
Veno-Arterial Extracorporeal Membrane Oxygenation (ECMO) Impairs Bradykinin-Induced Relaxation in Neonatal Porcine Coronary Arteries

 , , , , , , , , ,
, , , , , , , , ,  ,
,  add
Show full author list
add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Blood Collection for Extracorporeal Priming Circuit
2.2. Surgical Procedures
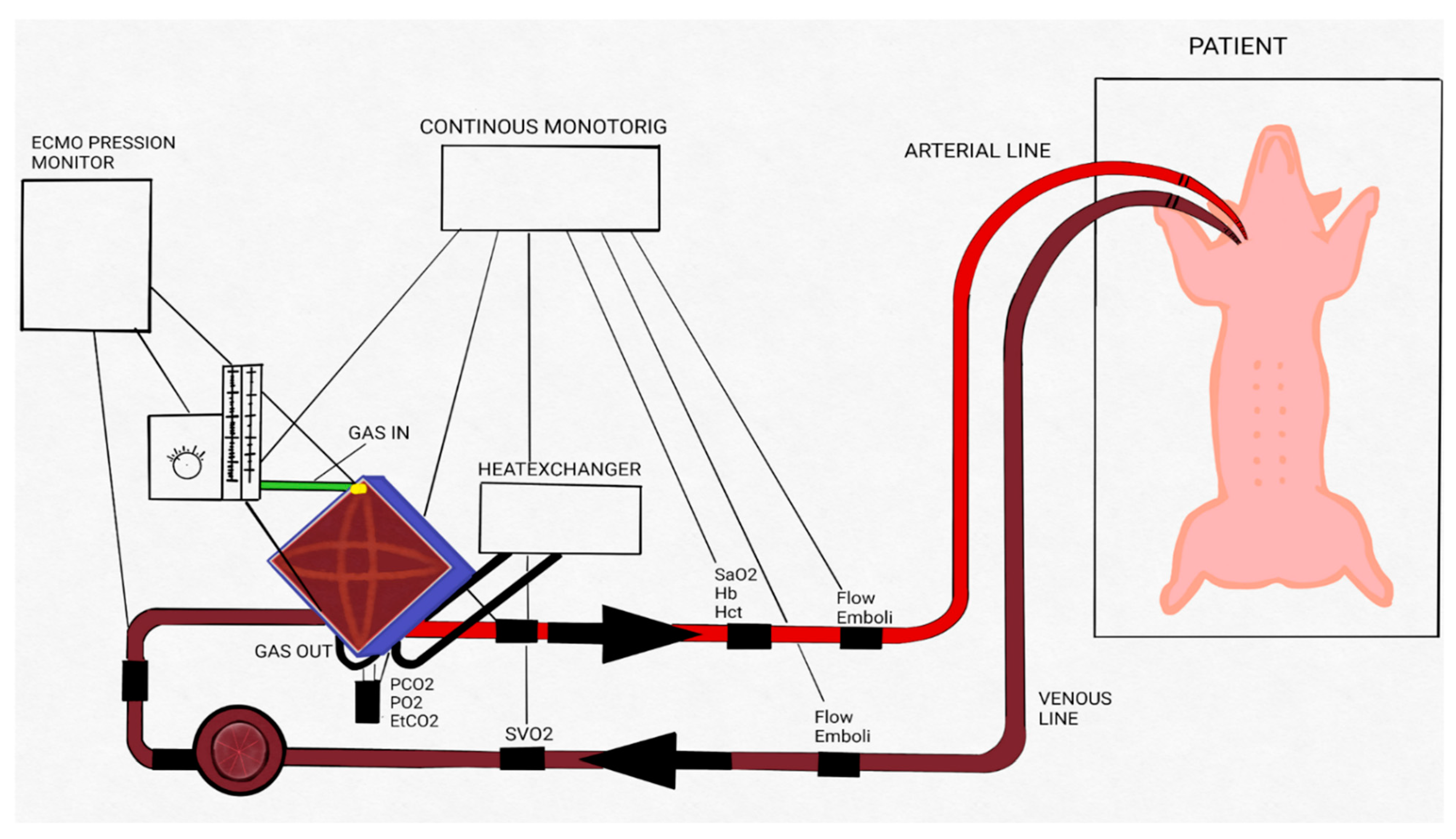
2.3. ECMO Procedure
2.4. Euthanasia and Tissue Collection
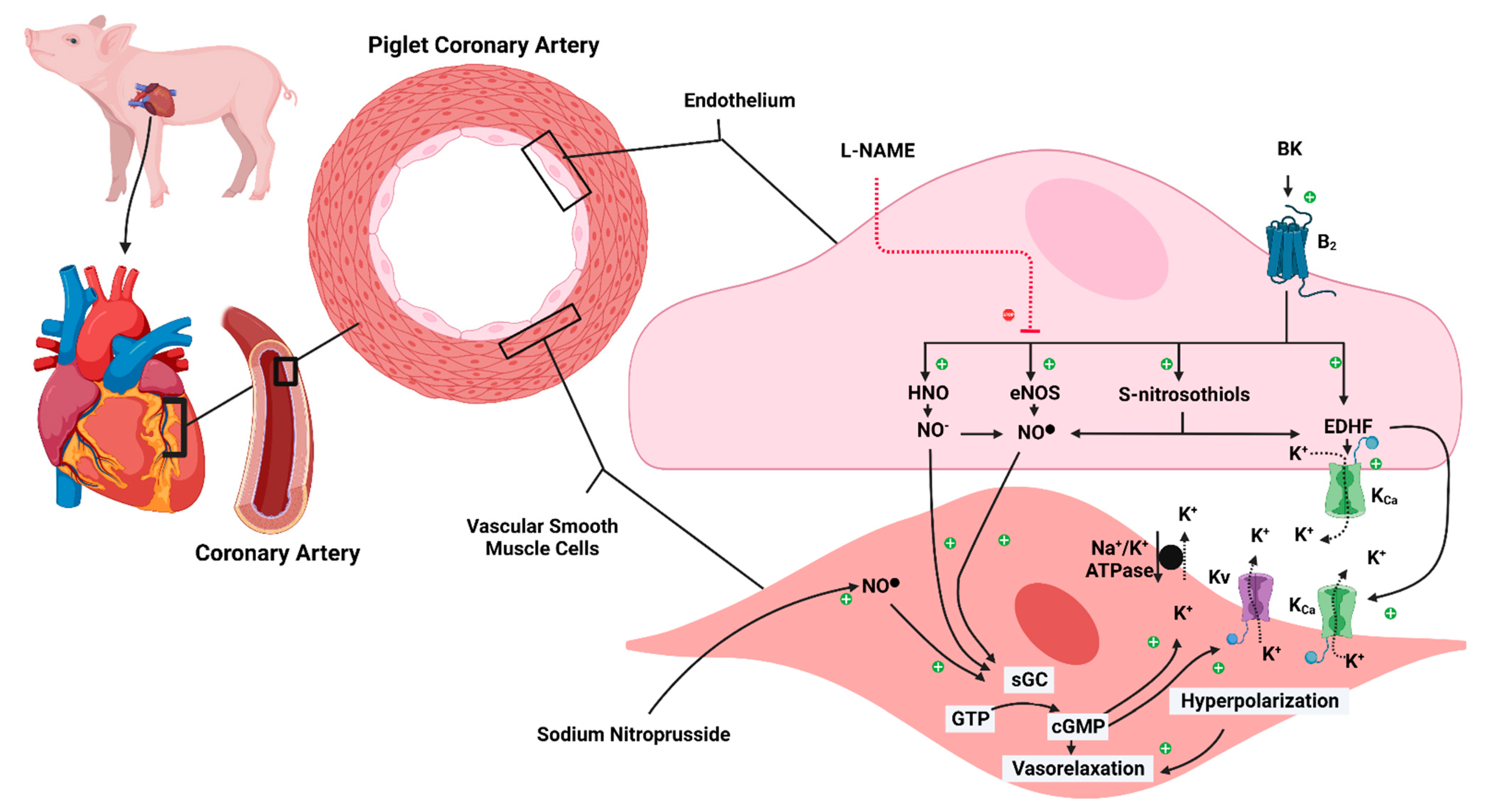
2.5. Vascular Reactivity Studies
2.6. Drugs and Solutions
2.7. Data Analysis
3. Results
3.1. ECMO Procedure
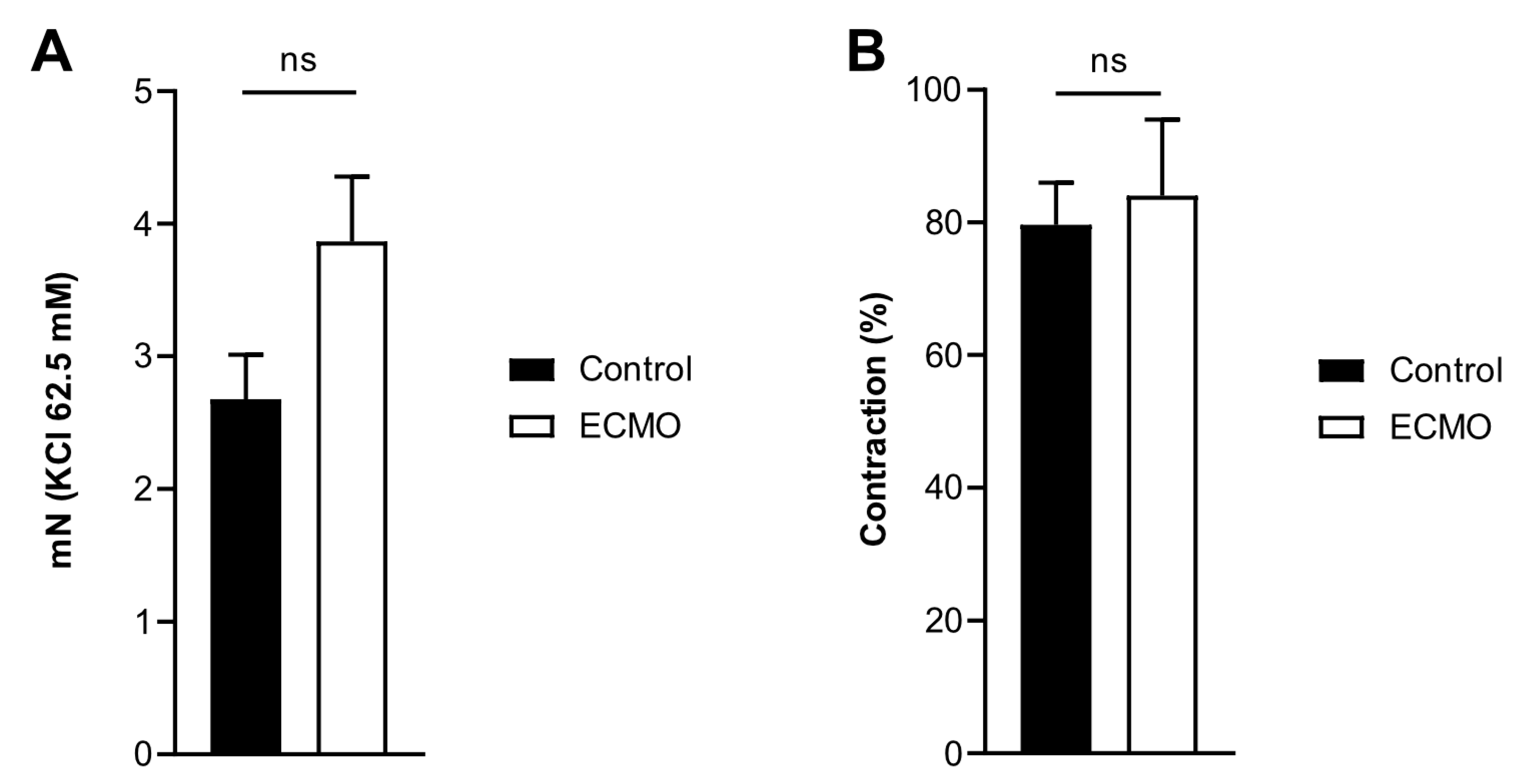
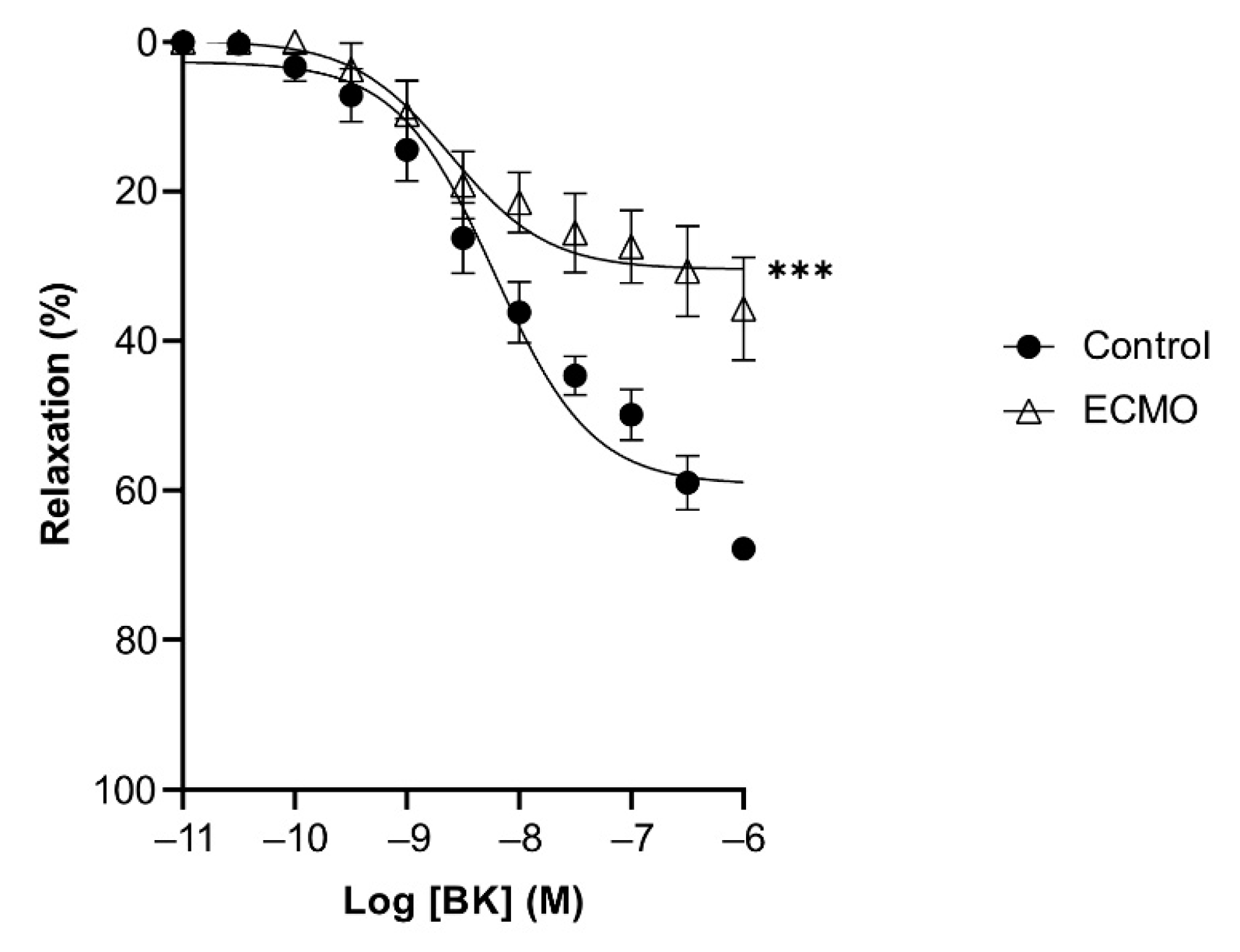
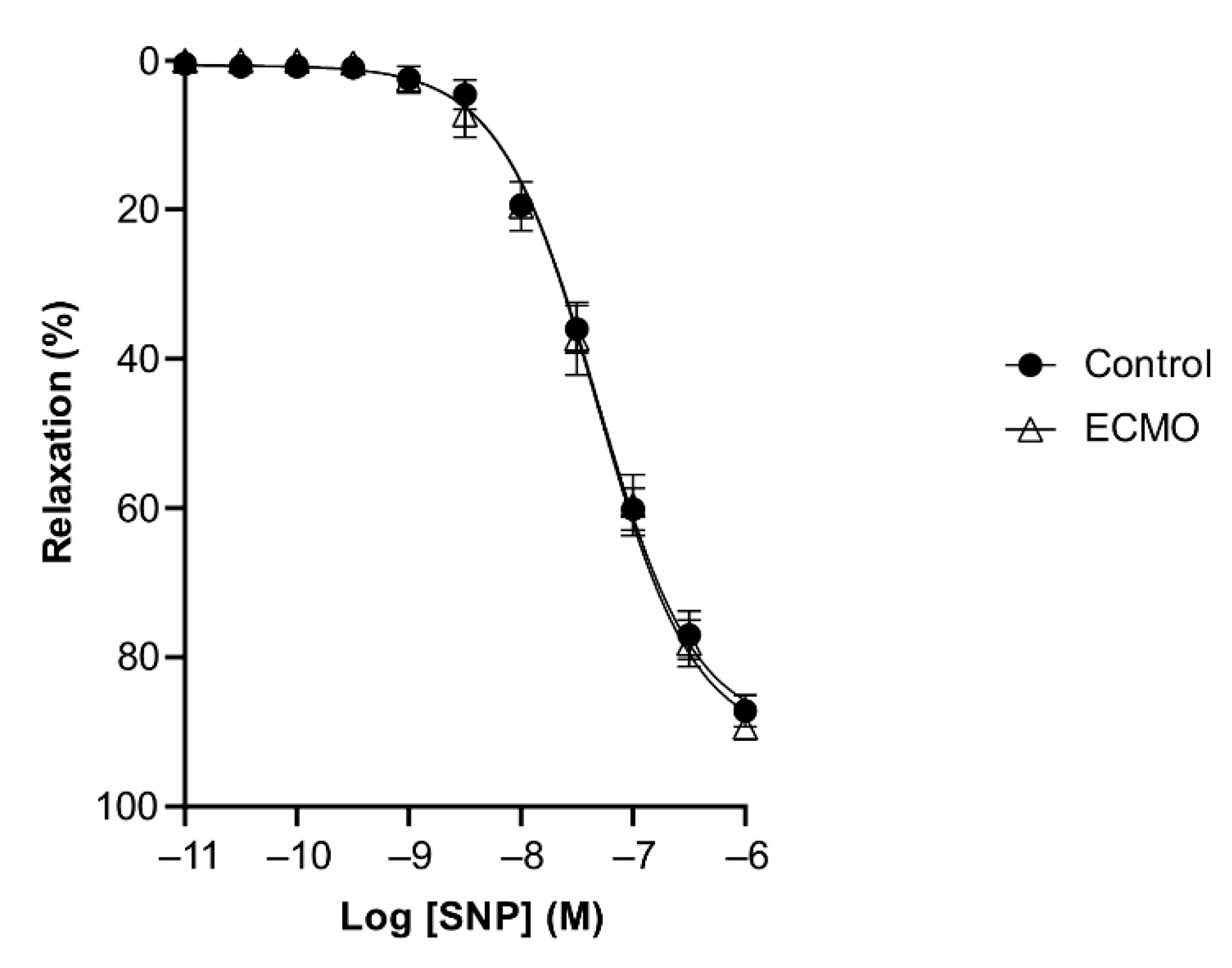
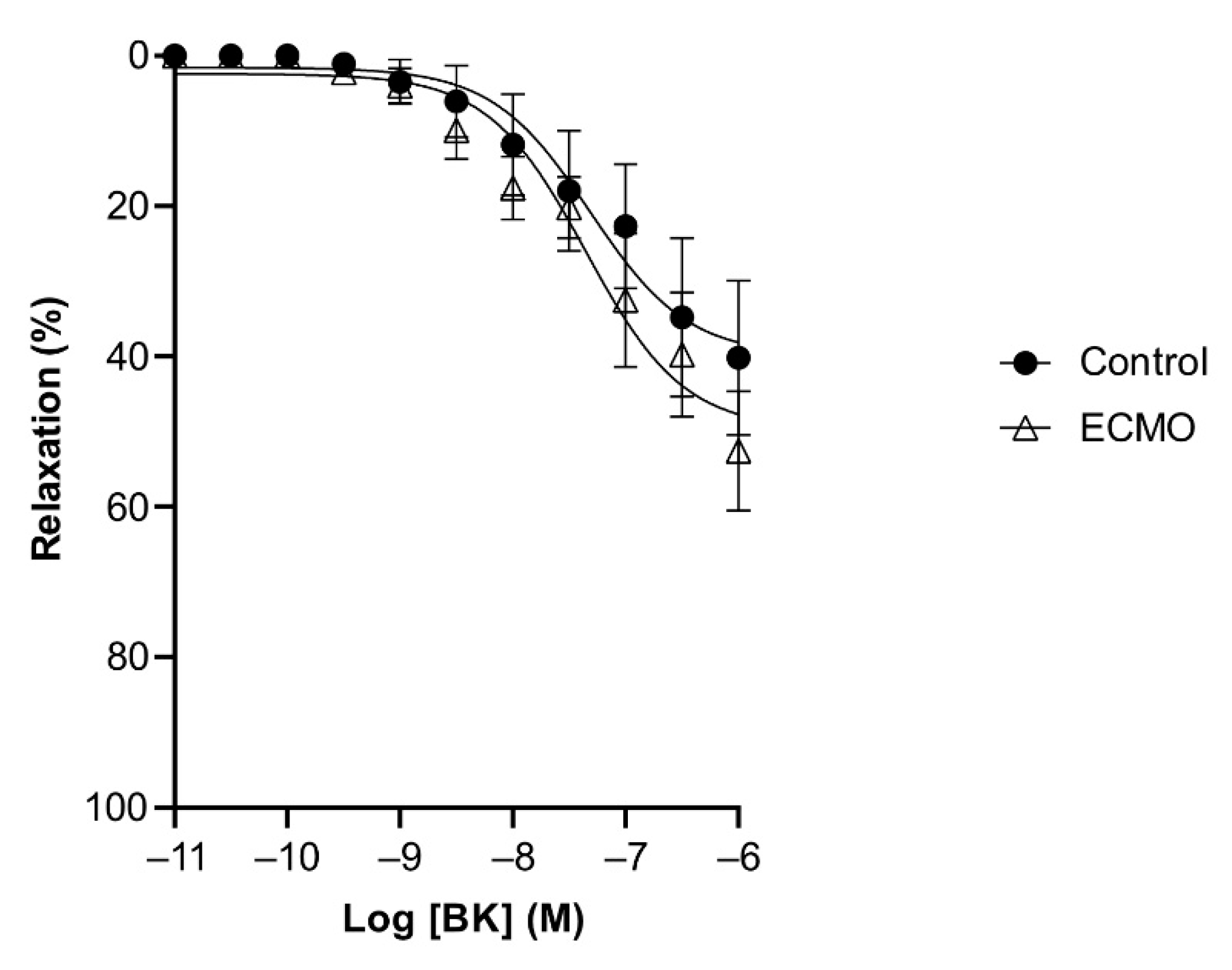
3.2. Reactivity of Coronary Arteries
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xiong, J.; Zhang, L.; Bao, L. Complications and mortality of venovenous extracorporeal membrane oxygenation in the treatment of neonatal respiratory failure: A systematic review and meta-analysis. BMC Pulm. Med. 2020, 20, 124. [Google Scholar] [CrossRef] [PubMed]
- Butt, W.W.; Chiletti, R. ECMO for neonatal sepsis in 2019. Front. Pediatr. 2020, 8, 50. [Google Scholar] [CrossRef] [PubMed]
- Roeleveld, P.P.; Mendonca, M. Neonatal cardiac ECMO in 2019 and beyond. Front. Pediatr. 2019, 7, 327. [Google Scholar] [CrossRef] [PubMed]
- Rousse, N.; Juthier, F.; Pinçon, C.; Hysi, I.; Banfi, C.; Robin, E.; Fayad, G.; Jegou, B.; Prat, A.; Vincentelli, A. ECMO as a bridge to decision: Recovery, VAD, or heart transplantation? Int. J. Cardiol. 2015, 187, 620–627. [Google Scholar] [CrossRef]
- Short, B. Neonatal ECMO: Are indications changing? Int. J. Artif. Organs 1995, 18, 562–564. [Google Scholar] [CrossRef] [PubMed]
- Van Ommen, C.H.; Neunert, C.E.; Chitlur, M.B. Neonatal ECMO. Front. Med. 2018, 5, 289. [Google Scholar] [CrossRef]
- ELSO. ELSO Registry Report. International Summary on 1 April 2022. Available online: https://www.elso.org (accessed on 11 June 2022).
- Amodeo, I.; di Nardo, M.; Raffaeli, G.; Kamel, S.; Macchini, F.; Amodeo, A.; Mosca, F.; Cavallaro, G. Neonatal respiratory and cardiac ECMO in Europe. Eur. J. Pediatr. 2021, 180, 1675–1692. [Google Scholar] [CrossRef]
- Fletcher, K.; Chapman, R.; Keene, S. An overview of medical ECMO for neonates. Semin. Perinatol. 2018, 42, 68–79. [Google Scholar] [CrossRef]
- Millar, J.E.; Fanning, J.P.; McDonald, C.I.; McAuley, D.F.; Fraser, J.F. The inflammatory response to extracorporeal membrane oxygenation (ECMO): A review of the pathophysiology. Crit. Care 2016, 20, 387. [Google Scholar] [CrossRef]
- Raffaeli, G.; Ghirardello, S.; Passera, S.; Mosca, F.; Cavallaro, G. Oxidative stress and neonatal respiratory extracorporeal membrane oxygenation. Front. Physiol. 2018, 9, 1739. [Google Scholar] [CrossRef]
- Wendel, H.P.; Scheule, A.M.; Eckstein, F.S.; Ziemer, G. Haemocompatibility of paediatric membrane oxygenators with heparin-coated surfaces. Perfusion 1999, 14, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Graulich, J.; Sonntag, J.; Marcinkowski, M.; Bauer, K.; Kossel, H.; Buhrer, C.; Obladen, M.; Versmold, H.T. Complement activation by in vivo neonatal and in vitro extracorporeal membrane oxygenation. Mediat. Inflamm. 2002, 11, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Fortenberry, J.D.; Bhardwaj, V.; Niemer, P.; Cornish, J.D.; Wright, J.A.; Bland, L. Neutrophil and cytokine activation with neonatal extracorporeal membrane oxygenation. J. Pediatr. 1996, 128, 670–678. [Google Scholar] [CrossRef]
- DePuydt, L.E.; Schuit, K.E.; Smith, S.D. Effect of extracorporeal membrane oxygenation on neutrophil function in neonates. Crit. Care Med. 1993, 21, 1324–1327. [Google Scholar] [CrossRef] [PubMed]
- Tataranno, M.L.; Perrone, S.; Longini, M.; Buonocore, G. New antioxidant drugs for neonatal brain injury. Oxid. Med. Cell. Longev. 2015, 2015, 108251. [Google Scholar] [CrossRef]
- Ingyinn, M.; Lee, J.; Short, B.L.; Viswanathan, M. Venoarterial extracorporeal membrane oxygenation impairs basal nitric oxide production in cerebral arteries of newborn lambs. Pediatr. Crit. Care Med. 2000, 1, 161–165. [Google Scholar] [CrossRef]
- Ingyinn, M.; Rais-Bahrami, K.; Viswanathan, M.; Short, B.L. Altered cerebrovascular responses after exposure to venoarterial extracorporeal membrane oxygenation: Role of the nitric oxide pathway. Pediatr. Crit. Care Med. 2006, 7, 368–373. [Google Scholar] [CrossRef]
- Danser, A.J.; Tom, B.; de Vries, R.; Saxena, P.R. L-NAME-resistant bradykinin-induced relaxation in porcine coronary arteries is NO-dependent: Effect of ACE inhibition. Br. J. Pharmacol. 2000, 131, 195–202. [Google Scholar] [CrossRef]
- Batenburg, W.W.; de Vries, R.; Saxena, P.R.; Danser, A.J. LS-nitrosothiols: Endothelium-derived hyperpolarizing factors in porcine coronary arteries? J. Hypertens. 2004, 22, 1927–1936. [Google Scholar] [CrossRef]
- Batenburg, W.W.; Kappers, M.H.; Eikmann, M.J.; Ramzan, S.N.; de Vries, R.; Danser, A.J. Light-induced vs. bradykinin-induced relaxation of coronary arteries: Do S-nitrosothiols act as endothelium-derived hyperpolarizing factors? J. Hypertens. 2009, 27, 1631–1640. [Google Scholar] [CrossRef]
- Batenburg, W.W.; Popp, R.; Fleming, I.; de Vries, R.; Garrelds, I.M.; Saxena, P.R.; Danser, A.J. Bradykinin-induced relaxation of coronary microarteries: S-nitrosothiols as EDHF? Br. J. Pharmacol. 2004, 142, 125. [Google Scholar] [CrossRef] [PubMed]
- Botden, I.P.; Batenburg, W.W.; de Vries, R.; Langendonk, J.G.; Sijbrands, E.J.; Danser, A.J. Nitrite-and nitroxyl-induced relaxation in porcine coronary (micro-) arteries: Underlying mechanisms and role as endothelium-derived hyperpolarizing factor(s). Pharmacol. Res. 2012, 66, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Thollon, C.; Fournet-Bourguignon, M.P.; Saboureau, D.; Lesage, L.; Reure, H.; Vanhoutte, P.M.; Vilaine, J.P. Consequences of reduced production of NO on vascular reactivity of porcine coronary arteries after angioplasty: Importance of EDHF. Br. J. Pharmacol. 2002, 136, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Campbell, D.J.; Dixon, B.; Kladis, A.; Kemme, M.; Santamaria, J.D. Activation of the kallikrein-kinin system by cardiopulmonary bypass in humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 281, R1059–R1070. [Google Scholar] [CrossRef]
- Raffaeli, G.; Ghirardello, S.; Vanzati, M.; Baracetti, C.; Canesi, F.; Conigliaro, F.; Gentilino, V.; Macchini, F.; Fumagalli, M.; Ciralli, F.; et al. Start a neonatal extracorporeal membrane oxygenation program: A multistep team training. Front. Pediatr. 2018, 6, 151. [Google Scholar] [CrossRef]
- Macchini, F.; di Cesare, A.; Morandi, A.; Ichino, M.; Raffaeli, G.; Conigliaro, F.; Sorrentino, G.; Neri, S.; Mosca, F.; Leva, E.; et al. Surgical expertise in neonatal extracorporeal membrane oxygenation (ECMO): A single center experience. Front. Pediatr. 2019, 7, 398. [Google Scholar] [CrossRef]
- Mohammed, R.; Provitera, L.; Cavallaro, G.; Lattuada, D.; Ercoli, G.; Mosca, F.; Villamor, E. Vasomotor effects of hydrogen sulfide in human umbilical vessels. J. Physiol. Pharmacol. 2017, 68, 737–747. [Google Scholar]
- Provitera, L.; Cavallaro, G.; Griggio, A.; Raffaeli, G.; Amodeo, I.; Gulden, S.; Lattuada, D.; Ercoli, G.; Lonati, C.; Tomaselli, A.; et al. Cyclic nucleotide-dependent relaxation in human umbilical vessels. J. Physiol. Pharmacol. 2019, 70, 619–630. [Google Scholar] [CrossRef]
- Gonzalez-Luis, G.; Perez-Vizcaino, F.; Blanco, C.E.; Villamor, E. Age-related changes in isoprostane-mediated relaxation of piglet blood vessels. Front. Biosci.-Elite 2010, 2, 369–379. [Google Scholar]
- Fortin, J.P.; Marceau, F. Advances in the development of bradykinin receptor ligands. Curr. Top. Med. Chem. 2006, 6, 1353–1363. [Google Scholar] [CrossRef]
- Marceau, F.; Regoli, D. Bradykinin receptor ligands: Therapeutic perspectives. Nat. Rev. Drug Discov. 2004, 3, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Basei, F.L.; Cabrini, D.A.; Figueiredo, C.P.; Forner, S.; Hara, D.B.; Nascimento, A.F.; Ceravolo, G.S.; Carvalho, M.H.C.; Bader, M.; Medeiros, R. Endothelium dependent expression and underlying mechanisms of des-Arg9-bradykinin-induced B1R-mediated vasoconstriction in rat portal vein. Peptides 2012, 37, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Larsson, M.; Rayzman, V.; Nolte, M.W.; Nickel, K.F.; Bjorkqvist, J.; Jamsa, A.; Hardy, M.P.; Fries, M.; Schmidbauer, S.; Hedenqvist, P.; et al. A factor XIIa inhibitory antibody provides thromboprotection in extracorporeal circulation without increasing bleeding risk. Sci. Transl. Med. 2014, 6, 222ra217. [Google Scholar] [CrossRef] [PubMed]
- Landis, R.C.; Brown, J.R.; Fitzgerald, D.; Likosky, D.S.; Shore-Lesserson, L.; Baker, R.A.; Hammon, J.W. Attenuating the systemic inflammatory response to adult cardiopulmonary bypass: A critical review of the evidence base. J. Extra Corpor. Technol. 2014, 46, 197–211. [Google Scholar]
- Zhang, Y.; Adner, M.; Cardell, L.O. Up-regulation of bradykinin receptors in a murine in-vitro model of chronic airway inflammation. Eur. J. Pharmacol. 2004, 489, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Short, B.L.; Walker, L.K.; Bender, K.S.; Traystman, R.J. Impairment of cerebral autoregulation during extracorporeal membrane oxygenation in newborn lambs. Pediatr. Res. 1993, 33, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Wagerle, L.C.; Russo, P.; Dahdah, N.S.; Kapadia, N.; Davis, D.A. Endothelial dysfunction in cerebral microcirculation during hypothermic cardiopulmonary bypass in newborn lambs. J. Thorac. Cardiovasc. Surg. 1998, 115, 1047–1054. [Google Scholar] [CrossRef]
- Short, B.L. The effect of extracorporeal life support on the brain: A focus on ECMO. Semin. Perinatol. 2005, 29, 45–50. [Google Scholar] [CrossRef]
- Taylor, G.A.; Catena, L.M.; Garin, D.B.; Miller, M.K.; Short, B.L. Intracranial flow patterns in infants undergoing extracorporeal membrane oxygenation: Preliminary observations with Doppler US. Radiology 1987, 165, 671–674. [Google Scholar] [CrossRef]
- Taylor, G.A.; Martin, G.R.; Short, B.L. Cardiac determinants of cerebral blood flow during extracorporeal membrane oxygenation. Investig. Radiol. 1989, 24, 511–516. [Google Scholar] [CrossRef]
- Starke, R.M.; Chalouhi, N.; Ali, M.S.; Jabbour, P.M.; Tjoumakaris, S.I.; Gonzalez, L.F.; Rosenwasser, R.H.; Koch, W.J.; Dumont, A.S. The role of oxidative stress in cerebral aneurysm formation and rupture. Curr. Neurovasc. Res. 2013, 10, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Granger, D.N.; Senchenkova, E. Integrated systems physiology—From cell to function. In Inflammation and the Microcirculation; Morgan & Claypool Publishers: San Rafael, CA, USA, 2010. [Google Scholar]
- Grisham, M.B.; Granger, D.N.; Lefer, D.J. Modulation of leukocyte-endothelial interactions by reactive metabolites of oxygen and nitrogen: Relevance to ischemic heart disease. Free Radic. Biol. Med. 1998, 25, 404–433. [Google Scholar] [CrossRef]
- Grisham, M.B.; Jourd’Heuil, D.; Wink, D.A. Nitric oxide. I. Physiological chemistry of nitric oxide and its metabolites: Implications in inflammation. Am. J. Physiol. 1999, 276, G315–G321. [Google Scholar] [CrossRef]
- Stokes, K.Y.; Cooper, D.; Tailor, A.; Granger, D.N. Hypercholesterolemia promotes inflammation and microvascular dysfunction: Role of nitric oxide and superoxide. Free Radic. Biol. Med. 2002, 33, 1026–1036. [Google Scholar] [CrossRef]
- Milstien, S.; Katusic, Z. Oxidation of tetrahydrobiopterin by peroxynitrite: Implications for vascular endothelial function. Biochem. Biophys. Res. Commun. 1999, 263, 681–684. [Google Scholar] [CrossRef]
- Landmesser, U.; Dikalov, S.; Price, S.R.; McCann, L.; Fukai, T.; Holland, S.M.; Mitch, W.E.; Harrison, D.G. Oxidation of tetrahydrobiopterin leads to uncoupling of endothelial cell nitric oxide synthase in hypertension. J. Clin. Investig. 2003, 111, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Griendling, K.K.; FitzGerald, G.A. Oxidative stress and cardiovascular injury: Part I: Basic mechanisms and in vivo monitoring of ROS. Circulation 2003, 108, 1912–1916. [Google Scholar] [CrossRef] [PubMed]
- Mc, I.R.B.; Timpa, J.G.; Kurundkar, A.R.; Holt, D.W.; Kelly, D.R.; Hartman, Y.E.; Neel, M.L.; Karnatak, R.K.; Schelonka, R.L.; Anantharamaiah, G.M.; et al. Plasma concentrations of inflammatory cytokines rise rapidly during ECMO-related SIRS due to the release of preformed stores in the intestine. Lab. Investig. 2010, 90, 128–139. [Google Scholar] [CrossRef]
- Rafat, N.; Patry, C.; Sabet, U.; Viergutz, T.; Weiss, C.; Tonshoff, B.; Beck, G.; Schaible, T. Endothelial progenitor and mesenchymal stromal cells in newborns with congenital diaphragmatic hernia undergoing extracorporeal membrane oxygenation. Front. Pediatr. 2019, 7, 490. [Google Scholar] [CrossRef]
- Pietrasanta, C.; Pugni, L.; Ronchi, A.; Bottino, I.; Ghirardi, B.; Sanchez-Schmitz, G.; Borriello, F.; Mosca, F.; Levy, O. Vascular endothelium in neonatal sepsis: Basic mechanisms and translational opportunities. Front. Pediatr. 2019, 7, 340. [Google Scholar] [CrossRef]
- Zwiers, A.J.; Jsselstijn, H.I.; van Rosmalen, J.; Gischler, S.J.; de Wildt, S.N.; Tibboel, D.; Cransberg, K. CKD and hypertension during long-term follow-up in children and adolescents previously treated with extracorporeal membrane oxygenation. Clin. J. Am. Soc. Nephrol. 2014, 9, 2070–2078. [Google Scholar] [CrossRef] [PubMed]
- Ijsselstijn, H.; Schiller, R.M.; Holder, C.; Shappley, R.K.H.; Wray, J.; Hoskote, A. Extracorporeal life support organization (ELSO) guidelines for follow-up after neonatal and pediatric extracorporeal membrane oxygenation. ASAIO J. 2021, 67, 955–963. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Mean (S.E.M.) |
|---|---|
| HR (bpm) | 142.3 (3.09) |
| SpO2 (%) | 97.7 (0.43) |
| sAP (mmHg) | 96.8 (2.35) |
| dAP (mmHg) | 67.3 (2.39) |
| mAP (mmHg) | 76.3 (2.22) |
| ACT (s) | 211.3 (17.25) |
| Diuresis (mL/kg/h) | 3.9 (0.57) |
| T (°C) | 37 (0.17) |
| pH | 7.3 (0.00) |
| pO2 (mmHg) | 47.6 (1.95) |
| pCO2 (mmHg) | 45.6 (1.07) |
| BE | −1.8 (0.46) |
| HCO3− | 23.1 (0.43) |
| Lactate (mEq/L) | 4.6 (0.26) |
| Hb (g/dL) | 11.5 (0.26) |
| Hct (%) | 35.4 (0.80) |
| K+ (mmol/L) | 4 (0.08) |
| Na+ (mmol/L) | 145.2 (0.53) |
| Cl− (mmol/L) | 107.3 (0.58) |
| Ca2+ (mmol/L) | 1.4 (0.02) |
| Cardiac Index (mL/kg/m) | 86.6 (2.32) |
| ven P (mmHg) | −28.4 (0.93) |
| int P (mmHg) | 154.2 (9.97) |
| Art P (mmHg) | 130.9 (4.24) |
| Sweep Gas (L/m) | 0.25 (0.02) |
| FiO2 ECMO | 0.4 (0.01) |
| SvO2 (%) | 78.2 (0.12) |
| SaO2 (%) | 98.5 (0.21) |
| DO2 (mL/m) | 166.8 (4.36) |
| VO2 (mL/m) | 37.2 (0.92) |
| O2ER | 0.2 (0.00) |
| PEEP (cmH2O) | 4.9 (0.01) |
| PIP (cmH2O) | 20.5 (0.74) |
| RR (rpm) | 31.9 (1.99) |
| FiO2 ventilator | 0.2 (0.01) |
| Control (n = 5) | ECMO (n = 5) | |||||
|---|---|---|---|---|---|---|
| Emax | pD2 | n | Emax | pD2 | n | |
| Bradykinin | 59.21 (2.06) *** | 8.22 (0.10) | 5 | 30.41 (2.16) | 8.61 (0.21) | 5 |
| Bradykinin + L-NAME | 39.88 (5.50) | 7.31 (0.27) | 5 | 49.68 (4.41) | 7.35 (0.17) | 5 |
| SNP | 89.79 (1.77) | 7.32 (0.04) | 5 | 91.31 (2.34) | 7.32 (0.05) | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Provitera, L.; Amelio, G.S.; Tripodi, M.; Raffaeli, G.; Macchini, F.; Amodeo, I.; Gulden, S.; Cortesi, V.; Manzoni, F.; Cervellini, G.; et al. Veno-Arterial Extracorporeal Membrane Oxygenation (ECMO) Impairs Bradykinin-Induced Relaxation in Neonatal Porcine Coronary Arteries. Biomedicines 2022, 10, 2083. https://doi.org/10.3390/biomedicines10092083
Provitera L, Amelio GS, Tripodi M, Raffaeli G, Macchini F, Amodeo I, Gulden S, Cortesi V, Manzoni F, Cervellini G, et al. Veno-Arterial Extracorporeal Membrane Oxygenation (ECMO) Impairs Bradykinin-Induced Relaxation in Neonatal Porcine Coronary Arteries. Biomedicines. 2022; 10(9):2083. https://doi.org/10.3390/biomedicines10092083
Chicago/Turabian StyleProvitera, Livia, Giacomo S. Amelio, Matteo Tripodi, Genny Raffaeli, Francesco Macchini, Ilaria Amodeo, Silvia Gulden, Valeria Cortesi, Francesca Manzoni, Gaia Cervellini, and et al. 2022. "Veno-Arterial Extracorporeal Membrane Oxygenation (ECMO) Impairs Bradykinin-Induced Relaxation in Neonatal Porcine Coronary Arteries" Biomedicines 10, no. 9: 2083. https://doi.org/10.3390/biomedicines10092083
APA StyleProvitera, L., Amelio, G. S., Tripodi, M., Raffaeli, G., Macchini, F., Amodeo, I., Gulden, S., Cortesi, V., Manzoni, F., Cervellini, G., Tomaselli, A., Zuanetti, G., Lonati, C., Battistin, M., Kamel, S., Parente, V., Pravatà, V., Villa, S., Villamor, E., ... Cavallaro, G. (2022). Veno-Arterial Extracorporeal Membrane Oxygenation (ECMO) Impairs Bradykinin-Induced Relaxation in Neonatal Porcine Coronary Arteries. Biomedicines, 10(9), 2083. https://doi.org/10.3390/biomedicines10092083