
Beneficial Effects of Empagliflozin Are Mediated by Reduced Renal Inflammation and Oxidative Stress in Spontaneously Hypertensive Rats Expressing Human C-Reactive Protein
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Metabolic Parameters in Epididymal Adipose Tissue and Myocardium
2.3. Tissue Triglycerides and Cholesterol Measurements
2.4. Biochemical Analyses
2.5. Parameters of Oxidative Stress
2.6. Urine Collection and Microalbuminuria
2.7. Cardiac Function and Blood Pressure Measurements
2.8. Quantitative PCR (qPCR)
2.9. Histology
2.10. Statistical Analysis
3. Results
3.1. Effects of Empagliflozin on Body Weight, Weights of Fat Depots, Cardiac Function and Blood Pressure
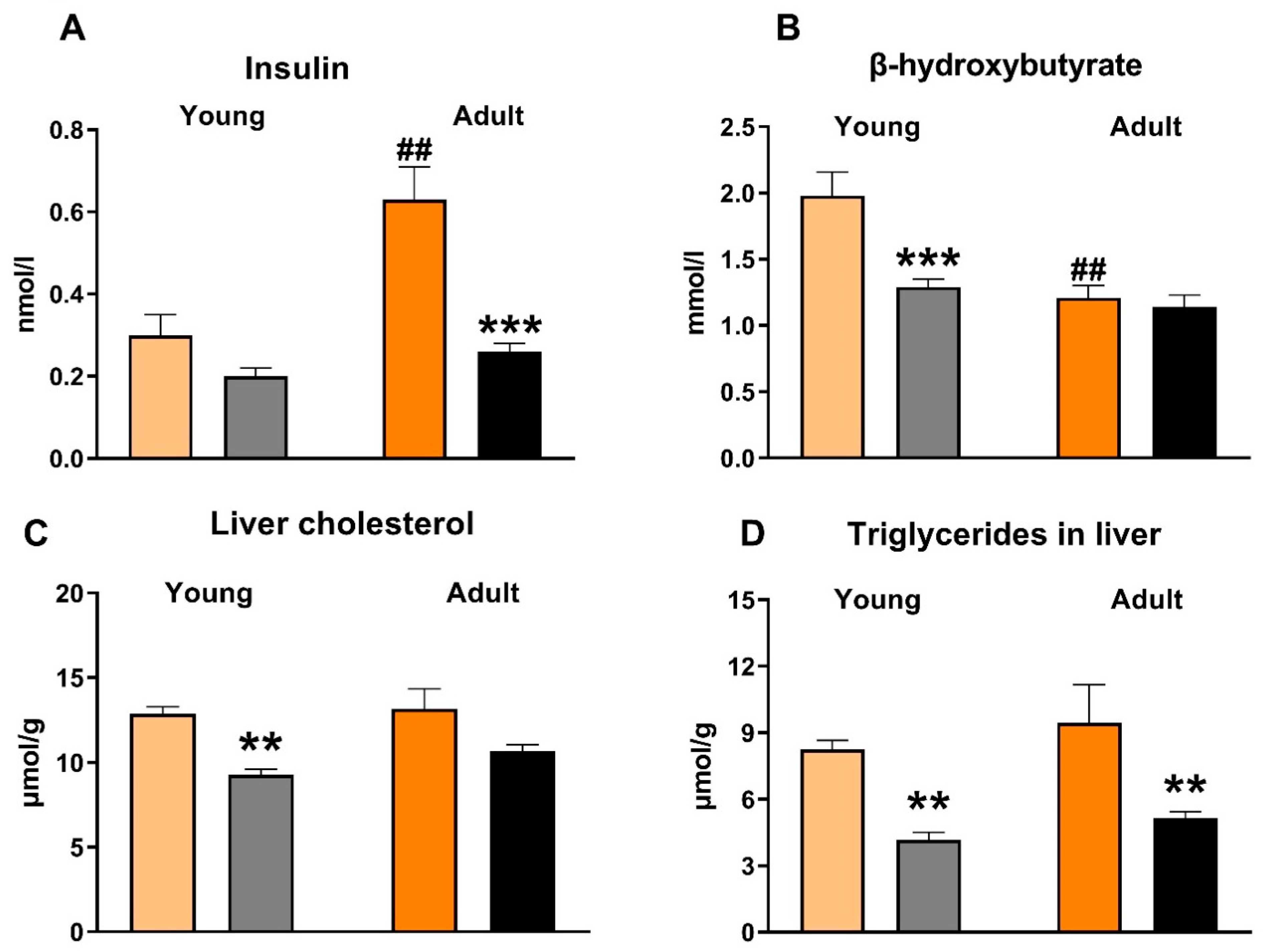
3.2. Effects of Empagliflozin on Metabolic Parameters and Insulin Sensitivity
3.3. Effects of Empagliflozin on Inflammatory and Oxidative Stress Parameters
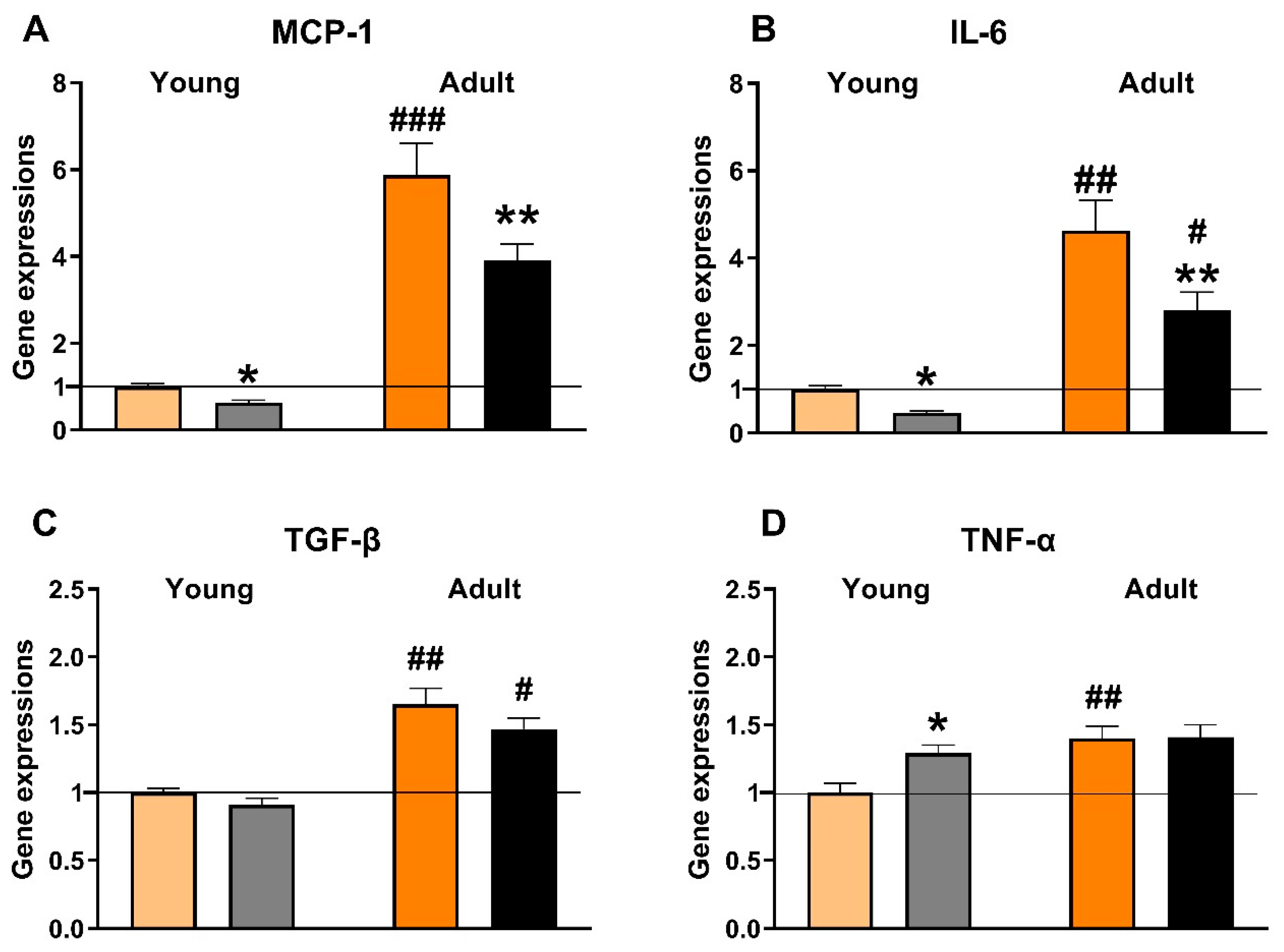
3.4. Effects of Empagliflozin on Renal Expression of Selected Pro-Inflammatory Genes
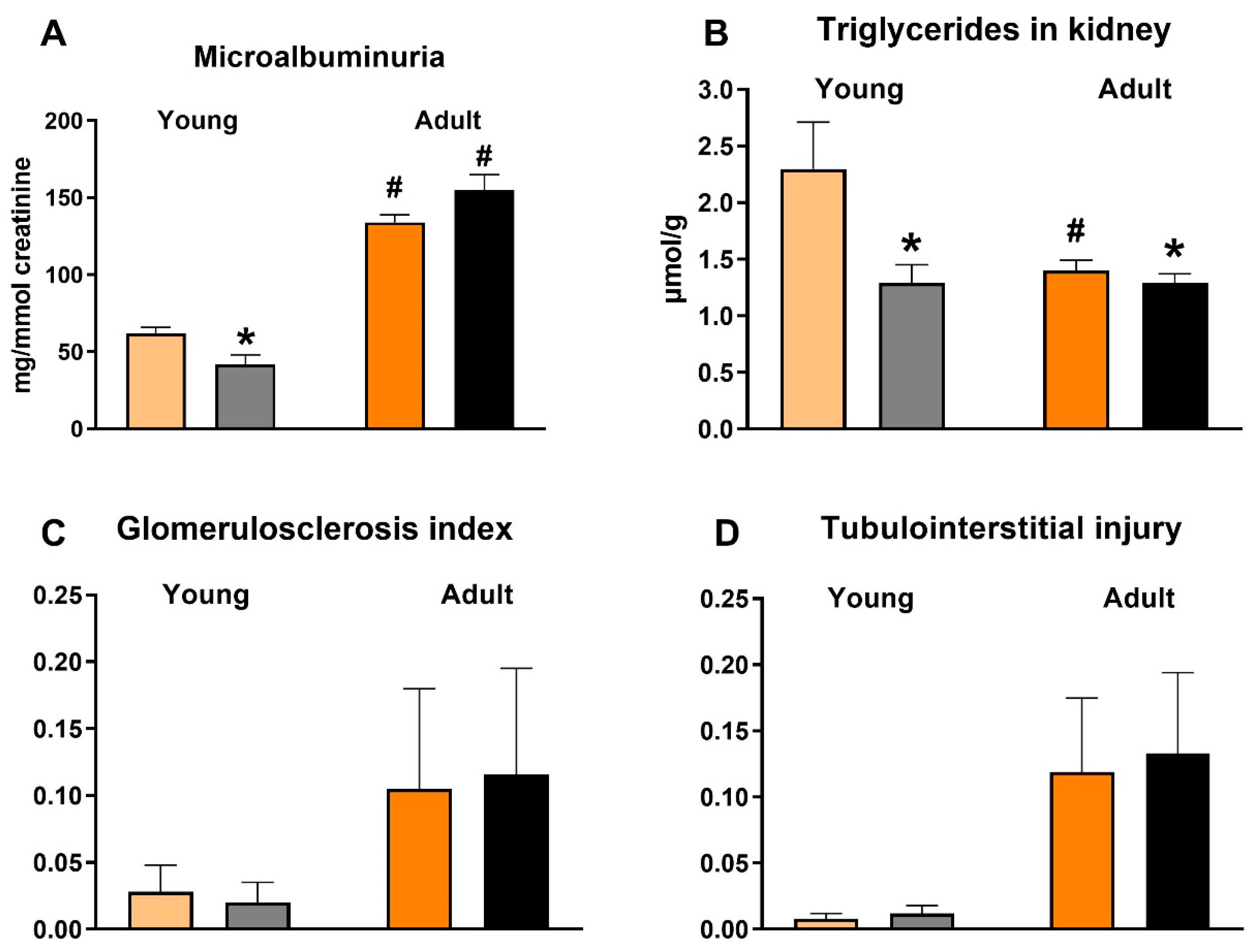
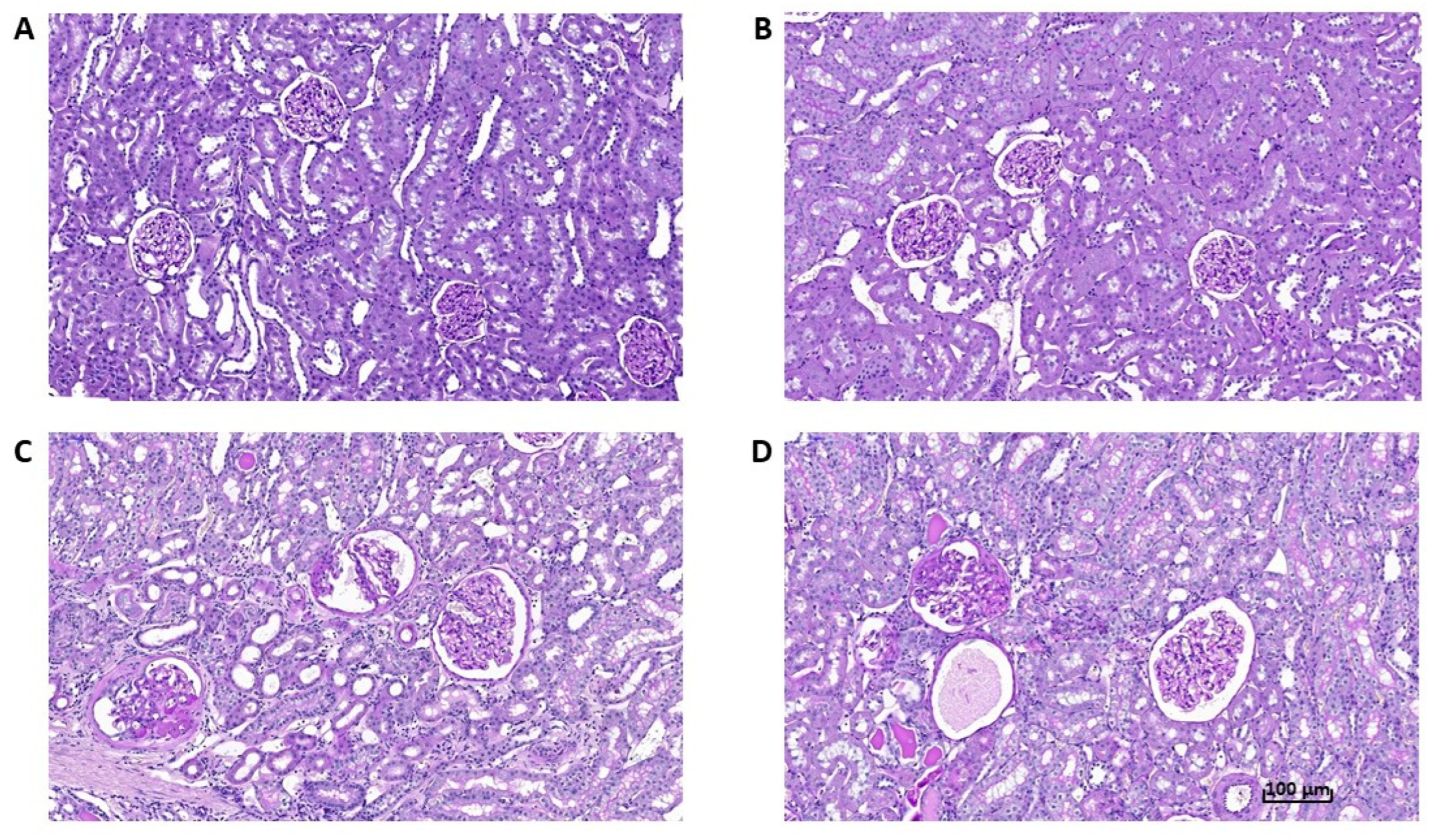
3.5. Effects of Empagliflozin on Renal Function Markers and Histological Analysis
4. Discussion
4.1. Renoprotective Effects of Empagliflozin Are Associated with Reduced Ectopic Fat Accumulation and Lower Inflammation and Oxidative Stress
4.2. Effects of Empagliflozin on Insulin, β-Hydroxybutyrate, NEFA and Leptin Concentrations
4.3. Effects of Empagliflozin on Liver Triglycerides and Cholesterol
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aldossari, K.K. Cardiovascular outcomes and safety with antidiabetic drugs. Int. J. Health Sci. 2018, 12, 70–83. [Google Scholar]
- Hediger, M.A.; Rhoads, D.B. Molecular physiology of sodium glucose cotransporters. Physiol. Rev. 1994, 74, 993–1026. [Google Scholar] [CrossRef] [PubMed]
- Prattichizzo, F.; De Nigris, V.; Spiga, R.; Mancuso, E.; La Sala, L.; Antonicelli, R.; Testa, R.; Procopio, A.D.; Olivieri, F. Inflammageing and metaflammation: The yin and yang of type 2 diabetes. Ageing Res. Rev. 2018, 41, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zinman, B.; Wanner, C.; Lachin, J.M.; Fitchett, D.; Bluhmki, E.; Hantel, S.; Mattheus, M.; Devins, T.; Johansen, O.E.; Woerle, H.J.; et al. Empagliflozin, Cardiovascular Outcomes, and Mortality in Type 2 Diabetes. N. Engl. J. Med. 2015, 373, 2117–2128. [Google Scholar] [CrossRef]
- Wanner, C.; Inzucchi, S.E.; Lachin, J.M.; Fitchett, D.; von Eynatten, M.; Mattheus, M.; Johansen, O.E.; Woerle, H.J.; Broedl, U.C.; Zinman, B.; et al. Empagliflozin and Progression of Kidney Disease in Type 2 Diabetes. N. Engl. J. Med. 2016, 375, 323–334. [Google Scholar] [CrossRef]
- Perkovic, V.; Jardine, M.J.; Neal, B.; Bompoint, S.; Heerspink, H.J.L.; Charytan, D.M.; Edwards, R.; Agarwal, R.; Bakris, G.; Bull, S.; et al. Canagliflozin and Renal Outcomes in Type 2 Diabetes and Nephropathy. N. Engl. J. Med. 2019, 380, 2295–2306. [Google Scholar] [CrossRef]
- Cannon, C.P.; Perkovic, V.; Agarwal, R.; Baldassarre, J.; Bakris, G.; Charytan, D.M.; de Zeeuw, D.; Edwards, R.; Greene, T.; Heerspink, H.J.L.; et al. Evaluating the Effects of Canagliflozin on Cardiovascular and Renal Events in Patients with Type 2 Diabetes Mellitus and Chronic Kidney Disease According to Baseline HbA1c, Including Those with HbA1c <7%: Results from the CREDENCE Trial. Circulation 2020, 141, 407–410. [Google Scholar] [CrossRef]
- Heerspink, H.J.L.; Jongs, N.; Chertow, G.M.; Langkilde, A.M.; McMurray, J.J.V.; Correa-Rotter, R.; Rossing, P.; Sjöstrom, C.D.; Stefansson, B.V.; Toto, R.D.; et al. Effect of dapagliflozin on the rate of decline in kidney function in patients with chronic kidney disease with and without type 2 diabetes: A prespecified analysis from the DAPA-CKD trial. Lancet Diabetes Endocrinol. 2021, 9, 743–754. [Google Scholar] [CrossRef]
- Mima, A. Sodium-Glucose Cotransporter 2 Inhibitors in Patients with Non-Diabetic Chronic Kidney Disease. Adv. Ther. 2021, 38, 2201–2212. [Google Scholar] [CrossRef]
- Benetti, E.; Mastrocola, R.; Vitarelli, G.; Cutrin, J.C.; Nigro, D.; Chiazza, F.; Mayoux, E.; Collino, M.; Fantozzi, R. Empagliflozin protects against diet-induced NLRP-3 inflammasome activation and lipid accumulation. J. Pharmacol. Exp. Ther. 2016, 359, 45–53. [Google Scholar] [CrossRef]
- Gembrandt, F.; Bartaun, C.; Jarzebska, N.; Mayoux, E.; Todorov, V.T.; Hohenstein, B.; Hugo, C. The SGLT2 inhibitor empagliflozin ameliorates early features of diabetic nephropathy in BTBR ob/ob type 2 diabetic mice with and without hypertension. Am. J. Physiol. Renal. Physiol. 2014, 307, F317–F325. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.X.; Levi, J.; Luo, Y.; Kopp, J.B.; Rosenberg, A.Z.; Levi, M. SGLT2 Protein expression is increased in human diabetic nephropathy: SGTL2 protein inhibition decreases renal lipid accumulation, inflammation, and the development of nephropathy in diabetic mice. J. Biol. Chem. 2017, 292, 5335–5348. [Google Scholar] [CrossRef] [PubMed]
- Yurista, S.R.; Silljé, H.H.W.; Oberdorf-Maass, S.U.; Schouten, E.M.; Pavez Giani, M.G.; Hillebrands, J.L.; van Goor, H.; van Veldhuisen, D.J.; de Boer, R.A.; Westenbrink, B.D. Sodium-glucose co-transporter 2 inhibition with empagliflozin improves cardiac function in non-diabetic rats with left ventricular dysfunction after myocardial infarction. Eur. J. Heart Fail. 2019, 21, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Connelly, K.A.; Zhang, Y.; Desjardins, J.F.; Nghiem, L.; Visram, A.; Batchu, S.N.; Yerra, V.G.; Kabir, G.; Thai, K.; Advani, A.; et al. Load-independent effects of empagliflozin contribute to improved cardiac function in experimental heart failure with reduced ejection fraction. Cardiovasc. Diabetol. 2020, 19, 13. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.C.; Shiou, Y.L.; Jhuo, S.J.; Chang, C.Y.; Liu, P.L.; Jhuang, W.J.; Dai, Z.K.; Chen, W.Y.; Chen, Y.F.; Lee, A.S. The sodium-glucose co-transporter 2 inhibitor empagliflozin attenuates cardiac fibrosis and improves ventricular hemodynamics in hypertensive heart failure rats. Cardiovasc. Diabetol. 2019, 18, 45. [Google Scholar] [CrossRef]
- Yang, C.C.; Chen, Y.T.; Wallace, C.G.; Chen, K.H.; Cheng, B.C.; Sung, P.H.; Li, Y.-C.; Ko, S.-F.; Chang, H.-W.; Yip, H.-K. Early administration of empagliflozin preserved heart function in cardiorenal syndrome in rat. Biomed. Pharmacother. 2019, 109, 658–670. [Google Scholar] [CrossRef]
- Hojná, S.; Rauchová, H.; Malínská, H.; Marková, I.; Hüttl, M.; Papoušek, F.; Behuliak, M.; Miklánková, D.; Vaňourková, Z.; Neckář, J.; et al. Antihypertensive and metabolic effects of empagliflozin in Ren-2 transgenic rats, an experimental non-diabetic model of hypertension. Biomed. Pharmacother. 2021, 144, 112246. [Google Scholar] [CrossRef]
- Hüttl, M.; Markova, I.; Miklankova, D.; Zapletalova, I.; Poruba, M.; Haluzik, M.; Vaněčkova, I.; Malinska, H. In a Prediabetic Model, Empagliflozin Improves Hepatic Lipid Metabolism Independently of Obesity and before Onset of Hyperglycemia. Int. J. Mol. Sci. 2021, 22, 11513. [Google Scholar] [CrossRef]
- Pravenec, M.; Kajiya, T.; Zídek, V.; Landa, V.; Mlejnek, P.; Simáková, M.; Silhavý, J.; Malínská, H.; Oliyarnyk, O.; Kazdová, L.; et al. Effects of human C-reactive protein on pathogenesis of features of the metabolic syndrome. Hypertension 2011, 57, 731–737. [Google Scholar] [CrossRef]
- Malinska, H.; Hüttl, M.; Oliyarnyk, O.; Bratova, M.; Kazdova, L. Conjugated linoleic acid reduces visceral and ectopic lipid accumulation and insulin resistance in chronic severe hypertriacylglycerolemia. Nutrition 2015, 31, 1045–1051. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Hrdlička, J.; Neckář, J.; Papoušek, F.; Husková, Z.; Kikerlová, S.; Vaňourková, Z.; Vernerová, Z.; Akat, F.; Vašinová, J.; Hammock, B.D.; et al. Epoxyeicosatrienoic Acid-Based Therapy Attenuates the Progression of Postischemic Heart Failure in Normotensive Sprague-Dawley but Not in Hypertensive Ren-2 Transgenic Rats. Front. Pharmacol. 2019, 10, 159. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]
- Nakano, Y.; Hirano, T.; Uehara, K.; Nishibayashi, S.; Hattori, K.; Aihara, M.; Yamada, Y. New rat model induced by anti-glomerular basement membrane antibody shows severe glomerular adhesion in early stage and quickly progresses to end-stage renal failure. Pathol. Int. 2008, 58, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Sumithran, E.; Glasgow, E.F.; Atkins, R.C. The enhancement of aminonucleoside nephrosis by the co-administration of protamine. Kidney Int. 1987, 32, 691–699. [Google Scholar] [CrossRef]
- Zicha, J.; Kunes, J. Ontogenetic Aspects of Hypertension Development: Analysis in the Rat. Physiol. Rev. 1999, 79, 1227–1282. [Google Scholar] [CrossRef]
- Jiang, T.; Wang, X.X.; Scherzer, P.; Wilson, P.; Tallman, J.; Takahashi, H.; Li, J.; Iwahashi, M.; Sutherland, E.; Arend, L.; et al. Farnesoid X receptor modulates renal lipid metabolism, fibrosis, and diabetic nephropathy. Diabetes 2007, 56, 2485–2493. [Google Scholar] [CrossRef]
- Sun, L.; Halaihel, N.; Zhang, W.; Rogers, T.; Levi, M. Role of sterol regulatory element-binding protein 1 in regulation of renal lipid metabolism and glomerulosclerosis in diabetes mellitus. J. Biol. Chem. 2002, 277, 18919–18927. [Google Scholar] [CrossRef]
- Proctor, G.; Jiang, T.; Iwahashi, M.; Wang, Z.; Li, J.; Levi, M. Regulation of renal fatty acid and cholesterol metabolism, inflammation, and fibrosis in Akita and OVE26 mice with type 1 diabetes. Diabetes 2006, 55, 2502–2509. [Google Scholar] [CrossRef]
- Wang, Z.; Jiang, T.; Li, J.; Proctor, G.; McManaman, J.L.; Lucia, S.; Chua, S.; Levi, M. Regulation of renal lipid metabolism, lipid accumulation, and glomerulosclerosis in FVBdb/db mice with type 2 diabetes. Diabetes 2005, 54, 2328–2335. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.X.; Edelstein, M.H.; Gafter, U.; Qiu, L.; Luo, Y.; Dobrinskikh, E.; Lucia, S.; Adorini, L.; D’Agati, V.D.; Levi, J.; et al. G protein-coupled bile acid receptor TGR5 activation inhibits kidney disease in obesity and diabetes. J. Am. Soc. Nephrol. 2016, 27, 1362–1378. [Google Scholar] [CrossRef] [PubMed]
- Herman-Edelstein, M.; Scherzer, P.; Tobar, A.; Levi, M.; Gafter, U. Altered renal lipid metabolism and renal lipid accumulation in human diabetic nephropathy. J. Lipid. Res. 2014, 55, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, K.; Takata, T.; Sugihara, T.; Matono, T.; Koda, M.; Kanda, T.; Taniguchi, S.; Ida, A.; Mae, Y.; Yamamoto, M.; et al. Ipragliflozin Ameliorates Endoplasmic Reticulum Stress and Apoptosis through Preventing Ectopic Lipid Deposition in Renal Tubules. Int. J. Mol. Sci. 2019, 21, 190. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Sugiura, Y.; Saito, H.; Sugahara, M.; Higashijima, Y.; Yamaguchi, J.; Inagi, R.; Suematsu, M.; Nangaku, M.; Tanaka, T. Sodium-glucose cotransporter 2 inhibition normalizes glucose metabolism and suppresses oxidative stress in the kidneys of diabetic mice. Kidney Int. 2018, 94, 912–925. [Google Scholar] [CrossRef]
- Charlton, A.; Garzarella, J.; Jandeleit-Dahm, K.A.M.; Jha, J.C. Oxidative stress and inflammation in renal and cardiovascular complications of diabetes. Biology 2020, 10, 18. [Google Scholar] [CrossRef]
- Pirklbauer, M. Anti-inflammatory potencial of empagliflozin. Inflammopharmacology 2021, 29, 573–576. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Matsui, T.; Yamagishi, S. Tofogliflozin, a highly selective inhibitor of SGLT2 blocks proinflammatory and proapoptotic effects of glucose overload on proximal tubular cells partly by suppressing oxidative stress generation. Horm. Metab. Res. 2016, 48, 191–195. [Google Scholar] [CrossRef]
- Nishimura, R.; Tanaka, Y.; Koiwai, K.; Ishida, K.; Salsali, A. Effect of empagliflozin on free fatty acids and ketone bodies in Japanese patients with type 2 diabetes mellitus: A randomized controlled trial. Adv. Ther. 2019, 36, 2769–2782. [Google Scholar] [CrossRef]
- Abdurrachim, D.; Teo, X.Q.; Woo, C.C. Empagliflozin reduces myocardial ketone utilization while preserving glucose utilization in diabetic hypertensive heart disease: A hyperpolarized 13-C magnetic resonance spectroscopy study. Diabetes Obes. Metab. 2019, 21, 357–365. [Google Scholar] [CrossRef]
- Packer, M. Do sodium-glucose co-transporter-2 inhibitors prevent heart failure with a preserved ejection fraction by counterbalancing the effects of leptin? A novel hypothesis. Diabetes Obes. Metab. 2018, 20, 1361–1366. [Google Scholar] [CrossRef]
- Lieb, W.; Sullivan, L.M.; Harris, T.B. Plasma leptin levels and incidence of heart failure, cardiovascular disease, and total mortality in elderly individuals. Diabetes Care 2009, 32, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Pedone, C.; Roshanravan, B.; Scarlata, S.; Patel, K.V.; Ferrucci, L.; Incalzi, R.A. Longitudial association between serum leptin concetration and glomerular filtration rate in humans. PLoS ONE 2015, 10, e0117828. [Google Scholar] [CrossRef] [PubMed]
- Tentolouris, A.; Vlachakis, P.; Tzeravini, E.; Eleftheriadou, I.; Tentolouris, N. SGLT2 inhibitor: A review of their antidiabetic and cardioprotective effects. Int. J. Environ. Res. Public Health 2019, 16, 2965. [Google Scholar] [CrossRef]
- Vickers, S.P.; Cheetham, S.C.; Headland, K.R. Combination of the sodium-glucose cotransporter-2 inhibitor empagliflozin with orlistat or sibutramine further improves the body-weight reduction and glucose homeostasis of obese rats fed a cafeteria diet. Diabetes Metab. Syndr. Obes. 2014, 7, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Nasiri-Ansari, N.; Nikolopoulou, C.; Papoutsi, K.; Kyrou, I.; Mantzoros, C.S.; Kyriakopoulos, G.; Chatzigeorgiou, A.; Kalotychou, V.; Randeva, M.S.; Chatha, K.; et al. Empagliflozin Attenuates Non-Alcoholic Fatty Liver Disease (NAFLD) in High Fat Diet Fed ApoE(−/−) Mice by Activating Autophagy and Reducing ER Stress and Apoptosis. Int. J. Mol. Sci. 2021, 22, 818. [Google Scholar] [CrossRef] [PubMed]
- Lebeaupin, C.; Vallée, D.; Hazari, Y.; Hetz, C.; Chevet, E.; Bailly-Maitre, B. Endoplasmic reticulum stress signalling and the pathogenesis of non-alcoholic fatty liver disease. J. Hepatol. 2018, 69, 927–947. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Young | Adult | ||||||
|---|---|---|---|---|---|---|---|
| Control | Empa | Control | Empa | PAGE | PTREATMENT | PINTER-ACTION | |
| Body weight (g) | 347 ± 6 | 316 ± 7 ** | 379 ± 6 # | 368 ± 5 # | <0.001 | <0.01 | n.s. |
| Epididymal fat (g/100 g BW) | 0.58 ± 0.02 | 0.56 ± 0.01 | 0.52 ± 0.03 | 0.39 ± 0.02 *** | <0.001 | <0.01 | n.s. |
| Perirenal fat (g/100 g BW) | 0.43 ± 0.04 | 0.33 ± 0.01 | 0.57 ± 0.06 | 0.41 ± 0.03 * | <0.01 | <0.01 | n.s. |
| Liver weight (g/100 g BW) | 3.05 ± 0.11 | 3.30 ± 0.04 * | 4.03 ± 0.09 # | 3.68 ± 0.04 **# | <0.001 | n.s. | <0.001 |
| Heart weight (g/100 g BW) | 0.36 ± 0.01 | 0.34 ± 0.01 | 0.41 ± 0.01 # | 0.39 ± 0.01 *# | <0.001 | <0.05 | n.s. |
| Kidney weight (g/100 g BW) | 0.66 ± 0.03 | 0.70 ± 0.01 | 0.74 ± 0.01 | 0.75 ± 0.01 | <0.01 | n.s. | n.s. |
| Mean arterial pressure (mm Hg) | 192 ± 11 | 197 ± 7 | 190 ± 4 | 197 ± 6 | n.s. | n.s. | n.s. |
| Heart rate (bpm) | 373 ± 8 | 380 ± 5 | 377 ± 8 | 355 ± 8 # | n.s. | n.s. | n.s. |
| SHR-CRP | ||||
|---|---|---|---|---|
| Baseline | End of Study | |||
| Control | Empagliflozin | Control | Empagliflozin | |
| AWTd (mm) | 1.96 ± 0.02 | 1.93 ± 0.03 | 2.14 ± 0.03 @ | 1.96 ± 0.04 * |
| LVDd (mm) | 7.75 ± 0.14 | 7.20 ± 0.09 | 7.72 ± 0.11 | 7.41 ± 0.27 |
| PWTd (mm) | 1.95 ± 0.02 | 1.95 ± 0.03 | 2.08 ± 0.02 @ | 1.92 ± 0.05 * |
| AWTs (mm) | 2.87 ± 0.06 | 2.86 ± 0.06 | 2.99 ± 0.06 @ | 2.80 ± 0.04 * |
| LVDs (mm) | 4.75 ± 0.09 | 4.79 ± 0.07 | 4.35 ± 0.05 | 4.58 ± 0.20 |
| PWTs (mm) | 2.87 ± 0.04 | 2.88 ± 0.06 | 3.00 ± 0.04 @ | 2.80 ± 0.04 * |
| FS (%) | 39.1 ± 0.47 | 39.5 ± 0.26 | 37.8 ± 0.31 | 38.3 ± 0.53 |
| HR (bpm) | 329 ± 9 | 334 ± 5 | 316 ± 9 | 306 ± 11 |
| Young | Adult | ||||||
|---|---|---|---|---|---|---|---|
| Control | Empagliflozin | Control | Empagliflozin | PAGE | PTREATMENT | PINTER -ACTION | |
| Triglycerides (mmol/L) | 0.35 ± 0.04 | 0.39 ± 0.04 | 0.72 ± 0.13 # | 0.57 ± 0.04 # | <0.01 | n.s. | n.s. |
| Total cholesterol (mmol/L) | 1.24 ± 0.19 | 1.00 ± 0.03 | 1.33 ± 0.08 | 1.50 ± 0.05 # | <0.01 | n.s. | n.s. |
| HDL-cholesterol (mmol/L) | 1.06 ± 0.13 | 0.85 ± 0.01 * | 1.06 ± 0.05 | 1.22 ± 0.02 # | <0.05 | n.s. | <0.05 |
| NEFA (mmol/L) | 0.51 ± 0.02 | 0.44 ± 0.01 * | 0.44 ± 0.02 | 0.47 ± 0.02 | n.s. | n.s. | <0.05 |
| Fasting glucose (mmol/L) | 4.3 ± 0.1 | 4.3 ± 0 1 | 4.6 ± 0.1 | 4.5 ± 0.2 | n.s. | n.s. | n.s. |
| Non-fasting glucose (mmol/L) | 7.1 ± 0.2 | 6.8 ± 0.1 | 7.4 ± 0.3 | 6.9 ± 0.3 | n.s. | n.s. | n.s. |
| AUC0–120 (OGT test) | 842 ± 19 | 802 ± 11 | 842 ± 25 | 905 ± 63 | n.s. | n.s. | n.s. |
| Lipogenesis 0 (nmol/g/2 h) | 1397 ± 161 | 1582 ± 204 | 1105 ± 82 | 1206 ± 92 | <0.05 | n.s. | n.s. |
| Lipogenesis 250 (nmol/g/2 h) | 2007 ± 106 | 2372 ± 350 | 1948 ± 313 | 1705 ± 232 | <0.05 | n.s. | n.s. |
| Lipolysis 0 (nmol/g/2 h) | 4.70 ± 0.24 | 3.84 ± 0.32 | 4.61 ± 0.54 | 4.78 ± 0.49 | n.s. | n.s. | n.s. |
| Lipolysis 250 (nmol/g/2 h) | 6.52 ± 0.34 | 5.28 ± 0.76 | 8.06 ± 0.76 | 8.36 ± 0.98 # | n.s. | n.s. | n.s. |
| Glycogenesis 0 (nmol/g/2 h) | 1366 ± 240 | 1505 ± 307 | 1803 ± 128 | 1843 ± 243 | <0.01 | n.s. | n.s. |
| Glycogenesis 250 (nmol/g/2 h) | 1642 ± 248 | 2522 ± 462 | 1995 ± 214 | 2059 ± 211 | n.s. | n.s. | n.s. |
| Young | Adult | ||||||
|---|---|---|---|---|---|---|---|
| Control | Empagliflozin | Control | Empagliflozin | PAGE | PTREATMENT | PINTER- ACTION | |
| hs CRP human (mg/mL) | 299 ± 59 | 221 ± 5 * | 201 ± 9 | 227 ± 10 | n.s. | n.s. | <0.05 |
| hs CRP rat (mg/mL) | 0.82 ± 0.15 | 0.51 ± 0.15 | 1.95 ± 0.10 # | 1.42 ± 0.13 *# | <0.001 | <0.01 | n.s. |
| MCP-1 (ng/mL) | 4.36 ± 0.67 | 3.53 ± 0.17 | 9.47 ± 0.52 # | 7.43 ± 0.17 **# | <0.001 | <0.01 | n.s. |
| IL-6 (pg/mL) | 51.86 ± 23.17 | 50.62 ± 17.96 | 49.75 ± 15.86 | 50.26 ± 17.23 | n.s. | n.s. | n.s. |
| Leptin (ng/mL) | 2.39 ± 0.39 | 0.73 ± 0.23 *** | 1.35 ± 0.03 | 1.23 ± 0.02 | n.s. | <0.001 | <0.01 |
| Young | Adult | ||||||
|---|---|---|---|---|---|---|---|
| Control | Empagliflozin | Control | Empagliflozin | PAGE | PTREATMENT | PINTER- ACTION | |
| SOD | 0.045 ± 0.003 | 0.048 ± 0.003 | 0.046 ± 0.003 | 0.050 ± 0.002 | n.s. | n.s. | n.s. |
| GSH-Px | 148 ± 14 | 199 ± 12 ** | 163 ± 14 | 214 ± 8 ** | n.s. | <0.001 | n.s. |
| GR | 73 ± 5 | 67 ± 7 | 78 ± 4 | 86 ± 3 # | <0.05 | n.s. | n.s. |
| CAT | 626 ± 33 | 790 ± 32 *** | 553 ± 21 | 636 ± 30 *# | <0.001 | <0.001 | n.s. |
| CD | 21.7 ± 1.2 | 17.0 ± 1.8 * | 22.6 ± 1.0 | 16.6 ± 1.2 ** | n.s. | <0.001 | n.s. |
| TBARS | 0.78 ± 0.03 | 0.50 ± 0.03 *** | 0.65 ± 0.02 | 0.52 ± 0.03 ** | n.s. | <0.001 | <0.05 |
| GSH | 25.8 ± 5.3 | 37.8 ± 4.3 *** | 13.7 ± 0.7 # | 14.3 ± 0.1 # | <0.001 | <0.001 | <0.001 |
| GSSG | 2.4 ± 0.1 | 2.7 ± 0.1 | 1.8 ± 0.1 # | 1.8 ± 0.1 # | <0.01 | n.s. | n.s. |
| GSH/GSSG | 12.1 ± 0.8 | 14.3 ± 0.6 | 7.9 ± 0.4 # | 7.9 ± 0.2 # | <0.001 | n.s. | n.s. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malínská, H.; Hüttl, M.; Marková, I.; Miklánková, D.; Hojná, S.; Papoušek, F.; Šilhavý, J.; Mlejnek, P.; Zicha, J.; Hrdlička, J.; et al. Beneficial Effects of Empagliflozin Are Mediated by Reduced Renal Inflammation and Oxidative Stress in Spontaneously Hypertensive Rats Expressing Human C-Reactive Protein. Biomedicines 2022, 10, 2066. https://doi.org/10.3390/biomedicines10092066
Malínská H, Hüttl M, Marková I, Miklánková D, Hojná S, Papoušek F, Šilhavý J, Mlejnek P, Zicha J, Hrdlička J, et al. Beneficial Effects of Empagliflozin Are Mediated by Reduced Renal Inflammation and Oxidative Stress in Spontaneously Hypertensive Rats Expressing Human C-Reactive Protein. Biomedicines. 2022; 10(9):2066. https://doi.org/10.3390/biomedicines10092066
Chicago/Turabian StyleMalínská, Hana, Martina Hüttl, Irena Marková, Denisa Miklánková, Silvie Hojná, František Papoušek, Jan Šilhavý, Petr Mlejnek, Josef Zicha, Jaroslav Hrdlička, and et al. 2022. "Beneficial Effects of Empagliflozin Are Mediated by Reduced Renal Inflammation and Oxidative Stress in Spontaneously Hypertensive Rats Expressing Human C-Reactive Protein" Biomedicines 10, no. 9: 2066. https://doi.org/10.3390/biomedicines10092066
APA StyleMalínská, H., Hüttl, M., Marková, I., Miklánková, D., Hojná, S., Papoušek, F., Šilhavý, J., Mlejnek, P., Zicha, J., Hrdlička, J., Pravenec, M., & Vaněčková, I. (2022). Beneficial Effects of Empagliflozin Are Mediated by Reduced Renal Inflammation and Oxidative Stress in Spontaneously Hypertensive Rats Expressing Human C-Reactive Protein. Biomedicines, 10(9), 2066. https://doi.org/10.3390/biomedicines10092066