuronal apoptosis. Biochem. Biophys. Res. Commun. 2005, 331, 761–777. [Google Scholar] [CrossRef]
- Joshi, Y.; Sória, M.G.; Quadrato, G.; Inak, G.; Zhou, L.; Hervera, A.; Rathore, K.I.; Elnaggar, M.; Cucchiarini, M.; Marine, J.C.; et al. The MDM4/MDM2-p53-IGF1 axis controls axonal regeneration, sprouting and functional recovery after CNS injury. Brain 2015, 138, 1843–1862. [Google Scholar] [CrossRef] [PubMed]
- Di Giovanni, S.; Knights, C.D.; Rao, M.; Yakovlev, A.; Beers, J.; Catania, J.; Avantaggiati, M.L.; Faden, A.I. The tumor suppressor protein p53 is required for neurite outgrowth and axon regeneration. EMBO J. 2006, 25, 4084–4096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodkin, S.; Khaitin, A.; Pitinova, M.; Dzreyan, V.; Guzenko, V.; Rudkovskii, M.; Sharifulina, S.; Uzdensky, A. The Localization of p53 in the Crayfish Mechanoreceptor Neurons and Its Role in Axotomy-Induced Death of Satellite Glial Cells Remote from the Axon Transection Site. J. Mol. Neurosci. 2020, 70, 532–541. [Google Scholar] [CrossRef]
- Dzreyan, V.; Rodkin, S.; Nikul, V.; Pitinova, M.; Uzdensky, A. The Expression of E2F1, p53, and Caspase 3 in the Rat Dorsal Root Ganglia After Sciatic Nerve Transection. J. Mol. Neurosci. 2021, 71, 826–835. [Google Scholar] [CrossRef]
- Savastano, L.E.; Laurito, S.R.; Fitt, M.R.; Rasmussen, J.A.; Gonzalez Polo, V.; Patterson, S.I. Sciatic nerve injury: A simple and subtle model for investigating many aspects of nervous system damage and recovery. J. Neurosci. Methods 2014, 227, 166–180. [Google Scholar] [CrossRef]
- Li, J.; Zhang, W.; Zhu, S.; Shi, F. Nitric Oxide Synthase Is Involved in Follicular Development via the PI3K/AKT/FoxO3a Pathway in Neonatal and Immature Rats. Animals 2020, 10, 248. [Google Scholar] [CrossRef] [Green Version]
- Naik, A.K.; Tandan, S.K.; Kumar, D.; Dudhgaonkar, S.P. Nitric oxide and its modulators in chronic constriction injury-induced neuropathic pain in rats. Eur. J. Pharmacol. 2006, 530, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Crowley, L.C.; Marfell, B.J.; Waterhouse, N.J. Analyzing Cell Death by Nuclear Staining with Hoechst 33342. Cold Spring Harb. Protoc. 2016, 2016, pdb.prot087205. [Google Scholar] [CrossRef] [PubMed]
- Bolte, S.; Cordelieres, F.P. A guided tour into subcellular colocalization analysis in light microscopy. J. Microsc. 2006, 224, 213–232. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, P.A.; Scriven, D.R.L.; Schulson, M.N.; Moore, E.D.W. Multi-Image Colocalization and Its Statistical Significance. Biophys. J. 2010, 99, 1996–2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandau, K.; Pfeilschifter, J.; Brüne, B. Nitric oxide and superoxide induced p53 and Bax accumulation during mesangial cell apoptosis. Kidney Int. 1997, 52, 378–386. [Google Scholar] [CrossRef] [Green Version]
- Schneiderhan, N.; Budde, A.; Zhang, Y.; Brüne, B. Nitric oxide induces phosphorylation of p53 and impairs nuclear export. Oncogene 2003, 22, 2857–2868. [Google Scholar] [CrossRef] [Green Version]
- Poluha, W.; Schonhoff, C.M.; Harrington, K.S.; Lachyankar, M.B.; Crosbie, N.E.; Bulseco, D.A.; Ross, A.H. A Novel, Nerve Growth Factor-activated Pathway Involving Nitric Oxide, p53, and p21WAF1 Regulates Neuronal Differentiation of PC12 Cells. J. Biol. Chem. 1997, 272, 24002–24007. [Google Scholar] [CrossRef] [Green Version]
- Martin, L.J. Adult Motor Neuron Apoptosis Is Mediated by Nitric Oxide and Fas Death Receptor Linked by DNA Damage and p53 Activation. J. Neurosci. 2005, 25, 6449–6459. [Google Scholar] [CrossRef] [Green Version]
- Khodapasand, E.; Jafarzadeh, N.; Farrokhi, F.; Kamalidehghan, B.; Houshmand, M. Is Bax/Bcl-2 ratio considered as a prognostic marker with age and tumor location in colorectal cancer? Iran. Biomed. J. 2015, 19, 69–75. [Google Scholar] [CrossRef]
- Fletcher, J.I.; Meusburger, S.; Hawkins, C.J.; Riglar, D.T.; Lee, E.F.; Fairlie, W.D.; Huang, D.C.S.; Adams, J.M. Apoptosis is triggered when prosurvival Bcl-2 proteins cannot restrain Bax. Proc. Natl. Acad. Sci. USA 2008, 105, 18081–18087. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Zhang, J.; Ma, P.; Myers, D.E.; Goldberg, I.G.; Sittler, K.J.; Barb, J.J.; Munson, P.J.; del Pilar Cintron, A.; McCoy, J.P.; et al. cGMP-independent nitric oxide signaling and regulation of the cell cycle. BMC Genom. 2005, 6, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelmann, D.; Pützer, B.M. Translating DNA damage into cancer cell death—A roadmap for E2F1 apoptotic signalling and opportunities for new drug combinations to overcome chemoresistance. Drug Resist. Updates 2010, 13, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Meng, P.; Ghosh, R. Transcription addiction: Can we garner the Yin and Yang functions of E2F1 for cancer therapy? Cell Death Dis. 2014, 5, e1360. [Google Scholar] [CrossRef] [Green Version]
- Pandareesh, M.D.; Anand, T. Neuroprotective and Anti-Apoptotic Propensity of Bacopa monniera Extract Against Sodium Nitroprusside Induced Activation of iNOS, Heat Shock Proteins and Apoptotic Markers in PC12 Cells. Neurochem. Res. 2014, 39, 800–814. [Google Scholar] [CrossRef]
- Yakovlev, V.A.; Bayden, A.S.; Graves, P.R.; Kellogg, G.E.; Mikkelsen, R.B. Nitration of the Tumor Suppressor Protein p53 at Tyrosine 327 Promotes p53 Oligomerization and Activation. Biochemistry 2010, 49, 5331–5339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.K.; Awasthi, D.; Dubey, M.; Nagarkoti, S.; Kumar, A.; Chandra, T.; Barthwal, M.K.; Tripathi, A.K.; Dikshit, M. High oxidative stress adversely affects NFκB mediated induction of inducible nitric oxide synthase in human neutrophils: Implications in chronic myeloid leukemia. Nitric Oxide 2016, 58, 28–41. [Google Scholar] [CrossRef]
- Villanueva, C.; Giulivi, C. Subcellular and cellular locations of nitric oxide synthase isoforms as determinants of health and disease. Free Radic. Biol. Med. 2010, 49, 307–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordano, A.; Tonello, C.; Bulbarelli, A.; Cozzi, V.; Cinti, S.; Carruba, M.O.; Nisoli, E. Evidence for a functional nitric oxide synthase system in brown adipocyte nucleus. FEBS Lett. 2002, 514, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Provost, C.; Choufani, F.; Avedanian, L.; Bkaily, G.; Gobeil, F.; Jacques, D. Nitric oxide and reactive oxygen species in the nucleus revisitedThis review is one of a selection of papers published in a Special Issue on Oxidative Stress in Health and Disease. Can. J. Physiol. Pharmacol. 2010, 88, 296–304. [Google Scholar] [CrossRef]
- Sierra, A.; Navascués, J.; Cuadros, M.A.; Calvente, R.; Martín-Oliva, D.; Ferrer-Martín, R.M.; Martín-Estebané, M.; Carrasco, M.-C.; Marín-Teva, J.L. Expression of Inducible Nitric Oxide Synthase (iNOS) in Microglia of the Developing Quail Retina. PLoS ONE 2014, 9, e106048. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Hou, C.; Chen, D. Altered Expression of Inducible Nitric Oxide Synthase After Sciatic Nerve Injury in Rat. Cell Biochem. Biophys. 2011, 61, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Faden, A.I.; Yakovlev, A.G. Caspase-Dependent Apoptotic Pathways in CNS Injury. Mol. Neurobiol. 2001, 24, 131–144. [Google Scholar] [CrossRef]
- Wan, C.; Ma, X.; Shi, S.; Zhao, J.; Nie, X.; Han, J.; Xiao, J.; Wang, X.; Jiang, S.; Jiang, J. Pivotal roles of p53 transcription-dependent and -independent pathways in manganese-induced mitochondrial dysfunction and neuronal apoptosis. Toxicol. Appl. Pharmacol. 2014, 281, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Gaub, P.; Tedeschi, A.; Puttagunta, R.; Nguyen, T.; Schmandke, A.; Di Giovanni, S. HDAC inhibition promotes neuronal outgrowth and counteracts growth cone collapse through CBP/p300 and P/CAF-dependent p53 acetylation. Cell Death Differ. 2010, 17, 1392–1408. [Google Scholar] [CrossRef] [Green Version]
- Tedeschi, A.; Nguyen, T.; Puttagunta, R.; Gaub, P.; Di Giovanni, S. A p53-CBP/p300 transcription module is required for GAP-43 expression, axon outgrowth, and regeneration. Cell Death Differ. 2009, 16, 543–554. [Google Scholar] [CrossRef]
- Qin, Q.; Baudry, M.; Liao, G.; Noniyev, A.; Galeano, J.; Bi, X. A Novel Function for p53: Regulation of Growth Cone Motility through Interaction with Rho Kinase. J. Neurosci. 2009, 29, 5183–5192. [Google Scholar] [CrossRef] [Green Version]
- Keilhoff, G.; Fansa, H.; Wolf, G. Neuronal nitric oxide synthase is the dominant nitric oxide supplier for the survival of dorsal root ganglia after peripheral nerve axotomy. J. Chem. Neuroanat. 2002, 24, 181–187. [Google Scholar] [CrossRef]
- Keilhoff, G.; Fansa, H.; Wolf, G. Neuronal NOS deficiency promotes apoptotic cell death of spinal cord neurons after peripheral nerve transection. Nitric Oxide 2004, 10, 101–111. [Google Scholar] [CrossRef]
- Novikov, L.; Novikova, L.; Kellerth, J.-O. Brain-derived neurotrophic factor promotes survival and blocks nitric oxide synthase expression in adult rat spinal motoneurons after ventral root avulsion. Neurosci. Lett. 1995, 200, 45–48. [Google Scholar] [CrossRef]
- Wu, W.; Li, L.; Yick, L.-W.; Chai, H.; Xie, Y.; Yang, Y.; Prevette, D.M.; Oppenheim, R.W. GDNF and BDNF Alter the Expression of Neuronal NOS, c-Jun, and p75 and Prevent Motoneuron Death following Spinal Root Avulsion in Adult Rats. J. Neurotrauma 2003, 20, 603–612. [Google Scholar] [CrossRef]
- Zilundu, P.L.M.; Xu, X.; Liaquat, Z.; Wang, Y.; Zhong, K.; Fu, R.; Zhou, L. Long-Term Suppression of c-Jun and nNOS Preserves Ultrastructural Features of Lower Motor Neurons and Forelimb Function after Brachial Plexus Roots Avulsion. Cells 2021, 10, 1614. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.C.; Neher, J.J. Inflammatory Neurodegeneration and Mechanisms of Microglial Killing of Neurons. Mol. Neurobiol. 2010, 41, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Dzreyan, V.A.; Rodkin, S.V.; Pitinova, M.A.; Uzdensky, A.B. HDAC1 Expression, Histone Deacetylation, and Protective Role of Sodium Valproate in the Rat Dorsal Root Ganglia After Sciatic Nerve Transection. Mol. Neurobiol. 2021, 58, 217–228. [Google Scholar] [CrossRef] [PubMed]
- McKay Hart, A.; Brannstrom, T.; Wiberg, M.; Terenghi, G. Primary sensory neurons and satellite cells after peripheral axotomy in the adult rat. Exp. Brain Res. 2002, 142, 308–318. [Google Scholar] [CrossRef]
- Emirandetti, A.; Simões, G.F.; Zanon, R.G.; Oliveira, A.L.R. Spinal motoneuron synaptic plasticity after axotomy in the absence of inducible nitric oxide synthase. J. Neuroinflamm. 2010, 7, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, H.; Wang, Y.; Chao, X.; Sun, G.; Bai, X.; Xu, L.; Han, Y.; Li, J.; Wang, H.; Fan, Z. Artemin transiently increases iNOS expression in primary cultured trigeminal ganglion neurons. Neurosci. Lett. 2017, 660, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Caggiano, A.O.; Kraig, R.P. Neuronal Nitric Oxide Synthase Expression is Induced in Neocortical Astrocytes after Spreading Depression. J. Cereb. Blood Flow Metab. 1998, 18, 75–87. [Google Scholar] [CrossRef] [Green Version]
- Rapozzi, V.; Della Pietra, E.; Zorzet, S.; Zacchigna, M.; Bonavida, B.; Xodo, L.E. Nitric oxide-mediated activity in anti-cancer photodynamic therapy. Nitric Oxide 2013, 30, 26–35. [Google Scholar] [CrossRef]
- Rapozzi, V.; Della Pietra, E.; Bonavida, B. Dual roles of nitric oxide in the regulation of tumor cell response and resistance to photodynamic therapy. Redox Biol. 2015, 6, 311–317. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Cao, Y.; Liu, Y.; Ping, F.; Shang, J.; Zhang, Z.; Li, Y. Activating Mas receptor protects human pulmonary microvascular endothelial cells against LPS-induced apoptosis via the NF-kB p65/P53 feedback pathways. J. Cell. Physiol. 2019, 234, 12865–12875. [Google Scholar] [CrossRef]
- Kleinert, H.; Schwarz, P.M.; Förstermann, U. Regulation of the Expression of Inducible Nitric Oxide Synthase. Biol. Chem. 2003, 384, 1343–1364. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodkin, S.; Dzreyan, V.; Bibov, M.; Ermakov, A.; Derezina, T.; Kirichenko, E. NO-Dependent Mechanisms of p53 Expression and Cell Death in Rat’s Dorsal Root Ganglia after Sciatic-Nerve Transection. Biomedicines 2022, 10, 1664. https://doi.org/10.3390/biomedicines10071664
Rodkin S, Dzreyan V, Bibov M, Ermakov A, Derezina T, Kirichenko E. NO-Dependent Mechanisms of p53 Expression and Cell Death in Rat’s Dorsal Root Ganglia after Sciatic-Nerve Transection. Biomedicines. 2022; 10(7):1664. https://doi.org/10.3390/biomedicines10071664
Chicago/Turabian StyleRodkin, Stanislav, Valentina Dzreyan, Mikhail Bibov, Alexey Ermakov, Tatyana Derezina, and Evgeniya Kirichenko. 2022. "NO-Dependent Mechanisms of p53 Expression and Cell Death in Rat’s Dorsal Root Ganglia after Sciatic-Nerve Transection" Biomedicines 10, no. 7: 1664. https://doi.org/10.3390/biomedicines10071664
APA StyleRodkin, S., Dzreyan, V., Bibov, M., Ermakov, A., Derezina, T., & Kirichenko, E. (2022). NO-Dependent Mechanisms of p53 Expression and Cell Death in Rat’s Dorsal Root Ganglia after Sciatic-Nerve Transection. Biomedicines, 10(7), 1664. https://doi.org/10.3390/biomedicines10071664